
The Vaginal Microbiota, Bacterial Biofilms and Polymeric Drug-Releasing Vaginal Rings

 ,
,
Abstract
:1. Introduction
2. The Vaginal Microbiota: Healthy and Dysbiotic
2.1. Lactobacillus spp.
2.2. Gardnerella vaginalis
2.3. Atopobium vaginae and Prevotella spp.
2.4. Candida albicans
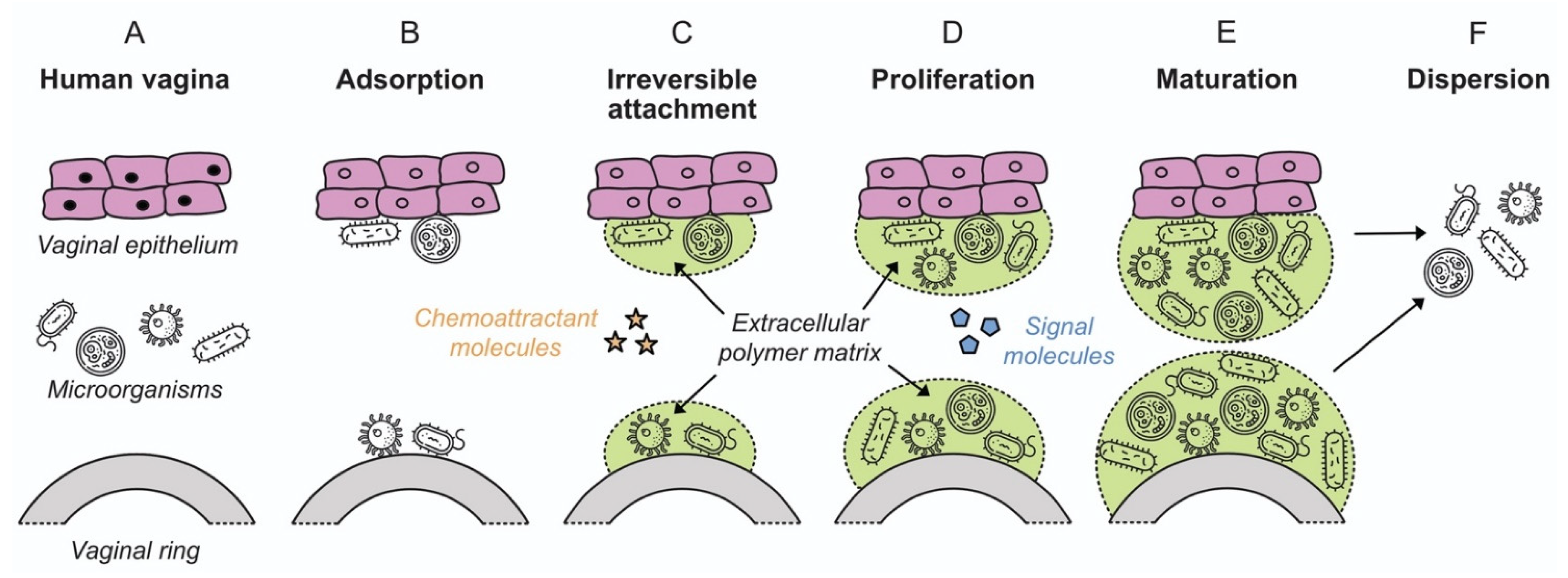
3. Biofilm Formation in the Human Vagina and Its Role in Vaginal Dysbiosis
4. Mechanistic Aspects of Biofilm Formation on Polymeric Surfaces
4.1. Bacterial Attachment to Polymeric Surfaces
4.2. Biofilm Proliferation and Maturation
4.3. Dispersion and Spread
4.4. Biofilm Formation on Medical Devices
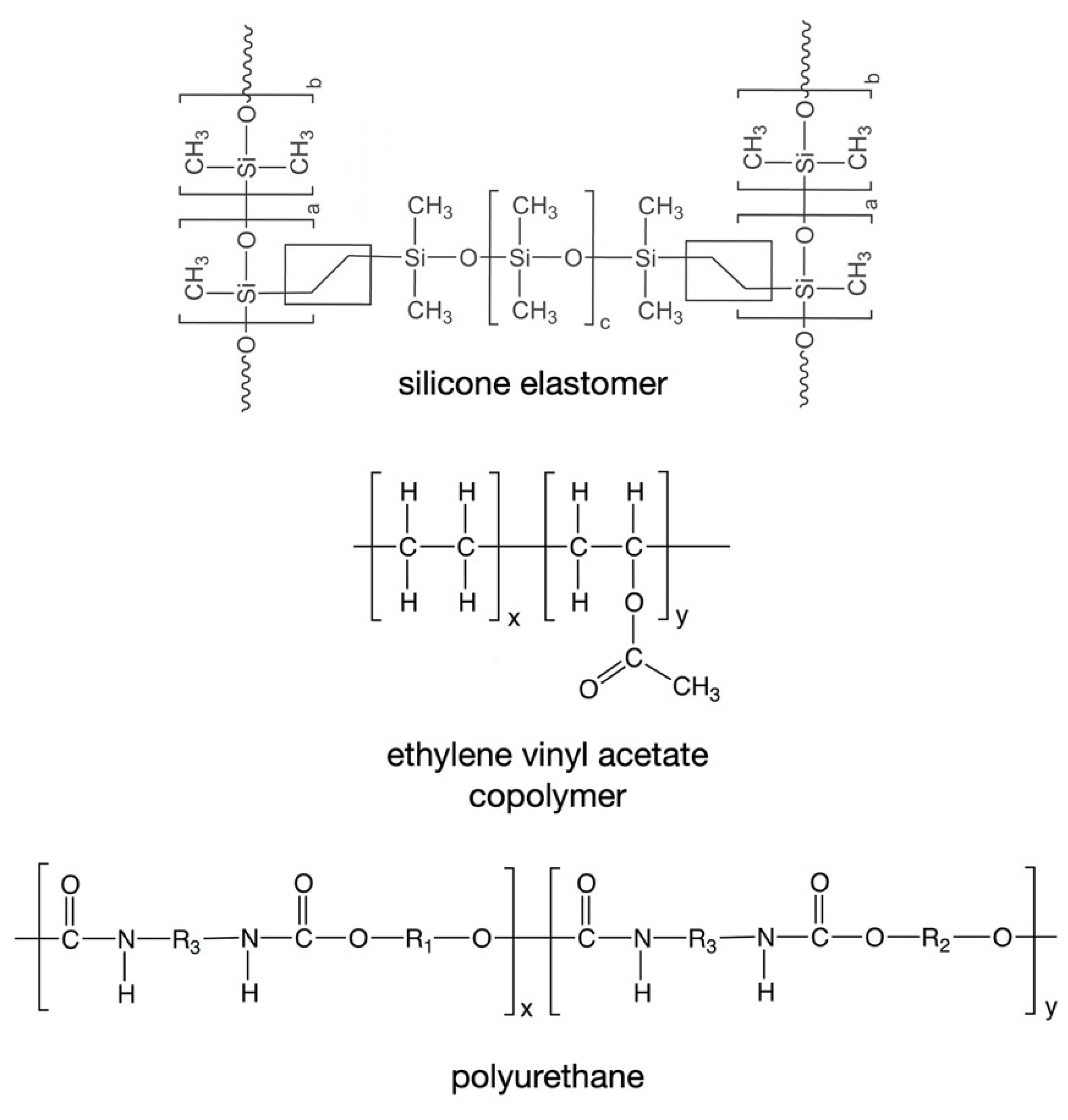
5. Polymeric Materials Used to Manufacture Vaginal Rings
5.1. Silicone Elastomers
5.2. Ethylene Vinyl Acetate Copolymers
5.3. Thermoplastic Polyurethanes
6. Surface Morphology Characteristics of Vaginal Rings
6.1. Method of Manufacture: Injection Molding, Extrusion, Casting and 3D Printing
6.2. Drug Particle Size and Drug Loading
6.3. Extent of Drug Release
7. Biomass Accumulation and Biofilm Formation on Vaginal Rings
7.1. NuvaRing®
7.2. Ornibel®
7.3. Silicone Elastomer Vaginal Rings
7.4. Ring Discoloration
8. Regulatory Considerations
9. Conclusions
Funding
Conflicts of Interest
References
- Malcolm, R.K.; Edwards, K.L.; Kiser, P.; Romano, J.; Smith, T.J. Advances in microbicide vaginal rings. Antivir. Res. 2010, 88, 30–39. [Google Scholar] [CrossRef]
- Brache, V.; Faundes, A. Contraceptive vaginal rings: A review. Contraception 2010, 82, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Brache, V.; Payán, L.J.; Faundes, A. Current status of contraceptive vaginal rings. Contraception 2013, 87, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, R.K.; Boyd, P.J.; McCoy, C.F.; Murphy, D.J. Microbicide vaginal rings: Technological challenges and clinical development. Adv. Drug Deliv. Rev. 2016, 103, 33–56. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.; Variano, B.; Spence, P.; McCoy, C.F.; Murphy, D.J.; Dallal Bashi, Y.H.; Malcolm, R.K. In vitro release testing methods for drug-releasing vaginal rings. J. Control. Release 2019, 313, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, I.; Guazzelli, C.F.; Bahamondes, L. Advances in contraceptive vaginal rings: What does the future hold? Expert Opin. Pharmacother. 2018, 19, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Baeten, J.M.; Palanee-Phillips, T.; Brown, E.R.; Schwartz, K.; Soto-Torres, L.E.; Govender, V.; Mgodi, N.M.; Matovu Kiweewa, F.; Nair, G.; Mhlanga, F.; et al. Use of a vaginal ring containing dapivirine for HIV-1 prevention in women. N. Engl. J. Med. 2016, 375, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.Y.; Zhang, J.; Anderson, P.L.; Wagner, T.; Pan, Z.; Peda, M.; Gomez, K.; Beamer, M.; Jacobson, C.; Strizki, J.; et al. Phase 1 pharmacokinetic trial of 2 intravaginal rings containing different dose strengths of vicriviroc (MK-4176) and MK-2048. Clin. Infect. Dis. 2019, 68, 1129–1135. [Google Scholar] [CrossRef]
- Ferring Pharmaceuticals Randomized Trial Assessing Quinagolide Vaginal Ring for Endometriosis-Related Pain (RAQUEL). ClinicalTrials.gov [Internet]; National Library of Medicine (US): Bethesda, MD, USA, Identifier NCT03692403. Available online: https://clinicaltrials.gov/ct2/show/NCT03692403 (accessed on 10 March 2020).
- Nel, A.; van Niekerk, N.; Kapiga, S.; Bekker, L.-G.; Gama, C.; Gill, K.; Kamali, A.; Kotze, P.; Louw, C.; Mabude, Z.; et al. Safety and efficacy of a dapivirine vaginal ring for HIV prevention in women. N. Engl. J. Med. 2016, 375, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.W.; Karl Malcolm, R.; Dias, N.; Cameron, D.; Offord, R.E.; Hartley, O.; Kett, V.L.; Devlin, B.; Boyd, P. Development and pharmacokinetics of a combination vaginal ring for sustained release of dapivirine and the protein microbicide 5P12-RANTES. Int. J. Pharm. 2019, 564, 207–213. [Google Scholar] [CrossRef] [Green Version]
- McBride, J.W.; Boyd, P.; Dias, N.; Cameron, D.; Offord, R.E.; Hartley, O.; Kett, V.L.; Malcolm, R.K. Vaginal rings with exposed cores for sustained delivery of the HIV CCR5 inhibitor 5P12-RANTES. J. Control. Release 2019, 298, 1–11. [Google Scholar] [CrossRef]
- Nelson, A.L. Comprehensive overview of the recently FDA-approved contraceptive vaginal ring releasing segesterone acetate and ethinylestradiol: A new year-long, patient controlled, reversible birth control method. Expert Rev. Clin. Pharmacol. 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Temmerman, M. A new woman-controlled contraceptive vaginal ring: A global step forward. Lancet Glob. Health 2019, 7, e986–e987. [Google Scholar] [CrossRef] [Green Version]
- Micks, E.A.; Jensen, J.T. A technology evaluation of Annovera: A segesterone acetate and ethinyl estradiol vaginal ring used to prevent pregnancy for up to one year. Expert Opin. Drug Deliv. 2020, 17, 743–752. [Google Scholar] [CrossRef]
- Zhao, C.; Gunawardana, M.; Villinger, F.; Baum, M.M.; Remedios-Chan, M.; Moench, T.R.; Zeitlin, L.; Whaley, K.J.; Bohorov, O.; Smith, T.J.; et al. Pharmacokinetics and preliminary safety of pod-intravaginal rings delivering the monoclonal antibody VRC01-N for HIV prophylaxis in a macaque model. Antimicrob. Agents Chemother. 2017, 61, e02465-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, P.F.; Mann, J.F.S.; Pattani, A.; Kett, V.; Aldon, Y.; King, D.; Malcolm, R.K.; Shattock, R.J. Intravaginal immunisation using a novel antigen-releasing ring device elicits robust vaccine antigen-specific systemic and mucosal humoral immune responses. J. Control. Release 2017, 249, 74–83. [Google Scholar] [CrossRef]
- Kimball, A.B.; Javorsky, E.; Ron, E.S.; Crowley, W.; Langer, R. A novel approach to administration of peptides in women: Systemic absorption of a GnRH agonist via transvaginal ring delivery system. J. Control. Release 2016, 233, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.M.; Moss, J.A.; Srinivasan, P.; Butkyavichene, I.; Gunawardana, M.; Fanter, R.; Miller, C.S.; Sanchez, D.; Yang, F.; Ellis, S.; et al. Novel multipurpose pod-intravaginal ring for the prevention of HIV, HSV, and unintended pregnancy: Pharmacokinetic evaluation in a macaque model. PLoS ONE 2017, 12, e0185946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.T.; Clark, M.R.; Shelke, N.B.; Johnson, T.J.; Smith, E.M.; Andreasen, A.K.; Nebeker, J.S.; Fabian, J.; Friend, D.R.; Kiser, P.F. Engineering a segmented dual-reservoir polyurethane intravaginal ring for simultaneous prevention of HIV transmission and unwanted pregnancy. PLoS ONE 2014, 9, e88509. [Google Scholar] [CrossRef]
- Bash, K.L. Review of vaginal pessaries. Obstet. Gynecol. Surv. 2000, 55, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.A.; Harmanli, O. Pessary use in pelvic organ prolapse and urinary incontinence. Rev. Obstet. Gynecol. 2010, 3, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Vierhout, M.E. The use of pessaries in vaginal prolapse. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 117, 4–9. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amabebe, E.; Anumba, D.O.C. The vaginal microenvironment: The physiologic role of lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Fettweis, J.M.; Brooks, J.P.; Jefferson, K.K.; Buck, G.A. The changing landscape of the vaginal microbiome. Clin. Lab. Med. 2014, 34, 747–761. [Google Scholar] [CrossRef] [Green Version]
- Hardy, L.; Jespers, V.; De Baetselier, I.; Buyze, J.; Mwambarangwe, L.; Musengamana, V.; Van De Wijgert, J.; Crucitti, T. Association of vaginal dysbiosis and biofilm with contraceptive vaginal ring biomass in African women. PLoS ONE 2017, 12, e0178324. [Google Scholar] [CrossRef]
- Sailer, M.; Colli, E.; Regidor, P. In vitro evaluation of microbial adhesion to a contraceptive vaginal ring with a new polymer composition. Eur. J. Contracept. Reprod. Health Care 2019, 24, 188–191. [Google Scholar] [CrossRef]
- Thurman, A.R.; Schwartz, J.L.; Ravel, J.; Gajer, P.; Marzinke, M.A.; Yousefieh, N.; Anderson, S.M.; Doncel, G.F. Vaginal microbiota and mucosal pharmacokinetics of tenofovir in healthy women using tenofovir and tenofovir/levonorgestrel vaginal rings. PLoS ONE 2019, 14, e0217229. [Google Scholar] [CrossRef] [PubMed]
- Camacho, D.P.; Consolaro, M.E.L.; Patussi, E.V.; Donatti, L.; Gasparetto, A.; Svidzinski, T.I.E.; Pereira, D.; Consolaro, M.E.L.; Valéria, E.; Donatti, L.; et al. Vaginal yeast adherence to the combined contraceptive vaginal ring (CCVR). Contraception 2007, 76, 439–443. [Google Scholar] [CrossRef]
- Gunawardana, M.; Moss, J.A.; Smith, T.J.; Kennedy, S.; Kopin, E.; Nguyen, C.; Malone, A.M.; Rabe, L.; Schaudinn, C.; Webster, P.; et al. Microbial biofilms on the surface of intravaginal rings worn in non-human primates. J. Med. Microbiol. 2011, 60, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.; MacFarlane, S.A.; Materi, H.L. A scanning electron microscopic study of the contraceptive vaginal ring. Contraception 2005, 71, 65–67. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, L.; Cerca, N.; Jespers, V.; Vaneechoutte, M.; Crucitti, T. Bacterial biofilms in the vagina. Res. Microbiol. 2017, 168, 865–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crucitti, T.; Hardy, L.; van de Wijgert, J.; Agaba, S.; Buyze, J.; Kestelyn, E.; Delvaux, T.; Mwambarangwe, L.; De Baetselier, I.; Jespers, V. Contraceptive rings promote vaginal lactobacilli in a high bacterial vaginosis prevalence population: A randomised, open-label longitudinal study in Rwandan women. PLoS ONE 2018, 13, e0201003. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Merkatz, R.B.; Hillier, S.L.; Roberts, K.; Blithe, D.L.; Sitruk-Ware, R.; Creinin, M.D. Effects of a one year reusable contraceptive vaginal ring on vaginal microflora and the risk of vaginal infection: An open-label prospective evaluation. PLoS ONE 2015, 10, e0134460. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, S.; Greenbaum, G.; Moran-Gilad, J.; Weintruab, A.Y. Ecological dynamics of the vaginal microbiome in relation to health and disease. Am. J. Obstet. Gynecol. 2019, 220, 324–335. [Google Scholar] [CrossRef]
- Martin, D.H.; Marrazzo, J.M. The vaginal microbiome: Current understanding and future directions. J. Infect. Dis. 2016, 214, S36–S41. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.A.; Miller, K.W.; Sobel, J.D. Dynamics of the vaginal ecosystem—Hormonal influences. Infect. Dis. Res. Treat. 2010, 3. [Google Scholar] [CrossRef] [Green Version]
- Whitney, B.M.; Guthrie, B.L.; Srinivasan, S.; Tapia, K.; Muriuki, E.M.; Chohan, B.H.; Wallis, J.M.; Liu, C.; McClelland, R.S.; Fredricks, D.N.; et al. Changes in key vaginal bacteria among postpartum African women initiating intramuscular depot-medroxyprogesterone acetate. PLoS ONE 2020, 15, e0229586. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S.; Tosh, P.K. A clinician’s primer on the role of the microbiome in human health and disease. Mayo Clin. Proc. 2014, 89, 107–114. [Google Scholar] [CrossRef] [Green Version]
- The Human Microbiome Project Consortium Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [CrossRef] [PubMed] [Green Version]
- Gliniewicz, K.; Schneider, G.M.; Ridenhour, B.J.; Williams, C.J.; Song, Y.; Farage, M.A.; Miller, K.; Forney, L.J. Comparison of the vaginal microbiomes of premenopausal and postmenopausal women. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noyes, N.; Cho, K.C.; Ravel, J.; Forney, L.J.; Abdo, Z. Associations between sexual habits, menstrual hygiene practices, demographics and the vaginal microbiome as revealed by Bayesian network analysis. PLoS ONE 2018, 13, e0191625. [Google Scholar] [CrossRef] [Green Version]
- Larsen, B.; Monif, G.R.G. Understanding the bacterial flora of the female genital tract. Clin. Infect. Dis. 2001, 32, e69–e77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.A.; Beasley, D.A.E.; Dunn, R.R.; Archie, E.A. Lactobacilli dominance and vaginal pH: Why is the human vaginal microbiome unique? Front. Microbiol. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.; Linhares, I. Why do lactobacilli dominate the human vaginal microbiota? BJOG Int. J. Obstet. Gynaecol. 2017, 124, 606–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroutcheva, A.; Gariti, D.; Simon, M.; Shott, S.; Faro, J.; Simoes, J.A.; Gurguis, A.; Faro, S. Defense factors of vaginal lactobacilli. Am. J. Obstet. Gynecol. 2001, 185, 375–379. [Google Scholar] [CrossRef]
- Vásquez, A.; Jakobsson, T.; Ahrné, S.; Forsum, U.; Molin, G. Vaginal lactobacillus flora of healthy Swedish women. J. Clin. Microbiol. 2002, 40, 2746–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wijgert, J.H.H.M.; Jespers, V. The global health impact of vaginal dysbiosis. Res. Microbiol. 2017, 168, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Gardner, H.L.; Dukes, C.D. Haemophilus vaginalis vaginitis. Am. J. Obstet. Gynecol. 1955, 69, 962–976. [Google Scholar] [CrossRef]
- Castro, J.; Machado, D.; Cerca, N. Unveiling the role of Gardnerella vaginalis in polymicrobial Bacterial Vaginosis biofilms: The impact of other vaginal pathogens living as neighbors. ISME J. 2019, 13, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Mikamo, H.; Sato, Y.; Hayasaki, Y.; Hua, Y.X.; Tamaya, T. Vaginal microflora in healthy women with Gardnerella vaginalis. J. Infect. Chemother. 2000, 6, 173–177. [Google Scholar] [CrossRef]
- Fredricks, D.N.; Fiedler, T.L.; Thomas, K.K.; Oakley, B.B.; Marrazzo, J.M. Targeted PCR for detection of vaginal bacteria associated with bacterial vaginosis. J. Clin. Microbiol. 2007, 45, 3270–3276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zozaya-Hinchliffe, M.; Lillis, R.; Martin, D.H.; Ferris, M.J. Quantitative PCR assessments of bacterial species in women with and without bacterial vaginosis. J. Clin. Microbiol. 2010, 48, 1812–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janulaitiene, M.; Paliulyte, V.; Grinceviciene, S.; Zakareviciene, J.; Vladisauskiene, A.; Marcinkute, A.; Pleckaityte, M. Prevalence and distribution of Gardnerella vaginalis subgroups in women with and without bacterial vaginosis. BMC Infect. Dis. 2017, 17, 394. [Google Scholar] [CrossRef]
- Vaneechoutte, M.; Guschin, A.; Van Simaey, L.; Gansemans, Y.; Van Nieuwerburgh, F.; Cools, P. Emended description of Gardnerella vaginalis and description of Gardnerella leopoldii sp. nov., Gardnerella piotii sp. nov. and Gardnerella swidsinskii sp. nov., with delineation of 13 genomic species within the genus Gardnerella. Int. J. Syst. Evol. Microbiol. 2019, 69, 679–687. [Google Scholar] [CrossRef]
- Hill, J.E.; Albert, A.Y.K. Resolution and Cooccurrence Patterns of Gardnerella leopoldii, G. swidsinskii, G. piotii, and G. vaginalis within the Vaginal Microbiome. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef]
- Janulaitiene, M.; Gegzna, V.; Baranauskiene, L.; Bulavaitė, A.; Simanavicius, M.; Pleckaityte, M. Phenotypic characterization of Gardnerella vaginalis subgroups suggests differences in their virulence potential. PLoS ONE 2018, 13, e0200625. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Achilles, S.L.; Bradshaw, C.S.; Burgener, A.; Crucitti, T.; Fredricks, D.N.; Jaspan, H.B.; Kaul, R.; Kaushic, C.; Klatt, N.; et al. The evolving facets of bacterial vaginosis: Implications for HIV transmission. AIDS Res. Hum. Retrovir. 2019, 35, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atashili, J.; Poole, C.; Ndumbe, P.M.; Adimora, A.A.; Smith, J.S. Bacterial vaginosis and HIV acquisition: A meta-analysis of published studies. AIDS 2008, 22, 1493–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, N.; Chersich, M.F.; Schmidlin, K.; Egger, M.; Francis, S.C.; van de Wijgert, J.H.H.M.; Hayes, R.J.; Baeten, J.M.; Brown, J.; Delany-Moretlwe, S.; et al. Intravaginal practices, bacterial vaginosis, and HIV infection in women: Individual participant data meta-analysis. PLoS Med. 2011, 8, e1000416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, C.R.; Lingappa, J.R.; Baeten, J.M.; Ngayo, M.O.; Spiegel, C.A.; Hong, T.; Donnell, D.; Celum, C.; Kapiga, S.; Delany, S.; et al. Bacterial vaginosis associated with increased risk of female-to-male HIV-1 transmission: A prospective cohort analysis among african couples. PLoS Med. 2012, 9, e1001251. [Google Scholar] [CrossRef] [Green Version]
- Verstraete, G.; Vandenbussche, L.; Kasmi, S.; Nuhn, L.; Brouckaert, D.; Van Renterghem, J.; Grymonpré, W.; Vanhoorne, V.; Coenye, T.; De Geest, B.G.G.; et al. Thermoplastic polyurethane-based intravaginal rings for prophylaxis and treatment of (recurrent) bacterial vaginosis. Int. J. Pharm. 2017, 529, 218–226. [Google Scholar] [CrossRef]
- Pathak, M.; Turner, M.; Palmer, C.; Coombes, A.G. Evaluation of polycaprolactone matrices for the intravaginal delivery of metronidazole in the treatment of bacterial vaginosis. J. Biomater. Appl. 2014, 29, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Verhelst, R.; Verstraelen, H.; Claeys, G.; Verschraegen, G.; Van Simaey, L.; De Ganck, C.; De Backer, E.; Temmerman, M.; Vaneechoutte, M. Comparison between Gram stain and culture for the characterization of vaginal microflora: Definition of a distinct grade that resembles grade I microflora and revised categorization of grade I microflora. BMC Microbiol. 2005, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Onderdonk, A.B.; Delaney, M.L.; Fichorova, R.N. The Human Microbiome during Bacterial Vaginosis. Clin. Microbiol. Rev. 2016, 29, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Si, J.; You, H.J.; Yu, J.; Sung, J.; Ko, G. Prevotella as a hub for vaginal microbiota under the influence of host genetics and their association with obesity. Cell Host Microbe 2017, 21, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The gut microbiome profile in obesity: A systematic review. Int. J. Endocrinol. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Cauchie, M.; Desmet, S.; Lagrou, K. Candida and its dual lifestyle as a commensal and a pathogen. Res. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bradford, L.L.; Ravel, J. The vaginal mycobiome: A contemporary perspective on fungi in women’s health and diseases. Virulence 2017, 8, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harriott, M.M.; Lilly, E.A.; Rodriguez, T.E.; Fidel, P.L.; Noverr, M.C. Candida albicans forms biofilms on the vaginal mucosa. Microbiology 2010, 156, 3635–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-B.; Xu, S.-R.; He, Y.; Deng, G.-H.; Sheng, H.-F.; Huang, X.-M.; Ouyang, C.-Y.; Zhou, H.-W. Diverse vaginal microbiomes in reproductive-age women with vulvovaginal candidiasis. PLoS ONE 2013, 8, e79812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrici, A.T. Studies of Freshwater Bacteria: I. A direct microscopic technique. J. Bacteriol. 1933, 25, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zobell, C.E.; Allen, E.C. Attachment of marine bacteria to submerged slides. Exp. Biol. Med. 1933, 30, 1409–1411. [Google Scholar] [CrossRef]
- Zobell, C.E.; Allen, E.C. The significance of marine bacteria in the fouling of submerged surfaces. J. Bacteriol. 1935, 29, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Geesey, G.G.; Cheng, K.-J. How bacteria stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef]
- Geesey, G.G.; Mutch, R.; Costerton, J.W.; Green, R.B. Sessile bacteria: An important component of the microbial population in small mountain streams 1. Limnol. Oceanogr. 1978, 23, 1214–1223. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Leccese Terraf, M.C.; Juárez Tomás, M.S.; Rault, L.; Le Loir, Y.; Even, S.; Nader-Macías, M.E.F. Biofilms of vaginal Lactobacillus reuteri CRL 1324 and Lactobacillus rhamnosus CRL 1332: Kinetics of formation and matrix characterization. Arch. Microbiol. 2016, 198, 689–700. [Google Scholar] [CrossRef]
- Walencka, E.; Różalska, S.; Sadowska, B.; Różalska, B. The influence of Lactobacillus acidophilus-derived surfactants on staphylococcal adhesion and biofilm formation. Folia Microbiol. 2008, 53, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Tahmourespour, A.; Kasra-Kermanshahi, R.; Salehi, R. Lactobacillus rhamnosus biosurfactant inhibits biofilm formation and gene expression of caries-inducing Streptococcus mutans. Dent. Res. J. 2019, 16, 87. [Google Scholar] [CrossRef]
- Salas-Jara, M.; Ilabaca, A.; Vega, M.; García, A. Biofilm forming lactobacillus: New challenges for the development of probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-S.; Ehlers, M.M.; Lombaard, H.; Redelinghuys, M.J.; Kock, M.M. Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 2017, 43, 651–667. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.; Cerca, N. Influence of biofilm formation by gardnerella vaginalis and other anaerobes on bacterial vaginosis. J. Infect. Dis. 2015, 212, 1856–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillan, A.; Macklaim, J.M.; Burton, J.P.; Reid, G. Adhesion of lactobacillus iners AB-1 to human fibronectin. Reprod. Sci. 2013, 20, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Peeters, M.; Piot, P. Adhesion of Gardnerella vaginalis to vaginal epithelial cells: Variables affecting adhesion and inhibition by metronidazole. Sex. Transm. Infect. 1985, 61, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Leccese Terraf, M.C.; Mendoza, L.M.; Juárez Tomás, M.S.; Silva, C.; Nader-Macías, M.E.F. Phenotypic surface properties (aggregation, adhesion and biofilm formation) and presence of related genes in beneficial vaginal lactobacilli. J. Appl. Microbiol. 2014, 117, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Harwich, M.D.; Alves, J.M.; Buck, G.A.; Strauss, J.F.; Patterson, J.L.; Oki, A.T.; Girerd, P.H.; Jefferson, K.K. Drawing the line between commensal and pathogenic Gardnerella vaginalis through genome analysis and virulence studies. BMC Genom. 2010, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Yeoman, C.J.; Yildirim, S.; Thomas, S.M.; Durkin, A.S.; Torralba, M.; Sutton, G.; Buhay, C.J.; Ding, Y.; Dugan-Rocha, S.P.; Muzny, D.M.; et al. Comparative genomics of Gardnerella vaginalis strains reveals substantial differences in metabolic and virulence potential. PLoS ONE 2010, 5, e12411. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Henriques, A.; Machado, A.; Henriques, M.; Jefferson, K.K.; Cerca, N. Reciprocal interference between lactobacillus spp. and gardnerella vaginalis on initial adherence to epithelial cells. Int. J. Med. Sci. 2013, 10, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.; Jefferson, K.; Cerca, N. Interactions between Lactobacillus crispatus and Bacterial Vaginosis (BV)-associated bacterial species in initial attachment and biofilm formation. Int. J. Mol. Sci. 2013, 14, 12004–12012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.; Turner, R.; Marques, L.; Ceri, H. Biofilms. Am. Sci. 2005, 93, 508. [Google Scholar] [CrossRef]
- Muli, F.W.; Struthers, J.K. The growth of Gardnerella vaginalis and Lactobacillus acidophilus in Sorbarod biofilms. J. Med. Microbiol. 1998, 47, 401–405. [Google Scholar] [CrossRef] [Green Version]
- Hymes, S.R.; Randis, T.M.; Sun, T.Y.; Ratner, A.J. DNase inhibits gardnerella vaginalis biofilms in vitro and in vivo. J. Infect. Dis. 2013, 207, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Patterson, J.L.; Stull-Lane, A.; Girerd, P.H.; Jefferson, K.K. Analysis of adherence, biofilm formation and cytotoxicity suggests a greater virulence potential of Gardnerella vaginalis relative to other bacterial-vaginosis-associated anaerobes. Microbiology 2010, 156, 392–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.; Machado, D.; Cerca, N. Escherichia coli and Enterococcus faecalis are able to incorporate and enhance a pre-formed Gardnerella vaginalis biofilm. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, C.W.; Lux, R.; Haake, S.K.; Shi, W. The Fusobacterium nucleatum outer membrane protein RadD is an arginine-inhibitable adhesin required for inter-species adherence and the structured architecture of multispecies biofilm. Mol. Microbiol. 2009, 71, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, T.; Kokubu, E.; Kawana, T.; Saito, A.; Okuda, K.; Ishihara, K. Synergy in biofilm formation between fusobacterium nucleatum and prevotella species. Anaerobe 2012. [Google Scholar] [CrossRef] [Green Version]
- Cassini, M.A.; Pilloni, A.; Condò, S.G.; Vitali, L.A.; Pasquantonio, G.; Cerroni, L. Periodontal bacteria in the genital tract: Are they related to adverse pregnancy outcome? Int. J. Immunopathol. Pharmacol. 2013, 26, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Cerca, N. BV and non-BV associated Gardnerella vaginalis establish similar synergistic interactions with other BV-associated microorganisms in dual-species biofilms. Anaerobe 2015, 36, 56–59. [Google Scholar] [CrossRef] [Green Version]
- Whitchurch, C.B. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Lewis, A.L.; Lewis, W.G. Host sialoglycans and bacterial sialidases: A mucosal perspective. Cell. Microbiol. 2012, 14, 1174–1182. [Google Scholar] [CrossRef]
- Olmsted, S.S.; Meyn, L.A.; Rohan, L.C.; Hillier, S.L. Glycosidase and proteinase activity of anaerobic gram-negative bacteria isolated from women with bacterial vaginosis. Sex. Transm. Dis. 2003, 30, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Cattò, C.; Cappitelli, F. Testing anti-biofilm polymeric surfaces: Where to start? Int. J. Mol. Sci. 2019, 20, 3794. [Google Scholar] [CrossRef] [Green Version]
- von Eiff, C.; Peters, G. Pathogenesis and detection of biofilm formation, on medical implants. In Medical Biofilms; John Wiley & Sons, Ltd.: Chichester, UK, 2005; pp. 51–72. [Google Scholar]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, C.R.; Parsek, M.R. New insight into the early stages of biofilm formation. Proc. Natl. Acad. Sci. USA 2018, 115, 4317–4319. [Google Scholar] [CrossRef] [Green Version]
- Ammar, Y.; Swailes, D.; Bridgens, B.; Chen, J. Influence of surface roughness on the initial formation of biofilm. Surf. Coat. Technol. 2015, 284, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Dunne, W.M. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, C.G.; Anand, S. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Gilmore, B.F.; Carson, L. Bioactive biomaterials for controlling biofilms. In Biomaterials and Medical Device-Associated Infections; Barnes, L., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 163–183. [Google Scholar]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.-F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, M.; Monteiro, F.J.; Ferraz, M.P. Infection of orthopedic implants with emphasis on bacterial adhesion process and techniques used in studying bacterial-material interactions. Biomatter 2012, 2, 176–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnnasouri, M.; Lemaitre, C.; Gentric, C.; Dagot, C.; Pons, M.-N. Influence of surface topography on biofilm development: Experiment and modeling. Biochem. Eng. J. 2011, 57, 38–45. [Google Scholar] [CrossRef]
- Costerton, W.; Veeh, R.; Shirtliff, M.; Pasmore, M.; Post, C.; Ehrlich, G. The application of biofilm science to the study and control of chronic bacterial infections. J. Clin. Investig. 2003, 112, 1466–1477. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Sarkar, S.; Das, B.; Bhattacharjee, S.; Tribedi, P. Biofilm, pathogenesis and prevention—A journey to break the wall: A review. Arch. Microbiol. 2016, 198, 1–15. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Escaping the biofilm in more than one way: Desorption, detachment or dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Toba, F.A.; Akashi, H.; Arrecubieta, C.; Lowy, F.D. Role of biofilm in Staphylococcus aureus and Staphylococcus epidermidis ventricular assist device driveline infections. J. Thorac. Cardiovasc. Surg. 2011, 141, 1259–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, A.; Mashak, A. Review on rubbers in medicine: Natural, silicone and polyurethane rubbers. Plast. Rubber Compos. 2013, 42, 223–230. [Google Scholar] [CrossRef]
- Mashmak, A.; Rahimi, A. Silicone polymers in controlled drug delivery systems: A review. Polym. J. 2009, 18, 279–295. [Google Scholar]
- Caruso, S.; Panella, M.; Giunta, G.; Matarazzo, M.G.; Cianci, A. Comparative randomized study on the sexual function and quality of life of women on contraceptive vaginal ring containing ethinylestradiol/etonogestrel 3.47/11.00 mg or 2.7/11.7 mg. Gynecol. Endocrinol. 2019, 35, 899–903. [Google Scholar] [CrossRef]
- Algorta, J.; Diaz, M.; de Benito, R.; Lefebvre, M.; Sicard, E.; Furtado, M.; Regidor, P.A.; Ronchi, C. Pharmacokinetic bioequivalence, safety and acceptability of Ornibel®, a new polymer composition contraceptive vaginal ring (etonogestrel/ethinylestradiol 11.00/3.474 mg) compared with Nuvaring® (etonogestrel/ethinylestradiol 11.7/2.7 mg). Eur. J. Contracept. Reprod. Health Care 2017, 22, 429–438. [Google Scholar] [CrossRef] [Green Version]
- McConville, C.; Andrews, G.P.; Laverty, T.P.; Woolfson, A.D.; Malcolm, R.K. Rheological evaluation of the isothermal cure characteristics of medical grade silicone elastomers. J. Appl. Polym. Sci. 2010, 116, 2320–2327. [Google Scholar] [CrossRef]
- Dallal Bashi, Y.H.; McCoy, C.F.; Murphy, D.J.; Boyd, P.; Spence, P.; Kleinbeck, K.; Devlin, B.; Malcolm, R.K. Towards a dapivirine and levonorgestrel multipurpose vaginal ring: Investigations into the reaction between levonorgestrel and addition-cure silicone elastomers. Int. J. Pharm. 2019, 569, 118574. [Google Scholar] [CrossRef]
- Murphy, D.J.; Boyd, P.; McCoy, C.F.; Kumar, S.; Holt, J.D.S.; Blanda, W.; Brimer, A.N.; Malcolm, R.K. Controlling levonorgestrel binding and release in a multi-purpose prevention technology vaginal ring device. J. Control. Release 2016, 226, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Schopflin, G.; Fuchs, P.; Kolb, K.H. Drug Excipient of Silicone Rubber. U.S. Patent US4230686, 28 October 1980. [Google Scholar]
- McCoy, C.F.; Apperley, D.C.; Variano, B.; Sussman, H.; Loeven, D.; Boyd, P.; Malcolm, R.K. Solid state 13 C NMR spectroscopy provides direct evidence for reaction between ethinyl estradiol and a silicone elastomer vaginal ring drug delivery system. Int. J. Pharm. 2018, 548, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolfson, A.D.; Malcolm, R.K.; Gallagher, R.J. Design of a silicone reservoir intravaginal ring for the delivery of oxybutynin. J. Control. Release 2003, 91, 465–476. [Google Scholar] [CrossRef]
- Nel, A.; Smythe, S.; Young, K.; Malcolm, K.; McCoy, C.; Rosenberg, Z.; Romano, J. Safety and pharmacokinetics of dapivirine delivery from matrix and reservoir intravaginal rings to HIV-negative women. JAIDS J. Acquir. Immune Defic. Syndr. 2009, 51, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.J.; Courtney, J.M. Silicones as biomaterials. Br. Polym. J. 1988, 20, 25–32. [Google Scholar] [CrossRef]
- Habal, M.B. The biologic basis for the clinical application of the silicones. Arch. Surg. 1984, 119, 843. [Google Scholar] [CrossRef]
- Everaert, E.P.J.M.; Mahieu, H.F.; van de Belt-Gritter, B.; Peeters, A.J.G.E.; Verkerke, G.J.; van der Mei, H.C.; Busscher, H.J. Biofilm formation in vivo on perfluoro-alkylsiloxane–modified voice prostheses. Arch. Otolaryngol. Neck Surg. 1999, 125, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fundeanu, I.; Klee, D.; Schouten, A.J.; Busscher, H.J.; van der Mei, H.C. Solvent-free functionalization of silicone rubber and efficacy of PAAm brushes grafted from an amino-PPX layer against bacterial adhesion. Acta Biomater. 2010, 6, 4271–4276. [Google Scholar] [CrossRef]
- Zhou, L.; Tong, Z.; Wu, G.; Feng, Z.; Bai, S.; Dong, Y.; Ni, L.; Zhao, Y. Parylene coating hinders Candida albicans adhesion to silicone elastomers and denture bases resin. Arch. Oral Biol. 2010, 55, 401–409. [Google Scholar] [CrossRef]
- MacCallum, N.; Howell, C.; Kim, P.; Sun, D.; Friedlander, R.; Ranisau, J.; Ahanotu, O.; Lin, J.J.; Vena, A.; Hatton, B.; et al. Liquid-Infused silicone as a biofouling-free medical material. ACS Biomater. Sci. Eng. 2015, 1, 43–51. [Google Scholar] [CrossRef]
- Wang, R.; Neoh, K.G.; Shi, Z.; Kang, E.-T.; Tambyah, P.A.; Chiong, E. Inhibition of escherichia coli and proteus mirabilis adhesion and biofilm formation on medical grade silicone surface. Biotechnol. Bioeng. 2012, 109, 336–345. [Google Scholar] [CrossRef]
- Leonhard, M.; Moser, D.; Reumueller, A.; Mancusi, G.; Bigenzahn, W.; Schneider-Stickler, B. Comparison of biofilm formation on new Phonax and Provox 2 voice prostheses-A pilot study. Head Neck 2009. [Google Scholar] [CrossRef] [PubMed]
- Kurtulmus, H.; Kumbuloglu, O.; Özcan, M.; Ozdemir, G.; Ozcan, M.; Vural, C. Candida albicans adherence on silicone elastomers: Effect of polymerisation duration and exposure to simulated saliva and nasal secretion. Dent. Mater. 2010, 26, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, R.; McCullagh, S.; Woolfson, A.; Gorman, S.; Jones, D.; Cuddy, J. Controlled release of a model antibacterial drug from a novel self-lubricating silicone biomaterial. J. Control. Release 2004, 97, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Park, J.U.; Ham, J.; Kim, S.; Seo, J.; Kim, S.-H.; Lee, S.; Min, H.J.; Choi, S.; Choi, R.M.; Kim, H.; et al. Alleviation of capsular formations on silicone implants in rats using biomembrane-mimicking coatings. Acta Biomater. 2014, 10, 4217–4225. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.C.; Karl Malcolm, R.; David Woolfson, A.; Gorman, S.P. Persistence of antimicrobial activity through sustained release of triclosan from pegylated silicone elastomers. Biomaterials 2009, 30, 6739–6747. [Google Scholar] [CrossRef]
- Woolfson, A.D.; Malcolm, R.K.; Gorman, S.P.; Jones, D.S.; Brown, A.F.; McCullagh, S.D. Self-lubricating silicone elastomer biomaterials. J. Mater. Chem. 2003, 13, 2465. [Google Scholar] [CrossRef]
- Mahieu, H.F.; van Saene, H.K.F.; Rosingh, H.J.; Schutte, H.K. Candida vegetations on silicone voice prostheses. Arch. Otolaryngol. Head Neck Surg. 1986, 112, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Van Den Hoogen, F.J.A.A.; Oudes, M.J.; Hombergen, G.; Nijdam, H.F.; Manni, J.J. The groningen, nijdam and provox voice prostheses: A prospective clinical comparison based on 845 replacements. Acta Otolaryngol. 1996, 116, 119–124. [Google Scholar] [CrossRef]
- Hilgers, F.J.; Balm, A.J. Long-term results of vocal rehabilitation after total laryngectomy with the low-resistance, indwelling Provox voice prosthesis system. Clin. Otolaryngol. Allied Sci. 1993, 18, 517–523. [Google Scholar] [CrossRef]
- Everaert, E.P.J.M.; van de Belt-Gritter, B.; van der Mei, H.C.; Busscher, H.J.; Verkerke, G.J.; Dijk, F.; Mahieu, H.F.; Reitsma, A. In vitro and in vivo microbial adhesion and growth on argon plasma-treated silicone rubber voice prostheses. J. Mater. Sci. Mater. Med. 1998, 9, 147–157. [Google Scholar] [CrossRef]
- Rodrigues, L.; van der Mei, H.; Teixeira, J.A.; Oliveira, R. Biosurfactant from Lactococcus lactis 53 inhibits microbial adhesion on silicone rubber. Appl. Microbiol. Biotechnol. 2004, 66, 306–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, H.; Martell, B.; Constantine, G.D.; Davis, S.M.; Vidal, J.D.; Mayer, P.R.; Doorbar, M.; Friend, D.R. Pharmacokinetics and tolerability of a novel 17β-Estradiol and progesterone intravaginal ring in sheep. J. Pharm. Sci. 2019, 108, 2677–2684. [Google Scholar] [CrossRef] [PubMed]
- Verstraelen, H.; Vervaet, C.; Remon, J.-P. Rationale and safety assessment of a novel intravaginal drug-delivery system with sustained DL-lactic acid release, intended for long-term protection of the vaginal microbiome. PLoS ONE 2016, 11, e0153441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConville, C.; Major, I.; Friend, D.R.; Clark, M.R.; Malcolm, R.K. Development of a UC781 releasing polyethylene vinyl acetate vaginal ring. Drug Deliv. Transl. Res. 2012, 2, 489–497. [Google Scholar] [CrossRef] [PubMed]
- McConville, C.; Major, I.; Friend, D.R.; Clark, M.R.; Woolfson, A.D.; Malcolm, R.K. Development of polylactide and polyethylene vinyl acetate blends for the manufacture of vaginal rings. J. Biomed. Mater. Res. Part B Appl. Biomater. 2012, 100B, 891–895. [Google Scholar] [CrossRef]
- Loxley, A.; Mitchnick, M.; Okoh, O.; McConnell, J.; Goldman, L.; Morgan, C.; Clark, M.; Friend, D.R. Ethylene vinyl acetate intravaginal rings for the simultaneous delivery of the antiretroviral UC781 and contraceptive levonorgestrel. Drug Deliv. Transl. Res. 2011, 1, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Koutsamanis, I.; Paudel, A.; Nickisch, K.; Eggenreich, K.; Roblegg, E.; Eder, S. Controlled-release from high-loaded reservoir-type systems—A case study of ethylene-vinyl acetate and progesterone. Pharmaceutics 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsamanis, I.; Eder, S.; Beretta, M.; Witschnigg, A.; Paudel, A.; Nickisch, K.; Friedrich, M.; Eggenreich, K.; Roblegg, E. Formulation and processability screening for the rational design of ethylene-vinyl acetate based intra-vaginal rings. Int. J. Pharm. 2019, 564, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hoesley, C.J.; Chen, B.A.; Anderson, P.L.; Dezzutti, C.S.; Strizki, J.; Sprinkle, C.; Heard, F.; Bauermeister, J.; Hall, W.; Jacobson, C.; et al. Phase 1 safety and pharmacokinetics study of MK-2048/Vicriviroc (MK-4176)/MK-2048A intravaginal rings. Clin. Infect. Dis. 2019, 68, 1136–1143. [Google Scholar] [CrossRef]
- Giannasca, N.J.; Suon, J.S.; Evans, A.C.; Margulies, B.J. Matrix-based controlled release delivery of acyclovir from poly-(ethylene co-vinyl acetate) rings. J. Drug Deliv. Sci. Technol. 2020, 55, 101391. [Google Scholar] [CrossRef]
- De Oliveira Filho, R.V.; de Jesus Antunes, N.; de Oliveira Ilha, J.; Moreno, R.A.; Wedemeyer, R.-S.; Warnke, A.; De Nucci, G. Pharmacokinetics and pharmacodynamics of three dosages of oestriol after continuous vaginal ring administration for 21 days in healthy, postmenopausal women. Br. J. Clin. Pharmacol. 2019, 85, 551–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.; Langer, R.; Loveday, D.; Hair, D. Applications of ethylene vinyl acetate copolymers (EVA) in drug delivery systems. J. Control. Release 2017, 262, 284–295. [Google Scholar] [CrossRef]
- Tiwari, R.V.; Patil, H.; Repka, M.A. Contribution of hot-melt extrusion technology to advance drug delivery in the 21st century. Expert Opin. Drug Deliv. 2016, 13, 451–464. [Google Scholar] [CrossRef]
- Nostro, A.; Scaffaro, R.; Ginestra, G.; D’Arrigo, M.; Botta, L.; Marino, A.; Bisignano, G. Control of biofilm formation by poly-ethylene-co-vinyl acetate films incorporating nisin. Appl. Microbiol. Biotechnol. 2010, 87, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J.; Maddocks, S.E.; Grady, H.J.; Collins, A.M.; Barbour, M.E. Functionalization of ethylene vinyl acetate with antimicrobial chlorhexidine hexametaphosphate nanoparticles. Int. J. Nanomed. 2014, 9, 4145–4152. [Google Scholar] [CrossRef] [Green Version]
- Engelsman, A.F.; Krom, B.P.; Busscher, H.J.; van Dam, G.M.; Ploeg, R.J.; van der Mei, H.C. Antimicrobial effects of an NO-releasing poly(ethylene vinylacetate) coating on soft-tissue implants in vitro and in a murine model. Acta Biomater. 2009, 5, 1905–1910. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Scaffaro, R.; D’Arrigo, M.; Botta, L.; Filocamo, A.; Marino, A.; Bisignano, G. Development and characterization of essential oil component-based polymer films: A potential approach to reduce bacterial biofilm. Appl. Microbiol. Biotechnol. 2013, 97, 9515–9523. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Yamasaki, S.; Honda, M.; Terao, Y.; Kawabata, S.; Maeda, Y. Long-term survival of salivary streptococci on dental devices made of ethylene vinyl acetate. Int. J. Oral Sci. 2012, 4, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Teno, J.; González-Gaitano, G.; González-Benito, J. Poly (ethylene-co-vinyl acetate) films prepared by solution blow spinning: Surface characterization and its relation with E. coli adhesion. Polym. Test. 2017, 60, 140–148. [Google Scholar] [CrossRef]
- Nostro, A.; Scaffaro, R.; Botta, L.; Filocamo, A.; Marino, A.; Bisignano, G. Effect of temperature on the release of carvacrol and cinnamaldehyde incorporated into polymeric systems to control growth and biofilms of Escherichia coli and Staphylococcus aureus. Biofouling 2015, 31, 639–649. [Google Scholar] [CrossRef]
- Doganci, M.D.; Cansoy, C.E.; Ucar, I.O.; Erbil, H.Y.; Mielczarski, E.; Mielczarski, J.A. Combined XPS and contact angle studies of flat and rough ethylene-vinyl acetate copolymer films. J. Appl. Polym. Sci. 2012, 124, 2100–2109. [Google Scholar] [CrossRef]
- Matsunaga, T.; Tamai, Y. Surface free energy analysis of ethylene-vinyl acetate copolymers. J. Appl. Polym. Sci. 1978, 22, 3525–3530. [Google Scholar] [CrossRef]
- Erbil, H.Y. Surface-free energy analysis of hydrolyzed ethylene-vinyl acetate copolymers. J. Appl. Polym. Sci. 1987. [Google Scholar] [CrossRef]
- Ucar, I.O.; Doganci, M.D.; Cansoy, C.E.; Erbil, H.Y.; Avramova, I.; Suzer, S. Combined XPS and contact angle studies of ethylene vinyl acetate and polyvinyl acetate blends. Appl. Surf. Sci. 2011, 257, 9587–9594. [Google Scholar] [CrossRef] [Green Version]
- Michalski, M.C.; Hardy, J.; Saramago, B.J.V. On the surface free energy of PVC/EVA polymer blends: Comparison of different calculation methods. J. Colloid Interface Sci. 1998, 208, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Erbil, H.Y.; Meriç, R.A. Determination of surface free energy components of polymers from contact angle data using nonlinear programming methods. Colloids Surf. 1988, 33, 85–97. [Google Scholar] [CrossRef]
- Devallencourt, C.; Marais, S.; Saiter, J.M.; Labbé, M.; Métayer, M. Study of transport of small molecules through ethylene-co-vinyl acetate copolymers films. Part A: Water molecules. Polym. Test. 2002, 21, 253–262. [Google Scholar] [CrossRef]
- Johnson, T.J.; Srinivasan, P.; Albright, T.H.; Watson-Buckheit, K.; Rabe, L.; Martin, A.; Pau, C.-P.; Hendry, R.M.; Otten, R.; McNicholl, J.; et al. Safe and sustained vaginal delivery of pyrimidinedione HIV-1 inhibitors from polyurethane intravaginal rings. Antimicrob. Agents Chemother. 2012, 56, 1291–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traore, Y.L.; Chen, Y.; Bernier, A.M.; Ho, E.A. Impact of hydroxychloroquine-loaded polyurethane intravaginal rings on lactobacilli. Antimicrob. Agents Chemother. 2015, 59, 7680–7686. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Gupta, K.M.; Fabian, J.; Albright, T.H.; Kiser, P.F. Segmented polyurethane intravaginal rings for the sustained combined delivery of antiretroviral agents dapivirine and tenofovir. Eur. J. Pharm. Sci. 2010, 39, 203–212. [Google Scholar] [CrossRef]
- Johnson, T.J.; Clark, M.R.; Albright, T.H.; Nebeker, J.S.; Tuitupou, A.L.; Clark, J.T.; Fabian, J.; McCabe, R.T.; Chandra, N.; Doncel, G.F.; et al. A 90-Day Tenofovir Reservoir Intravaginal Ring for Mucosal HIV Prophylaxis. Antimicrob. Agents Chemother. 2012, 56, 6272–6283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.M.; Pearce, S.M.; Poursaid, A.E.; Aliyar, H.A.; Tresco, P.A.; Mitchnik, M.A.; Kiser, P.F. Polyurethane intravaginal ring for controlled delivery of dapivirine, a nonnucleoside reverse transcriptase inhibitor of HIV-1. J. Pharm. Sci. 2008, 97, 4228–4239. [Google Scholar] [CrossRef] [PubMed]
- Welsh, N.R.; Malcolm, R.K.; Devlin, B.; Boyd, P. Dapivirine-releasing vaginal rings produced by plastic freeforming additive manufacturing. Int. J. Pharm. 2019, 572, 118725. [Google Scholar] [CrossRef]
- Teller, R.S.; Malaspina, D.C.; Rastogi, R.; Clark, J.T.; Szleifer, I.; Kiser, P.F. Controlling the hydration rate of a hydrophilic matrix in the core of an intravaginal ring determines antiretroviral release. J. Control. Release 2016, 224, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherng, J.Y.; Hou, T.Y.; Shih, M.F.; Talsma, H.; Hennink, W.E. Polyurethane-based drug delivery systems. Int. J. Pharm. 2013, 450, 145–162. [Google Scholar] [CrossRef]
- Kaur, M.; Gupta, K.M.; Poursaid, A.E.; Karra, P.; Mahalingam, A.; Aliyar, H.A.; Kiser, P.F. Engineering a degradable polyurethane intravaginal ring for sustained delivery of dapivirine. Drug Deliv. Transl. Res. 2011, 1, 223–237. [Google Scholar] [CrossRef]
- Wei, Q.; Becherer, T.; Angioletti-Uberti, S.; Dzubiella, J.; Wischke, C.; Neffe, A.T.; Lendlein, A.; Ballauff, M.; Haag, R. Protein interactions with polymer coatings and biomaterials. Angew. Chem. Int. Ed. 2014, 53, 8004–8031. [Google Scholar] [CrossRef]
- Veerachamy, S.; Yarlagadda, T.; Manivasagam, G.; Yarlagadda, P.K. Bacterial adherence and biofilm formation on medical implants: A review. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2014, 228, 1083–1099. [Google Scholar] [CrossRef]
- Smith, A.W. Biofilms and antibiotic therapy: Is there a role for combating bacterial resistance by the use of novel drug delivery systems? Adv. Drug Deliv. Rev. 2005, 57, 1539–1550. [Google Scholar] [CrossRef]
- Desrousseaux, C.; Sautou, V.; Descamps, S.; Traoré, O. Modification of the surfaces of medical devices to prevent microbial adhesion and biofilm formation. J. Hosp. Infect. 2013, 85, 87–93. [Google Scholar] [CrossRef]
- Buijssen, K.J.D.A.; Oosterhof, J.J.H.; Basil, L.; Waters, M.; Duits, M.A.; Busscher, H.J.; van der Mei, H.C.; van der Laan, B.F.A.M. Influence of surface roughness on silicone rubber voice prostheses on in vitro biofilm formation and clinical lifetime in laryngectomised patients. Clin. Otolaryngol. 2017, 42, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, J. Melt extrusion: From process to drug delivery technology. Eur. J. Pharm. Biopharm. 2002, 54, 107–117. [Google Scholar] [CrossRef]
- Molenaar, J.; Koopmans, R.J.; den Doelder, C.F.J. Onset of the sharkskin phenomenon in polymer extrusion. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 1998. [Google Scholar] [CrossRef]
- Fu, J.; Yu, X.; Jin, Y. 3D printing of vaginal rings with personalized shapes for controlled release of progesterone. Int. J. Pharm. 2018, 539, 75–82. [Google Scholar] [CrossRef]
- Saxena, B.B.; Singh, M.; Gospin, R.M.; Chu, C.C.; Ledger, W.J. Efficacy of nonhormonal vaginal contraceptives from a hydrogel delivery system. Contraception 2004, 70, 213–219. [Google Scholar] [CrossRef]
- Saxena, B.B.; Han, Y.A.; Fu, D.; Rathnam, P.; Singh, M.; Laurence, J.; Lerner, S. Sustained release of microbicides by newly engineered vaginal rings. AIDS 2009, 23, 917–922. [Google Scholar] [CrossRef]
- Saxena, B.B.; Koldras, K.E.; Singh, M.; Nguyen, N.; Premila, R.; Ledger, W.J.; Lerner, S. Development of a nanoporous elastomere intra-vaginal ring (IVR) for the sustained release of non-hormonal contraceptives. J. Pharm. Drug Deliv. Res. 2012, 1, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, R.A.; Behl, S.; Ameer, G.A. Nanoporous biodegradable elastomers. Adv. Mater. 2009, 21, 188–192. [Google Scholar] [CrossRef]
- Yang, J.; Webb, A.R.; Pickerill, S.J.; Hageman, G.; Ameer, G.A. Synthesis and evaluation of poly(diol citrate) biodegradable elastomers. Biomaterials 2006, 27, 1889–1898. [Google Scholar] [CrossRef]
- Rasenack, N.; Müller, B.W. Micron-size drug particles: Common and novel micronization techniques. Pharm. Dev. Technol. 2004, 9, 1–13. [Google Scholar] [CrossRef]
- Van Laarhoven, J.A.H.; Kruft, M.A.B.; Vromans, H. Effect of supersaturation and crystallization phenomena on the release properties of a controlled release device based on EVA copolymer. J. Control. Release 2002, 82, 309–317. [Google Scholar] [CrossRef]
- Almeida, A.; Possemiers, S.; Boone, M.N.; De Beer, T.; Quinten, T.; Van Hoorebeke, L.; Remon, J.P.; Vervaet, C. Ethylene vinyl acetate as matrix for oral sustained release dosage forms produced via hot-melt extrusion. Eur. J. Pharm. Biopharm. 2011, 77, 297–305. [Google Scholar] [CrossRef]
- El Kissi, N.; Piau, J.-M.; Toussaint, F. Sharkskin and cracking of polymer melt extrudates. J. Nonnewton. Fluid Mech. 1997, 68, 271–290. [Google Scholar] [CrossRef]
- Murphy, D.J.; McCoy, C.F.; Plagianos, M.; RamaRao, S.; Merkatz, R.; Clark, H.; Boyd, P.; Variano, B.; Malcolm, R.K. Post-use ring weight and residual drug content as potential objective measures of user adherence to a contraceptive progesterone vaginal ring. Contraception 2019, 100, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Devlin, B.; Nuttall, J.; Wilder, S.; Woodsong, C.; Rosenberg, Z. Development of dapivirine vaginal ring for HIV prevention. Antivir. Res. 2013, 100, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Spence, P.; Nel, A.; van Niekerk, N.; Derrick, T.; Wilder, S.; Devlin, B. Post-use assay of vaginal rings (VRs) as a potential measure of clinical trial adherence. J. Pharm. Biomed. Anal. 2016, 125, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.J.; Malone, A.M.; Carpenter, C.A.; Lo, Y.; Huang, M.; Corey, L.; Willis, R.; Nguyen, C.; Kennedy, S.; Gunawardana, M.; et al. Safety and pharmacokinetics of aciclovir in women following release from a silicone elastomer vaginal ring. J. Antimicrob. Chemother. 2012, 67, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Roumen, F.J.; Apter, D.; Mulders, T.M.; Dieben, T.O. Efficacy, tolerability and acceptability of a novel contraceptive vaginal ring releasing etonogestrel and ethinyl oestradiol. Hum. Reprod. 2001, 16, 469–475. [Google Scholar] [CrossRef]
- De Seta, F.; Restaino, S.; De Santo, D.; Stabile, G.; Banco, R.; Busetti, M.; Barbati, G.; Guaschino, S. Effects of hormonal contraception on vaginal flora. Contraception 2012, 86, 526–529. [Google Scholar] [CrossRef]
- De Seta, F.; Restaino, S.; Banco, R.; Conversano, E.; De Leo, R.; Tonon, M.; Maso, G.; Barbati, G.; Lello, S. Effects of estroprogestins containing natural estrogen on vaginal flora. Gynecol. Endocrinol. 2014, 30, 830–835. [Google Scholar] [CrossRef]
- Veres, S.; Miller, L.; Burington, B. A comparison between the vaginal ring and oral contraceptives. Obstet. Gynecol. 2004, 104, 555–563. [Google Scholar] [CrossRef]
- Kestelyn, E.; Agaba, S.; Van Nuil, J.I.; Uwineza, M.; Umulisa, M.M.; Mwambarangwe, L.; Ndagijimana, J.C.; De Baetselier, I.; Buyze, J.; Delvaux, T.; et al. A randomised trial of a contraceptive vaginal ring in women at risk of HIV infection in Rwanda: Safety of intermittent and continuous use. PLoS ONE 2018, 13, e0197572. [Google Scholar] [CrossRef] [Green Version]
- Davies, G.C.; Feng, L.X.; Newton, J.R.; Dieben, T.O.M.; Coelingh-Bennink, H. The effects of a combined contraceptive vaginal ring releasing ethinyloestradiol and 3-ketodesogestrel on vaginal flora. Contraception 1992, 45, 511–518. [Google Scholar] [CrossRef]
- Oddsson, K.; Leifels-Fischer, B.; de Melo, N.R.; Wiel-Masson, D.; Benedetto, C.; Verhoeven, C.H.J.; Dieben, T.O.M. Efficacy and safety of a contraceptive vaginal ring (NuvaRing) compared with a combined oral contraceptive: A 1-year randomized trial. Contraception 2005, 71, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Chassot, F.; Camacho, D.P.; Patussi, E.V.; Donatti, L.; Svidzinski, T.I.E.; Consolaro, M.E.L. Can Lactobacillus acidophilus influence the adhesion capacity of Candida albicans on the combined contraceptive vaginal ring? Contraception 2010, 81, 331–335. [Google Scholar] [CrossRef]
- Song, F.; Koo, H.; Ren, D. Effects of material properties on bacterial adhesion and biofilm formation. J. Dent. Res. 2015, 94, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G.; Timò, A.; Sammarini, M.; Del Savio, M.C.; Facchinetti, F. Surface roughness of different contraceptive vaginal rings: Evaluation by scanning electron microscope (SEM). Eur. J. Contracept. Reprod. Health Care 2020, 25, 60–64. [Google Scholar] [CrossRef]
- Final Study Report MTN 005 Expanded Safety and Adherence Study of a Non-Medicated Intravaginal Ring; Microbicide Trials Network. 2010. Available online: https://mtnstopshiv.org/sites/default/files/attachments/MTN-005%20Version%202%200_19OCT2010.pdf (accessed on 18 May 2021).
- Ursell, L.K.; Gunawardana, M.; Chang, S.; Mullen, M.; Moss, J.A.; Herold, B.C.; Keller, M.J.; McDonald, D.; González, A.; Knight, R.; et al. Comparison of the vaginal microbial communities in women with recurrent genital HSV receiving acyclovir intravaginal rings. Antivir. Res. 2014, 102, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Stalter, R.M.; Moench, T.R.; MacQueen, K.M.; Tolley, E.E.; Owen, D.H. Biomarkers and biometric measures of adherence to use of ARV-based vaginal rings. J. Int. AIDS Soc. 2016, 19, 20746. [Google Scholar] [CrossRef] [Green Version]
- McCoy, C.F.; Spence, P.; Dallal Bashi, Y.H.; Murphy, D.J.; Boyd, P.; Dangi, B.; Derrick, T.; Devlin, B.; Kleinbeck, K.; Malcolm, R.K. Use of simulated vaginal and menstrual fluids to model in vivo discoloration of silicone elastomer vaginal rings. Int. J. Pharm. X 2021, 3, 100081. [Google Scholar] [CrossRef]
- Fleming, H.-C. Relevance of biofilms for the biodeterioration of surfaces of polymeric materials. Polym. Degrad. Stab. 1997, 59, 309–325. [Google Scholar] [CrossRef]
- Burton, F.G.; Skiens, W.E.; Gordon, N.R.; Veal, J.T.; Kalkwarf, D.R.; Duncan, G.W. Fabrication and testing of vaginal contraceptive devices designed for release of prespecified dose levels of steroids. Contraception 1978, 17, 221–230. [Google Scholar] [CrossRef]
- Ballagh, S.A. Vaginal ring hormone delivery systems in contraception and menopause. Clin. Obstet. Gynecol. 2001, 44, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.J.; McCoy, C.F.; Boyd, P.; Derrick, T.; Spence, P.; Devlin, B.; Malcolm, R.K. Drug stability and product performance characteristics of a dapivirine-releasing vaginal ring under simulated real-world conditions. Int. J. Pharm. 2019, 565, 351–357. [Google Scholar] [CrossRef]
- Phillips, K.S.; Patwardhan, D.; Jayan, G. Biofilms, medical devices, and antibiofilm technology: Key messages from a recent public workshop. Am. J. Infect. Control 2015, 43, 2–3. [Google Scholar] [CrossRef]
- Sarkar, N. The combined contraceptive vaginal device (NuvaRing®): A comprehensive review. Eur. J. Contracept. Reprod. Health Care 2005, 10, 73–78. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vaginal Ring (Company) | Device Type (Duration of Release) | Active Agent(s) (Loading/Release Rate) | Polymer(s) | Indication | Ring Dimensions |
|---|---|---|---|---|---|
| Estring® (Pfizer) | reservoir (3 months) | 17β-estradiol (2 mg/7.5 μg/day) | silicone elastomer core and sheath (both Q7-4735, Dow) | estrogen replacementtherapy | Ring OD: 55 mm Ring CSD: 9.0 mm Core CSD: 2.0 mm Core length: 145 mm |
| NuvaRing® (Merck) EluRyng™ (Amneal) Myring™ (Mithra) Generic (TEVA) | reservoir (21 days) | etonogestrel (11.7 mg/120 μg/day) ethinyl estradiol (2.7 mg/15 μg/day) | 28% EVA * copolymer core and 9% EVA * sheath | combination contraception | Ring OD: 54 mm Ring CSD: 4.0 mm Membrane thickness: 110 μm |
| Femring® (Millicent) | reservoir (3 months) | 17β-estradiol-3-acetate (12.4, 24.8 mg/50, 100 μg/day) | silicone elastomer core and sheath (both MED-6382, NuSil) | estrogen replacement therapy | Ring OD: 56 mm Ring CSD: 7.6 mm Core CSD: 2.0 mm Core lengths: 8 and 16 mm |
| Progering® (Population Council/Silesia SA/Grupo Grünenthal Chile) | matrix (3 months) | progesterone (2074 mg/~10 mg/day) | silicone elastomer (MED-4211, NuSil) | post-partum contraception in breastfeeding women | Ring OD: 56 mm Ring CSD: 8.4 mm |
| Fertiring® (Population Council/Silesia SA/Grupo Grünenthal Chile) | matrix (3 months) | progesterone (1000 mg/~10 mg/day) | silicone elastomer (MED-4211, NuSil) | IVF/hormone supplementation | Ring OD: 56 mm Ring CSD: 8.4 mm |
| Ornibel® (Exeltis) SyreniRing (Crescent Pharma) Kirkos® (Farmitalia) | reservoir (21 days) | etonogestrel (11.0 mg/120 μg/day) ethinyl estradiol (3.47 mg/15 μg/day) | polyurethane sheath and 28% EVA* copolymer core | combination contraception | Ring OD: 54 mm Ring CSD: 4.0 mm Membrane thickness: 150 μm |
| Annovera™ (Population Council) | reservoir (1 year) | segesterone acetate (103 mg/150 μg/day) ethinyl estradiol (17.4 mg/13 μg/day) | silicone elastomer cores (x2, MED-6603 and MED-6385, NuSil) and sheath (MED-4224, NuSil) | combination contraception | Ring OD: 56 mm Ring CSD: 8.4 mm Core CSD: 3.0 mm Core lengths: 11 and 18 mm |
| Gram-Positive Rods | ||
| Diptheroids | +++ | |
| Lactobacilli | +++ | |
| Gram-positive cocci | ||
| Staphylococcus aureus | + | |
| Staphylococcus epidermidis | ++ | |
| Streptococcus species | ||
| α-Hemolytic | + | |
| Β-Hemolytic | + | |
| Non-hemolytic | + | |
| Group D | + | |
| Gram-Negative Rods | ||
| Escherichia coli | + | |
| Klebsiella and Enterobacter spp. | + | |
| Proteus spp. | + | |
| Pseudomonas spp. | + | |
| Anaerobic Species | ||
| Bacteroides spp. | ++ | |
| Bifidobacterium spp. | + | |
| Fuscobacterium spp. | + | |
| Lactobacillus spp. | ++ | |
| Peptococcus spp. | +++ | |
| Preptostreptococcus spp. | +++ | |
| Proprionibacterium spp. | + | |
| Veillonella spp. | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carson, L.; Merkatz, R.; Martinelli, E.; Boyd, P.; Variano, B.; Sallent, T.; Malcolm, R.K. The Vaginal Microbiota, Bacterial Biofilms and Polymeric Drug-Releasing Vaginal Rings. Pharmaceutics 2021, 13, 751. https://doi.org/10.3390/pharmaceutics13050751
Carson L, Merkatz R, Martinelli E, Boyd P, Variano B, Sallent T, Malcolm RK. The Vaginal Microbiota, Bacterial Biofilms and Polymeric Drug-Releasing Vaginal Rings. Pharmaceutics. 2021; 13(5):751. https://doi.org/10.3390/pharmaceutics13050751
Chicago/Turabian StyleCarson, Louise, Ruth Merkatz, Elena Martinelli, Peter Boyd, Bruce Variano, Teresa Sallent, and Robert Karl Malcolm. 2021. "The Vaginal Microbiota, Bacterial Biofilms and Polymeric Drug-Releasing Vaginal Rings" Pharmaceutics 13, no. 5: 751. https://doi.org/10.3390/pharmaceutics13050751