
Comparison of Different Liquid Chromatography-Based Purification Strategies for Adeno-Associated Virus Vectors


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. AAV Production
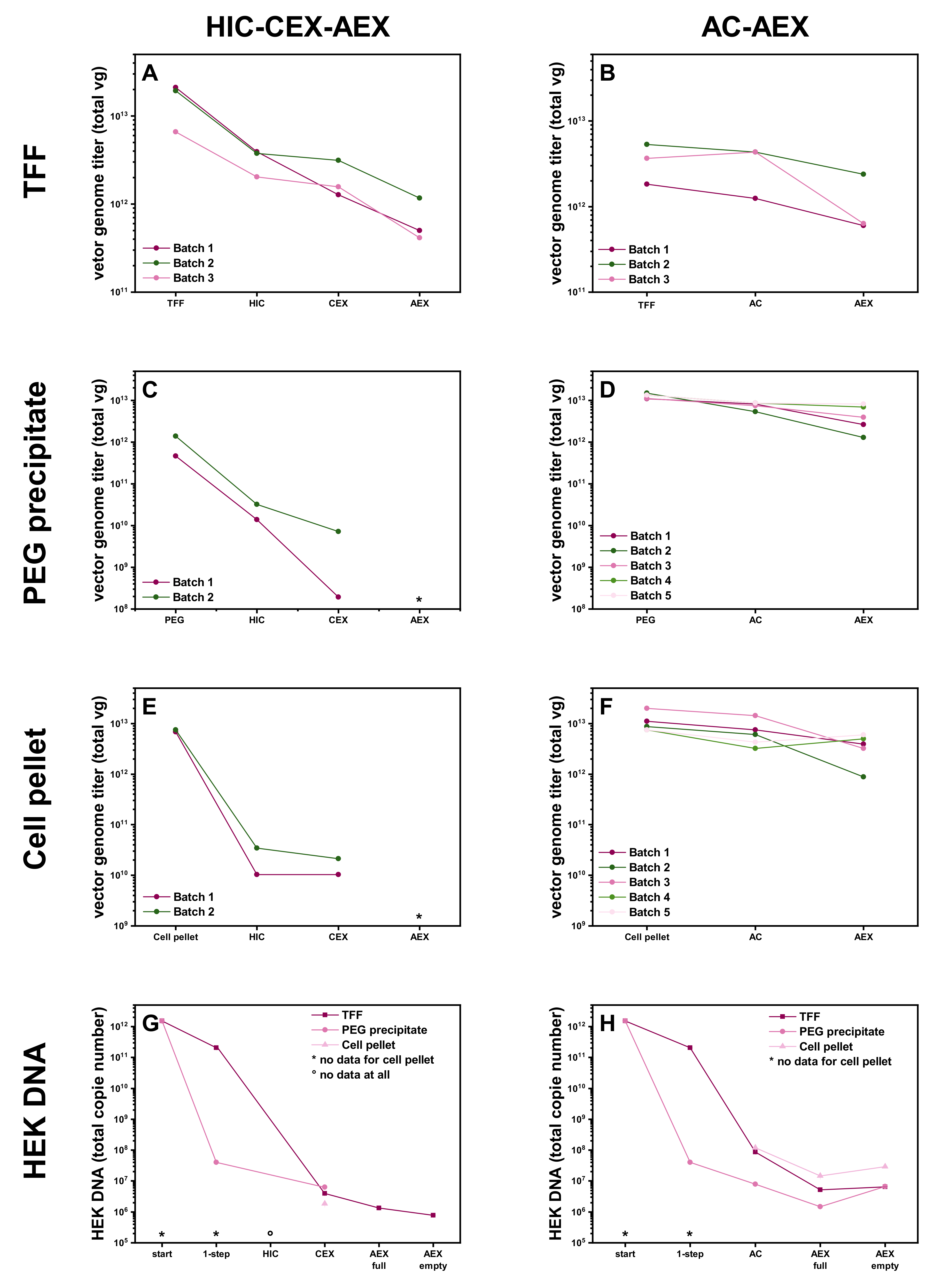
2.2. Tangential Flow Filtration—TFF
2.3. PEG Precipitation
2.4. Cell Pellet
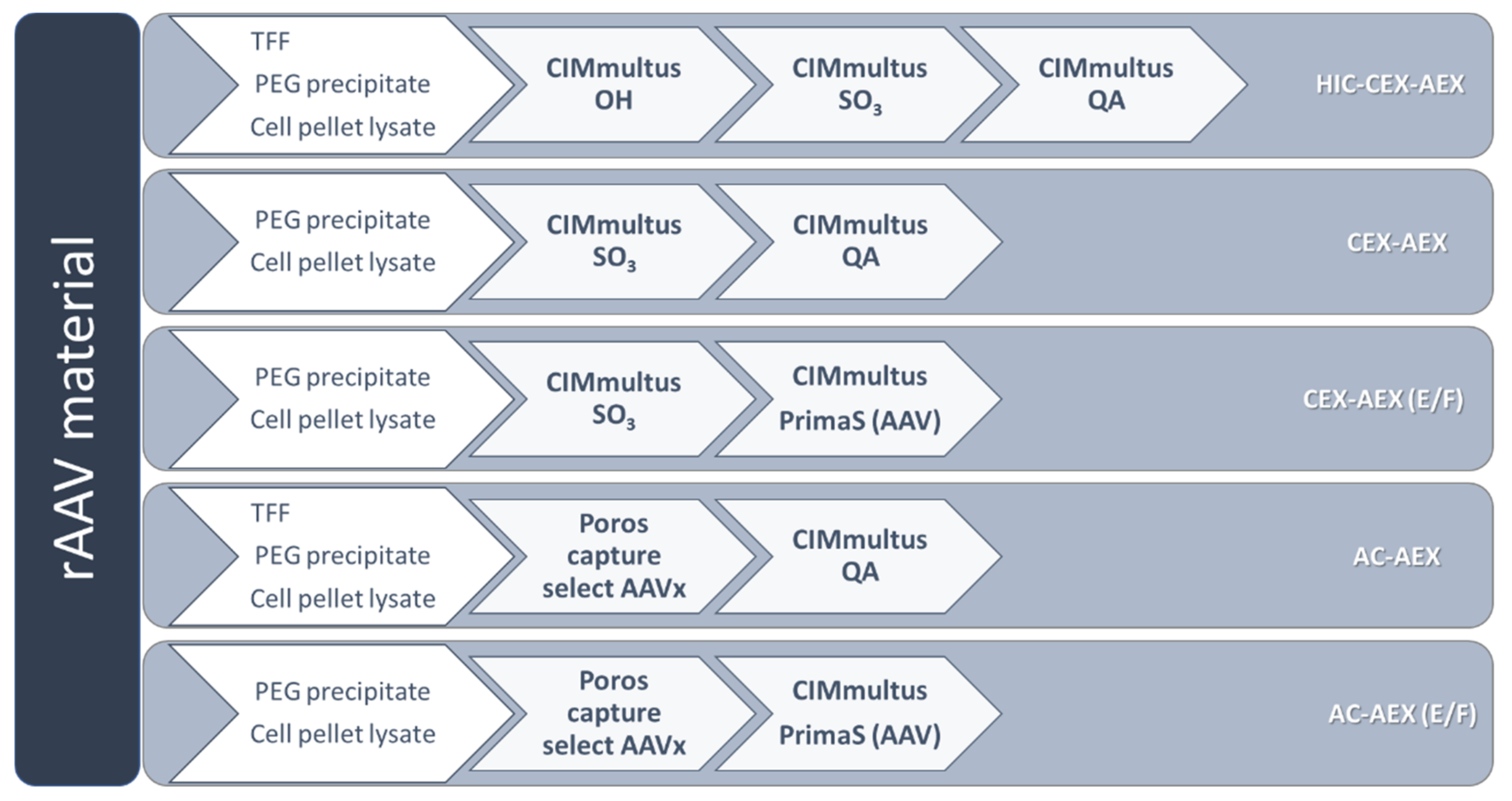
2.5. rAAV Vector Purification
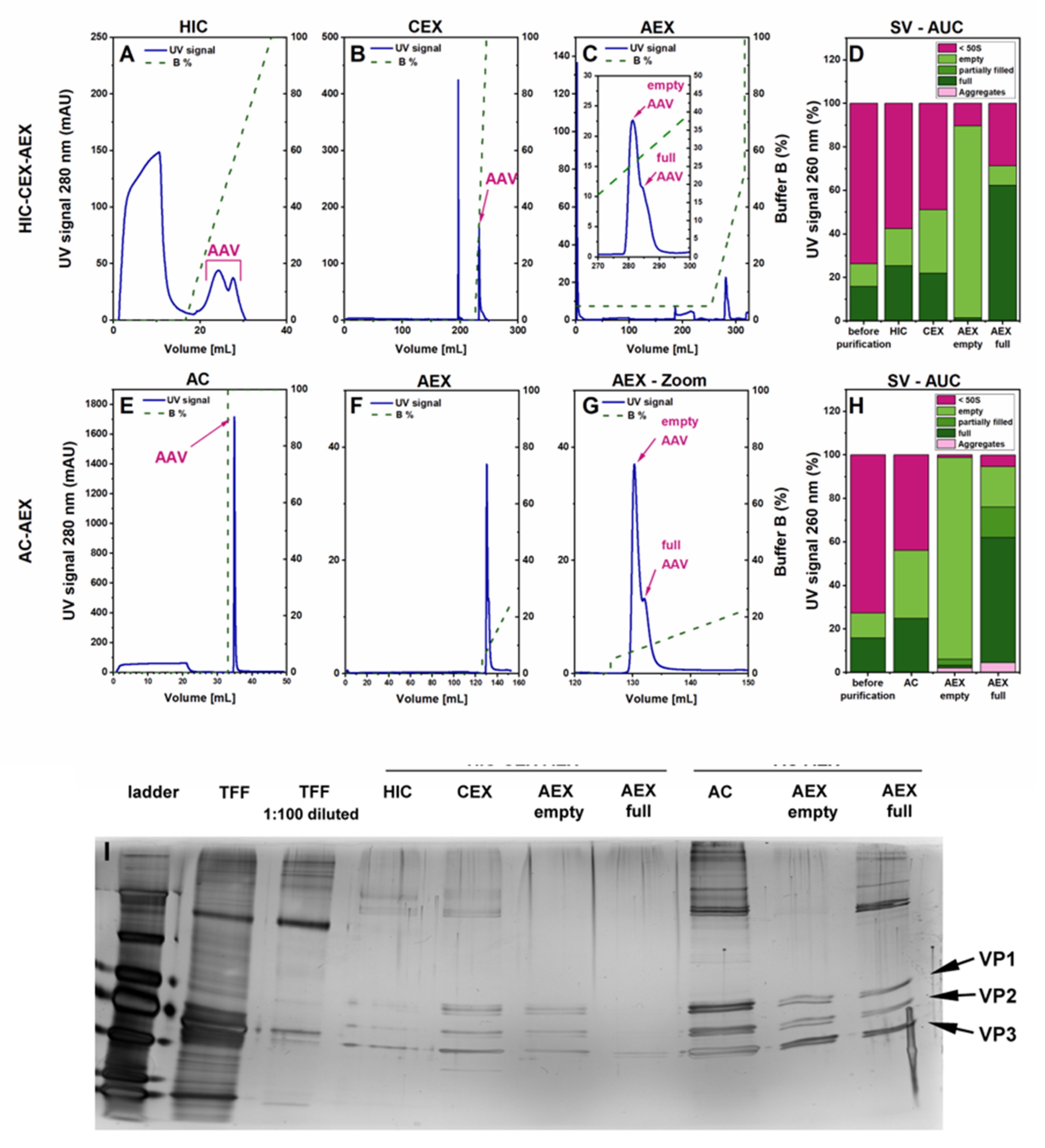
2.5.1. Hydrophobic Interaction Chromatography—HIC
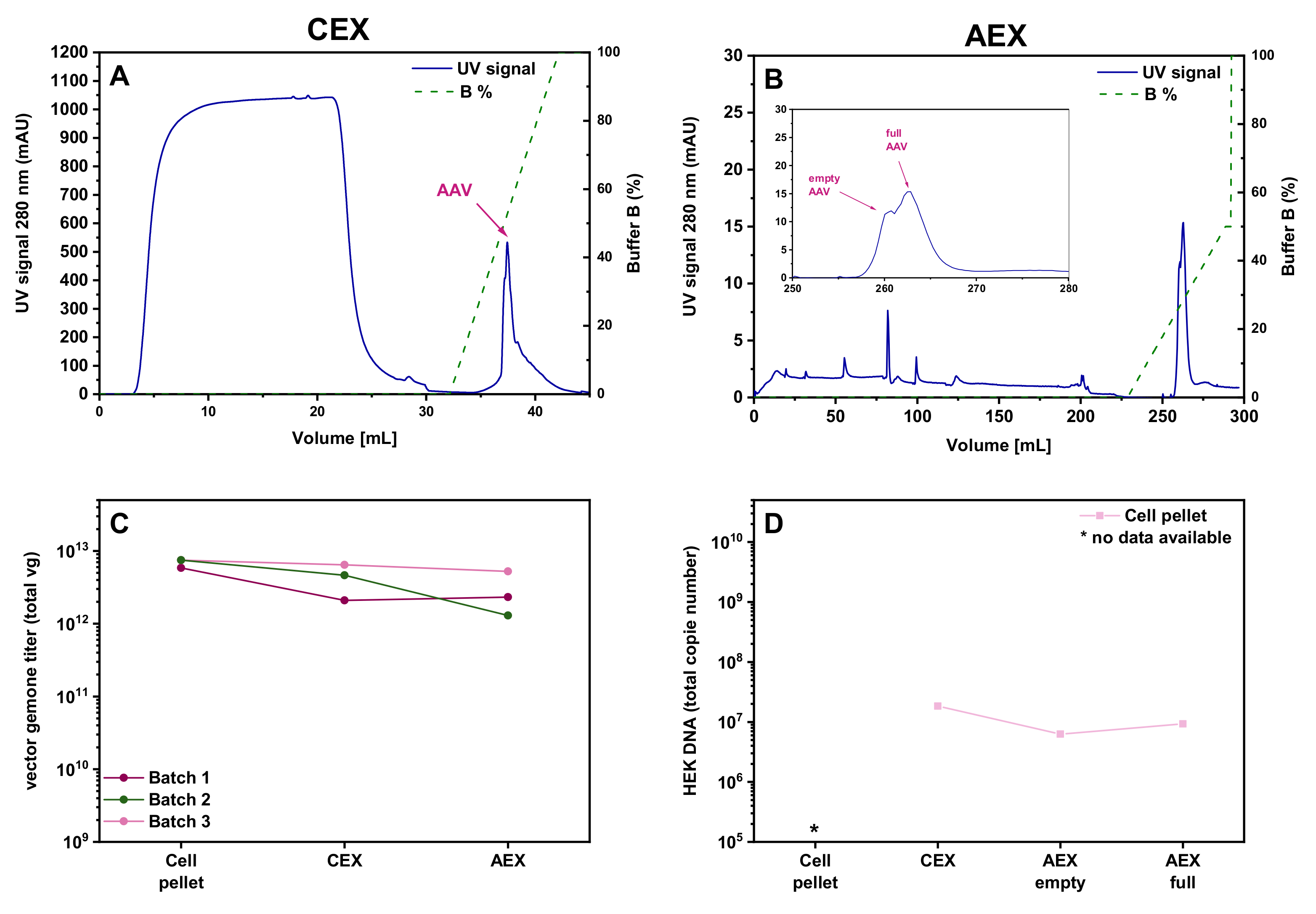
2.5.2. Cation Exchange Chromatography—CEX
2.5.3. Anion Exchange Chromatography—AEX—Salt Gradient
2.5.4. Anion Exchange Chromatography—AEX—pH Gradient
2.5.5. Affinity Chromatography—AC
2.6. Vector Genome Titer Determination by qPCR
2.7. Determination of HEK 293T DNA
2.8. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis—SDS-PAGE
2.9. Sedimentation Velocity Analytical Ultracentrifugation—SV-AUC
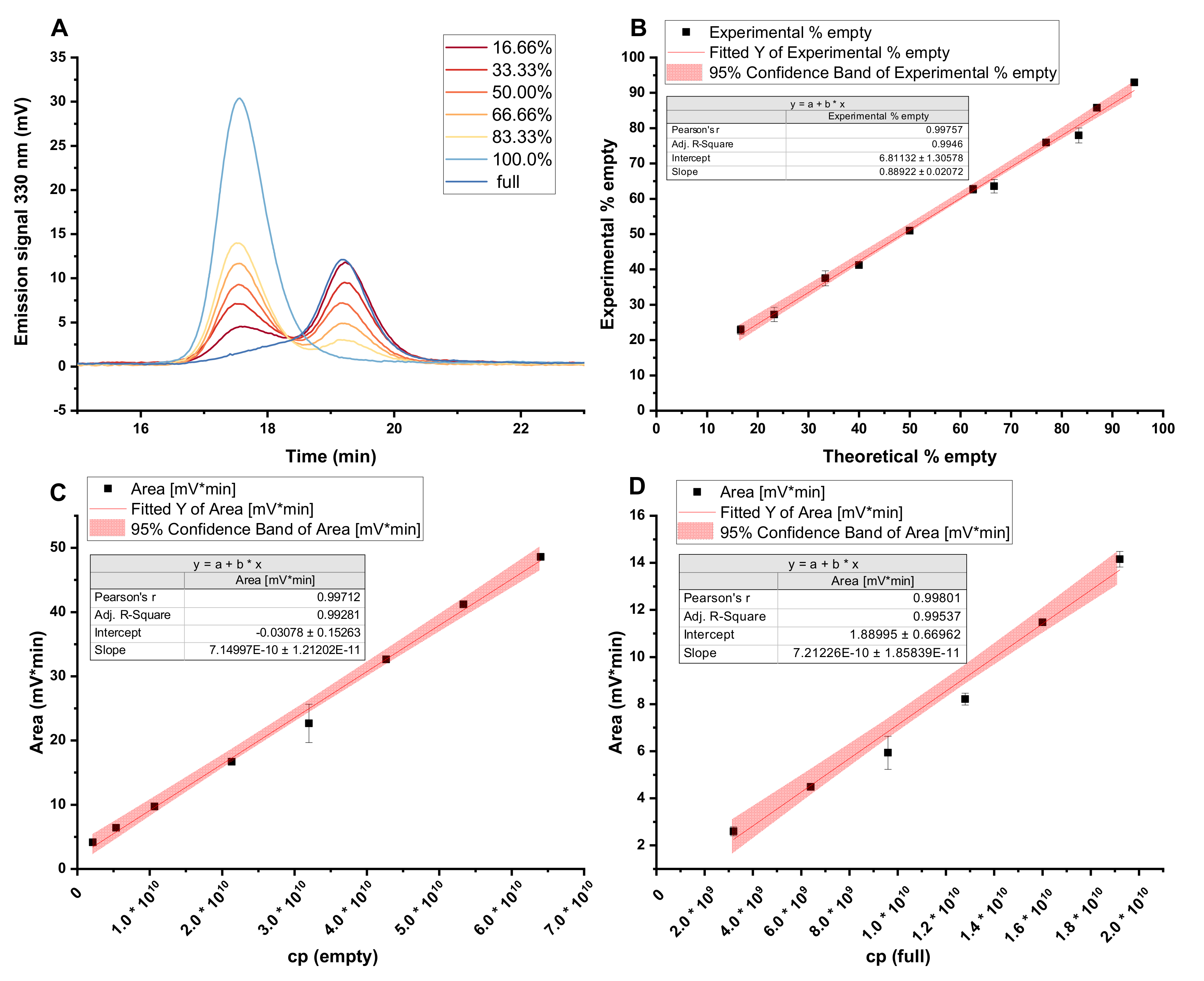
2.10. Analytical Anion Exchange Chromatography
3. Results
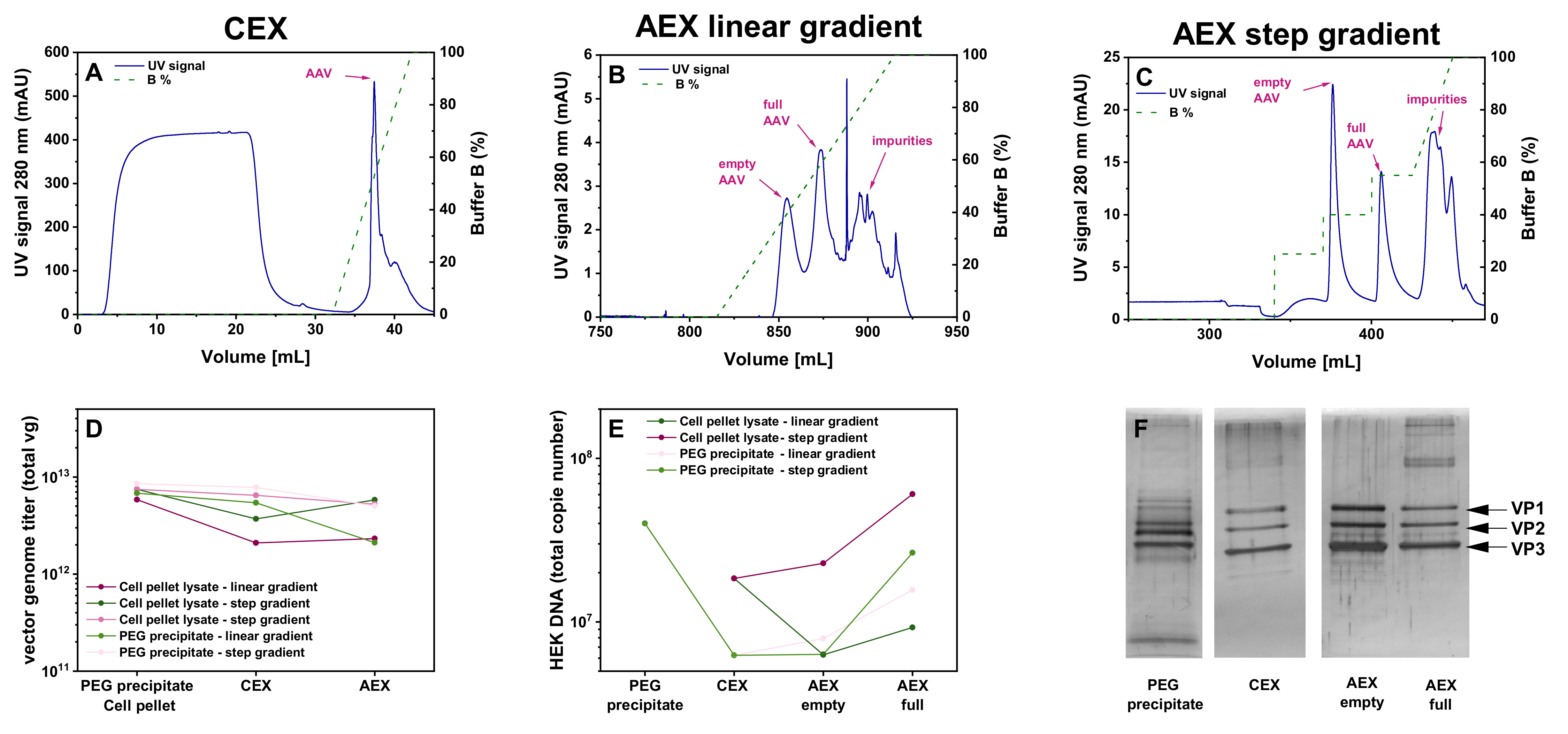
3.1. Evaluation of Two Different Liquid Chromatography-Based Purification Stragies for rAAV2/8 Vectors
3.2. Different Initial Steps for the Raw Material
3.3. Anion Exchange Chromatography Using a pH Gradient
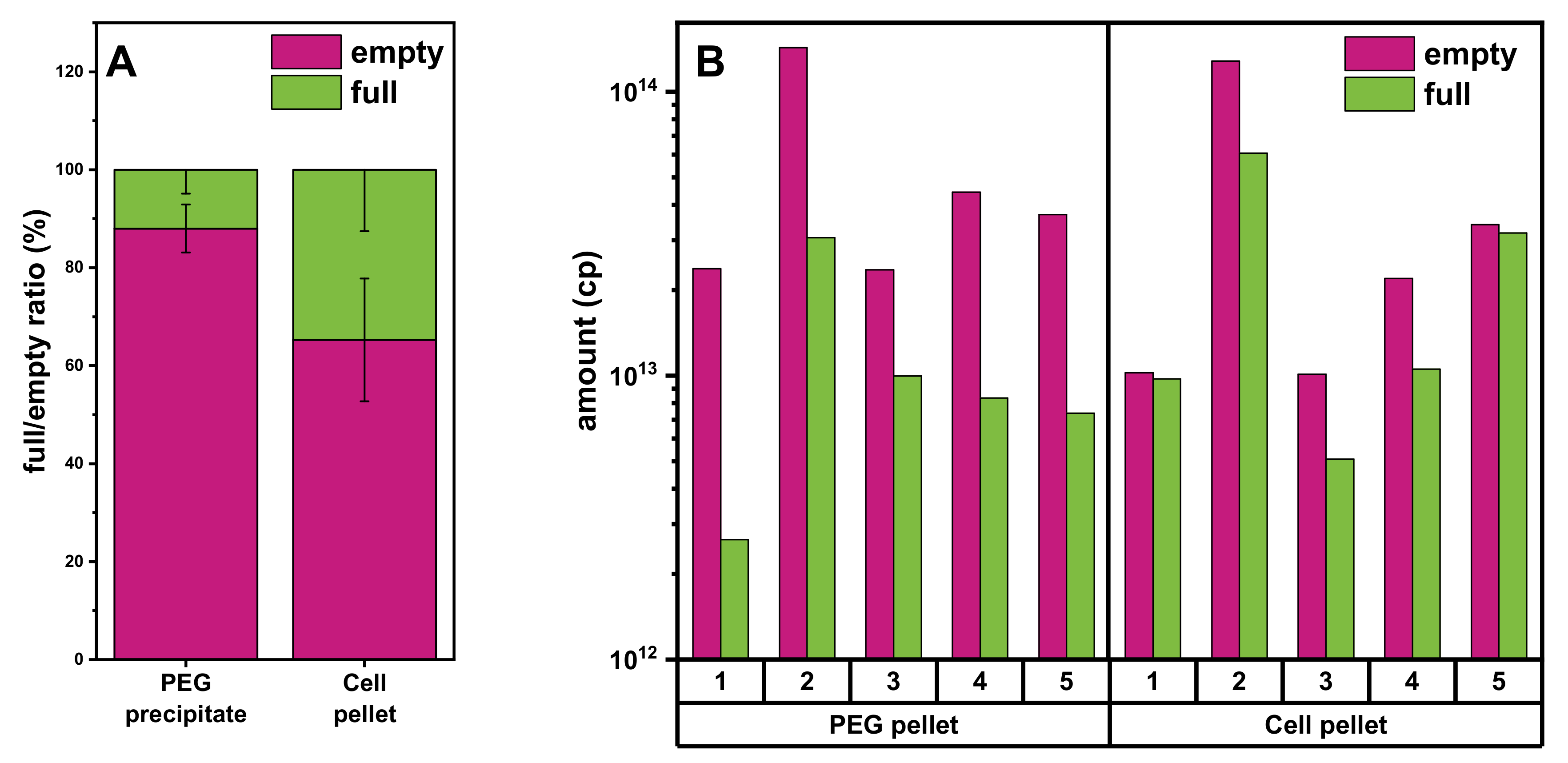
3.4. Comparison of Yields of Full rAAVs Harvested from Cell Pellet and Supernatant
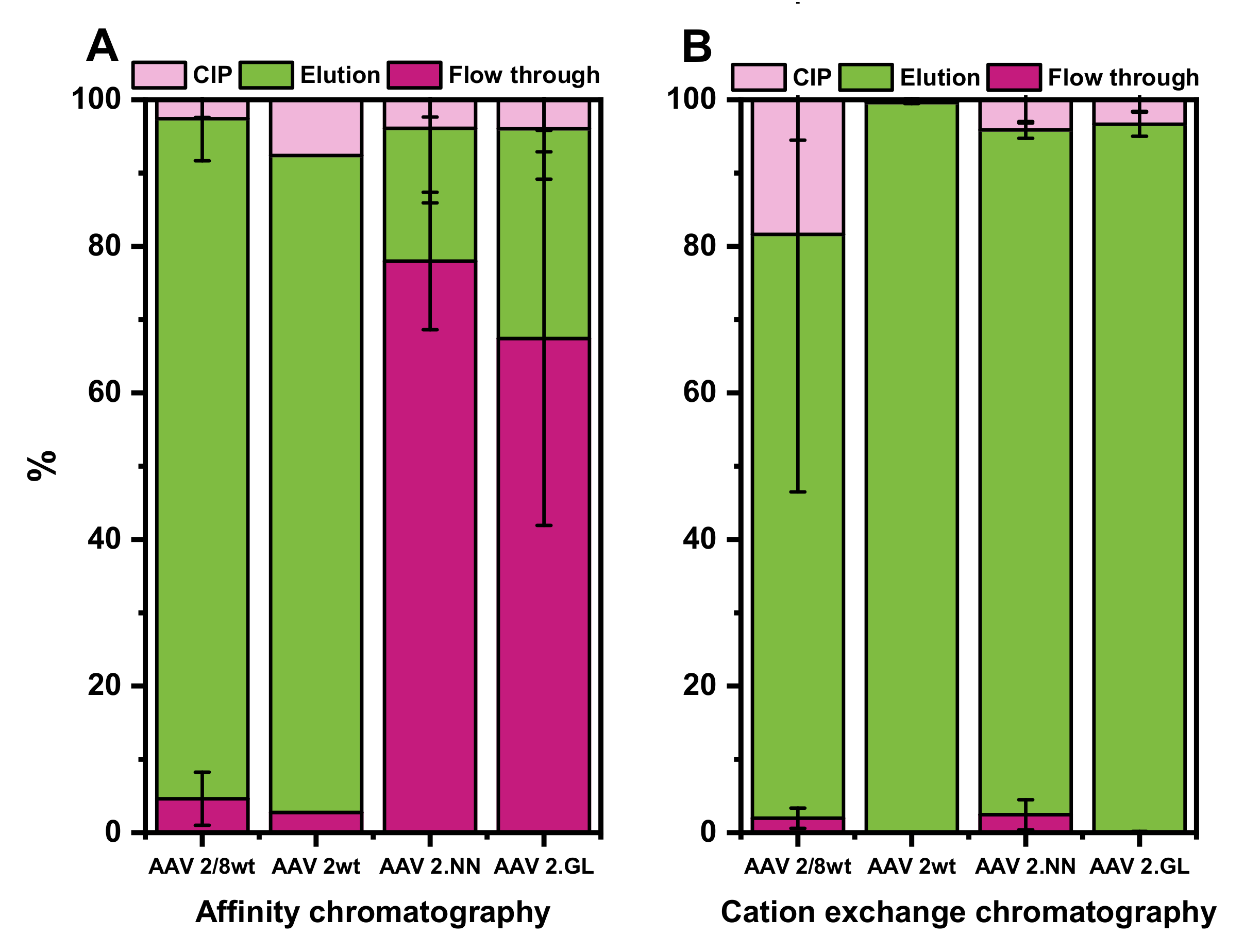
3.5. Evaluation of AC and CEX Purification Protocols for Four Different Serotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Ali, R.R.; Auricchio, A.; Smith, A.J. The future looks brighter after 25 years of retinal gene therapy. Hum. Gene Ther. 2017. [Google Scholar] [CrossRef] [Green Version]
- Boye, S.E.; Boye, S.L.; Lewin, A.S.; Hauswirth, W.W. A comprehensive review of retinal gene therapy. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Schön, C.; Biel, M.; Michalakis, S. Retinal gene delivery by adeno-associated virus (AAV) vectors: Strategies and applications. Eur. J. Pharm. Biopharm. 2015. [Google Scholar] [CrossRef]
- Trapani, I.; Puppo, A.; Auricchio, A. Vector platforms for gene therapy of inherited retinopathies. Prog. Retin. Eye Res. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samulski, R.J.; Srivastava, A.; Berns, K.I.; Muzyczka, N. Rescue of adeno-associated virus from recombinant plasmids: Gene correction within the terminal repeats of AAV. Cell 1983, 33, 135–143. [Google Scholar] [CrossRef]
- Adams, B.; Bak, H.; Tustian, A.D. Moving from the bench towards a large scale, industrial platform process for adeno-associated viral vector purification. Biotechnol. Bioeng. 2020. [Google Scholar] [CrossRef]
- Ayuso, E.; Mingozzi, F.; Montane, J.; Leon, X.; Anguela, X.M.; Haurigot, V.; Edmonson, S.A.; Africa, L.; Zhou, S.; High, K.A.; et al. High AAV vector purity results in serotype- and tissue-independent enhancement of transduction efficiency. Gene Ther. 2010, 17, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.; Bahr-Davidson, J.; Prado, J.; Tai, A.; Cataniag, F.; McDonnell, J.; Zhou, J.; Hauck, B.; Luna, J.; Sommer, J.M.; et al. Separation of adeno-associated virus type 2 empty particles from genome containing vectors by anion-exchange column chromatography. J. Virol. Methods 2007, 140, 183–192. [Google Scholar] [CrossRef]
- Burnham, B.; Nass, S.; Kong, E.; Mattingly, M.; Woodcock, D.; Song, A.; Wadsworth, S.; Cheng, S.H.; Scaria, A.; O’Riordan, C.R. Analytical Ultracentrifugation as an Approach to Characterize Recombinant Adeno-Associated Viral Vectors. Hum. Gene Ther. Methods 2015, 26, 228–242. [Google Scholar] [CrossRef]
- Nass, S.A.; Mattingly, M.A.; Woodcock, D.A.; Burnham, B.L.; Ardinger, J.A.; Osmond, S.E.; Frederick, A.M.; Scaria, A.; Cheng, S.H.; O’Riordan, C.R. Universal Method for the Purification of Recombinant AAV Vectors of Differing Serotypes. Mol. Ther. Methods Clin. Dev. 2018, 9, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Davidoff, A.M.; Ng, C.Y.; Sleep, S.; Gray, J.; Azam, S.; Zhao, Y.; McIntosh, J.H.; Karimipoor, M.; Nathwani, A.C. Purification of recombinant adeno-associated virus type 8 vectors by ion exchange chromatography generates clinical grade vector stock. J. Virol. Methods 2004, 121, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Qu, G.; Burnham, M.S.; Huang, J.; Chirmule, N.; Joshi, B.; Yu, Q.C.; Marsh, J.A.; Conceicao, C.M.; Wilson, J.M. Purification of recombinant adeno-associated virus vectors by column chromatography and its performance in vivo. Hum. Gene Ther. 2000, 11, 2079–2091. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Wang, M.; Wu, Y.; Xu, R. Scalable downstream strategies for purification of recombinant adeno- associated virus vectors in light of the properties. Curr. Pharm. Biotechnol. 2015, 16, 684–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaludov, N.; Handelman, B.; Chiorini, J.A. Scalable purification of adeno-associated virus type 2, 4, or 5 using ion-exchange chromatography. Hum. Gene Ther. 2002, 13, 1235–1243. [Google Scholar] [CrossRef]
- Wright, J. Product-Related Impurities in Clinical-Grade Recombinant AAV Vectors: Characterization and Risk Assessment. Biomedicines 2014, 2, 80–97. [Google Scholar] [CrossRef]
- Flotte, T.R.; Brantly, M.L.; Spencer, L.T.; Byrne, B.J.; Spencer, C.T.; Baker, D.J.; Humphries, M. Phase I trial of intramuscular injection of a recombinant adeno-associated virus alpha 1-antitrypsin (rAAV2-CB-hAAT) gene vector to AAT-deficient adults. Hum. Gene Ther. 2004, 15, 93–128. [Google Scholar] [CrossRef]
- Flotte, T.R.; Zeitlin, P.L.; Reynolds, T.C.; Heald, A.E.; Pedersen, P.; Beck, S.; Conrad, C.K.; Brass-Ernst, L.; Humphries, M.; Sullivan, K. Phase I trial of intranasal and endobronchial administration of a recombinant adeno-associated virus serotype 2 (rAAV2)-CFTR vector in adult cystic fibrosis patients: A two-part clinical study. Hum. Gene Ther. 2003, 14, 1079–1088. [Google Scholar] [CrossRef]
- Bowles, D.E.; McPhee, S.W.J.; Li, C.; Gray, S.J.; Samulski, J.J.; Camp, A.S.; Li, J.; Wang, B.; Monahan, P.E.; Rabinowitz, J.E.; et al. Phase 1 Gene Therapy for Duchenne Muscular Dystrophy Using a Translational Optimized AAV Vector. Mol. Ther. 2012, 20, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.F. AAV empty capsids: For better or for worse? Mol. Ther. 2014, 22, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becirovic, E.; Bohm, S.; Nguyen, O.N.; Riedmayr, L.M.; Hammelmann, V.; Schon, C.; Butz, E.S.; Wahl-Schott, C.; Biel, M.; Michalakis, S. AAV Vectors for FRET-Based Analysis of Protein-Protein Interactions in Photoreceptor Outer Segments. Front. Neurosci. 2016, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Schon, C.; Becirovic, E.; Biel, M.; Michalakis, S. Design and Development of AAV-based Gene Supplementation Therapies for Achromatopsia and Retinitis Pigmentosa. Methods Mol. Biol. 2018, 1715, 33–46. [Google Scholar] [CrossRef]
- Pavlou, M.; Schön, C.; Occelli, L.M.; Rossi, A.; Meumann, N.; Boyd, R.F.; Bartoe, J.T.; Siedlecki, J.; Gerhardt, M.J.; Babutzka, S.; et al. Novel AAV capsids for intravitreal gene therapy of photoreceptor disorders. EMBO Mol. Med. 2021, e13392. [Google Scholar] [CrossRef]
- Leskovec, M.; Primec, S.; Gagnon, P.; Štrancar, A. An Industrial Purification Platform for AAV. 2018. Available online: https://www.biaseparations.com/en/library/posters/988/an-industrialpurification-platform-for-aav (accessed on 24 March 2021).
- Lock, M.; Alvira, M.R.; Wilson, J.M. Analysis of Particle Content of Recombinant Adeno-Associated Virus Serotype 8 Vectors by Ion-Exchange Chromatography. Hum. Gene Ther. Methods 2012, 23, 56–64. [Google Scholar] [CrossRef]
- D’Costa, S.; Blouin, V.; Broucque, F.; Penaud-Budloo, M.; Francois, A.; Perez, I.C.; Le Bec, C.; Moullier, P.; Snyder, R.O.; Ayuso, E. Practical utilization of recombinant AAV vector reference standards: Focus on vector genomes titration by free ITR qPCR. Mol. Ther. Methods Clin. Dev. 2016, 5, 16019. [Google Scholar] [CrossRef]
- Wang, Y.; Cooper, R.; Kiladjian, A.; Bergelson, S.; Feschenko, M. A Digestion-free Method for Quantification of Residual Host Cell DNA in rAAV Gene Therapy Products. Mol. Ther. Methods Clin. Dev. 2019, 13, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlbrenner, E.; Henckaerts, E.; Rapti, K.; Gordon, R.E.; Linden, R.M.; Hajjar, R.J.; Weber, T. Quantification of AAV particle titers by infrared fluorescence scanning of coomassie-stained sodium dodecyl sulfate-polyacrylamide gels. Hum. Gene Ther. Methods 2012, 23, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Koza, S.M.; Chen, W. Anion-Exchange Chromatography for Determining Empty and Full Capsid; Waters Corporation: Milford, MA, USA, 2019; Available online: https://www.waters.com/content/dam/waters/en/app-notes/2020/720006825/720006825-zh.pdf (accessed on 26 November 2020).
- Terova, O.; Soltys, S.; Hermans, P.; De Rooij, J.; Detmers, F. Overcoming Downstream Purification Challenges for Viral Vector Manufacturing: Enabling Advancement of Gene Therapies in the Clinic. Cell Gene Ther. Insights 2018, 4, 101–111. [Google Scholar] [CrossRef]
- Mietzsch, M.; Jose, A.; Chipman, P.; Bhattacharya, N.; Daneshparvar, N.; McKenna, R.; Agbandje-McKenna, M. Completion of the AAV Structural Atlas: Serotype Capsid Structures Reveals Clade-Specific Features. Viruses 2021, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L., 3rd; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Hebben, M. Downstream bioprocessing of AAV vectors: Industrial challenges & regulatory requirements. Cell Gene Ther. Insights 2018, 4, 131–146. [Google Scholar] [CrossRef]
- Lins-Austin, B.; Patel, S.; Mietzsch, M.; Brooke, D.; Bennett, A.; Venkatakrishnan, B.; Van Vliet, K.; Smith, A.N.; Long, J.R.; McKenna, R. Adeno-associated virus (AAV) Capsid stability and liposome remodeling during Endo/Lysosomal pH trafficking. Viruses 2020, 12, 668. [Google Scholar] [CrossRef] [PubMed]
- Urabe, M.; Xin, K.Q.; Obara, Y.; Nakakura, T.; Mizukami, H.; Kume, A.; Okuda, K.; Ozawa, K. Removal of empty capsids from type 1 adeno-associated virus vector stocks by anion-exchange chromatography potentiates transgene expression. Mol. Ther. 2006, 13, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Schnodt, M.; Buning, H. Improving the Quality of Adeno-Associated Viral Vector Preparations: The Challenge of Product-Related Impurities. Hum. Gene Ther. Methods 2017, 28, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and dynamics of adeno-associated virus serotype 1 VP1-unique N-terminal domain and its role in capsid trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Mulagapati, S.H.R.; Chen, Z.; Du, J.; Zhao, X.; Xi, G.; Chen, L.; Linke, T.; Gao, C.; Schmelzer, A.E.; et al. Developing an Anion Exchange Chromatography Assay for Determining Empty and Full Capsid Contents in AAV6.2. Mol. Ther. Methods Clin. Dev. 2019, 15, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Tomono, T.; Hirai, Y.; Okada, H.; Miyagawa, Y.; Adachi, K.; Sakamoto, S.; Kawano, Y.; Chono, H.; Mineno, J.; Ishii, A.; et al. Highly Efficient Ultracentrifugation-free Chromatographic Purification of Recombinant AAV Serotype 9. Mol. Ther. Methods Clin. Dev. 2018, 11, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.R.; Barnier, A.; Moço, P.D.; Schrag, J.; Chahal, P.S.; Kamen, A. Development of a Scalable and Robust Anion-Exchange Chromatographic Method for Enriched Recombinant Adeno-Associated Virus Preparations in Genome Containing Vector Capsids of Serotypes-5, 6, 8, and 9. Mol. Ther. Methods Clin. Dev. 2021, 21, 341–356. [Google Scholar] [CrossRef]
- Dickerson, R.; Argento, C.; Pieracci, J.; Bakhshayeshi, M. Separating Empty and Full Recombinant Adeno-Associated Virus Particles Using Isocratic Anion Exchange Chromatography. Biotechnol. J. 2021, 16, 2000015. [Google Scholar] [CrossRef]
- Mary, B.; Maurya, S.; Arumugam, S.; Kumar, V.; Jayandharan, G.R. Post-translational modifications in capsid proteins of recombinant adeno-associated virus (AAV) 1-rh10 serotypes. FEBS J. 2019, 286, 4964–4981. [Google Scholar] [CrossRef]
- Zhong, L.; Li, B.; Jayandharan, G.; Mah, C.S.; Govindasamy, L.; Agbandje-McKenna, M.; Herzog, R.W.; Weigel-Van Aken, K.A.; Hobbs, J.A.; Zolotukhin, S.; et al. Tyrosine-phosphorylation of AAV2 vectors and its consequences on viral intracellular trafficking and transgene expression. Virology 2008, 381, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Giles, A.R.; Sims, J.J.; Turner, K.B.; Govindasamy, L.; Alvira, M.R.; Lock, M.; Wilson, J.M. Deamidation of Amino Acids on the Surface of Adeno-Associated Virus Capsids Leads to Charge Heterogeneity and Altered Vector Function. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 2848–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burova, E.; Ioffe, E. Chromatographic purification of recombinant adenoviral and adeno-associated viral vectors: Methods and implications. Gene Ther. 2005, 12 (Suppl. 1), S5–S17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosson, S.M.; Dib, P.; Smith, J.K.; Zolotukhin, S. Helper-free Production of Laboratory Grade AAV and Purification by Iodixanol Density Gradient Centrifugation. Mol. Ther. Methods Clin. Dev. 2018, 10, 1–7. [Google Scholar] [CrossRef]
- Merten, O.W.; Geny-Fiamma, C.; Douar, A.M. Current issues in adeno-associated viral vector production. Gene Ther. 2005, 12 (Suppl. 1), S51–S61. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Strategy | Purification Step | TFF (%) | SD (%) | PEG Precipitate (%) | SD (%) | Cell Pellet Lysate (%) | SD (%) |
|---|---|---|---|---|---|---|---|
| HIC-CEX-AEX | HIC | 23 | 6 | 3 | - | 0.3 | - |
| CEX | 15 | 7 | 0.3 | - | 0.2 | - | |
| AEX | 5 | 2 | ND | ND | ND | ND | |
| CEX-AEX | CEX | ND | ND | ND | ND | 65 | 13 |
| AEX | ND | ND | ND | ND | 59 | 24 | |
| AC-AEX | AC | 89 | 21 | 69 | 24 | 59 | 12 |
| AEX | 32 | 11 | 44 | 21 | 37 | 23 |
| Purification Strategy | Number of Preparations | Residual Amount of Empty Capsids in Full rAAV Fractions (%) | |
|---|---|---|---|
| Entire Peak | Peak Maximum | ||
| CEX-AEX(E/F) linear | 1 | 27 | 21 |
| 2 | 31 | 23 | |
| CEX-AEX(E/F) steps | 1 | 17 | 8 |
| 2 | 17 | 12 | |
| 3 | 20 | 9 | |
| AC-AEX(E/F) linear | 1 | 23 | 15 |
| 2 | 40 | 28 | |
| AC-AEX(E/F) steps | 1 | 20 | 13 |
| Purification Strategy | Number of Preparations | Purification Step | Yield (%) |
|---|---|---|---|
| CEX-AEX(E/F) linear | 1: Cell pellet | CEX | 64 |
| AEX | 70 | ||
| 2: PEG pellet | CEX | 80 | |
| AEX | 31 | ||
| CEX-AEX(E/F) steps | 1: Cell pellet | CEX | 86 |
| AEX | 70 | ||
| 2: Cell pellet | CEX | 55 | |
| AEX | 78 | ||
| 3: PEG pellet | CEX | 92 | |
| AEX | 63 |
| Purification Strategy | Yield for AAV2/8 | Working Time (Hours) | Scalability | GMP Readiness | Full/Empty Separation | Remaining Impurities |
|---|---|---|---|---|---|---|
| ultracentrifugation | good | 8–10 | poor | moderate | after 2 runs good | iodixanol/cesium chloride |
| AC | good to very good | 2–3 | good | good | poor | DNA |
| CEX | good to very good | 0.5–1 | good | good | poor | proteins, DNA |
| AEX | good | 2–3 | good | good | good | small amounts of DNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rieser, R.; Koch, J.; Faccioli, G.; Richter, K.; Menzen, T.; Biel, M.; Winter, G.; Michalakis, S. Comparison of Different Liquid Chromatography-Based Purification Strategies for Adeno-Associated Virus Vectors. Pharmaceutics 2021, 13, 748. https://doi.org/10.3390/pharmaceutics13050748
Rieser R, Koch J, Faccioli G, Richter K, Menzen T, Biel M, Winter G, Michalakis S. Comparison of Different Liquid Chromatography-Based Purification Strategies for Adeno-Associated Virus Vectors. Pharmaceutics. 2021; 13(5):748. https://doi.org/10.3390/pharmaceutics13050748
Chicago/Turabian StyleRieser, Ruth, Johanna Koch, Greta Faccioli, Klaus Richter, Tim Menzen, Martin Biel, Gerhard Winter, and Stylianos Michalakis. 2021. "Comparison of Different Liquid Chromatography-Based Purification Strategies for Adeno-Associated Virus Vectors" Pharmaceutics 13, no. 5: 748. https://doi.org/10.3390/pharmaceutics13050748