
Chondroitin Sulfate in USA Dietary Supplements in Comparison to Pharma Grade Products: Analytical Fingerprint and Potential Anti-Inflammatory Effect on Human Osteoartritic Chondrocytes and Synoviocytes

 , and
, and
Abstract
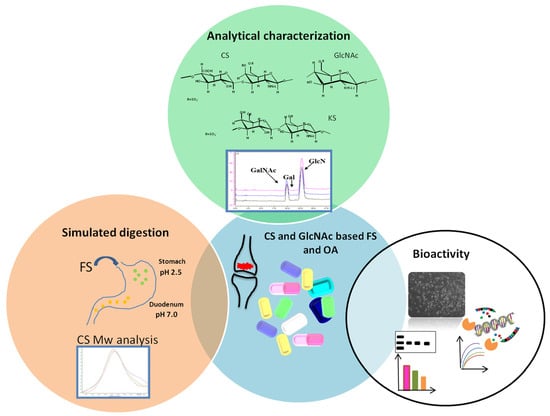
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Solid Fraction Determination
2.2.2. HPAE-PAD Analyses of the Monosaccharide Composition
2.2.3. HPCE Analyses of CS Disaccharide Composition
2.2.4. SEC-TDA Analyses of CS Average Molecular Weight
2.2.5. In Vitro Simulated Gastric and Intestinal Digestion of Ph and US FS Samples
2.2.6. HPCE Analyses of CS after In Vitro Simulated Gastric and Intestinal Digestion
2.2.7. Biological Activity Assays
2.2.8. Isolation and Culture of Cartilage Chondrocytes and Knee Joint Synoviocytes
2.2.9. Cell Viability Evaluation through MTT-Test
2.2.10. Cell Growth and Proliferation Using Time Lapse Video Microscopy (TLVM)
2.2.11. Gene Expression Analyses of COMP-2 and HAS-1 by qRT-PCR
2.2.12. COMP-2, NF-kB and PTX-3 Protein Expression Using Western Blotting Analyses
2.2.13. IL-6 and IL-8 Cytokines Quantification Using ELISA Assay
3. Results
3.1. Solid Fraction Determination
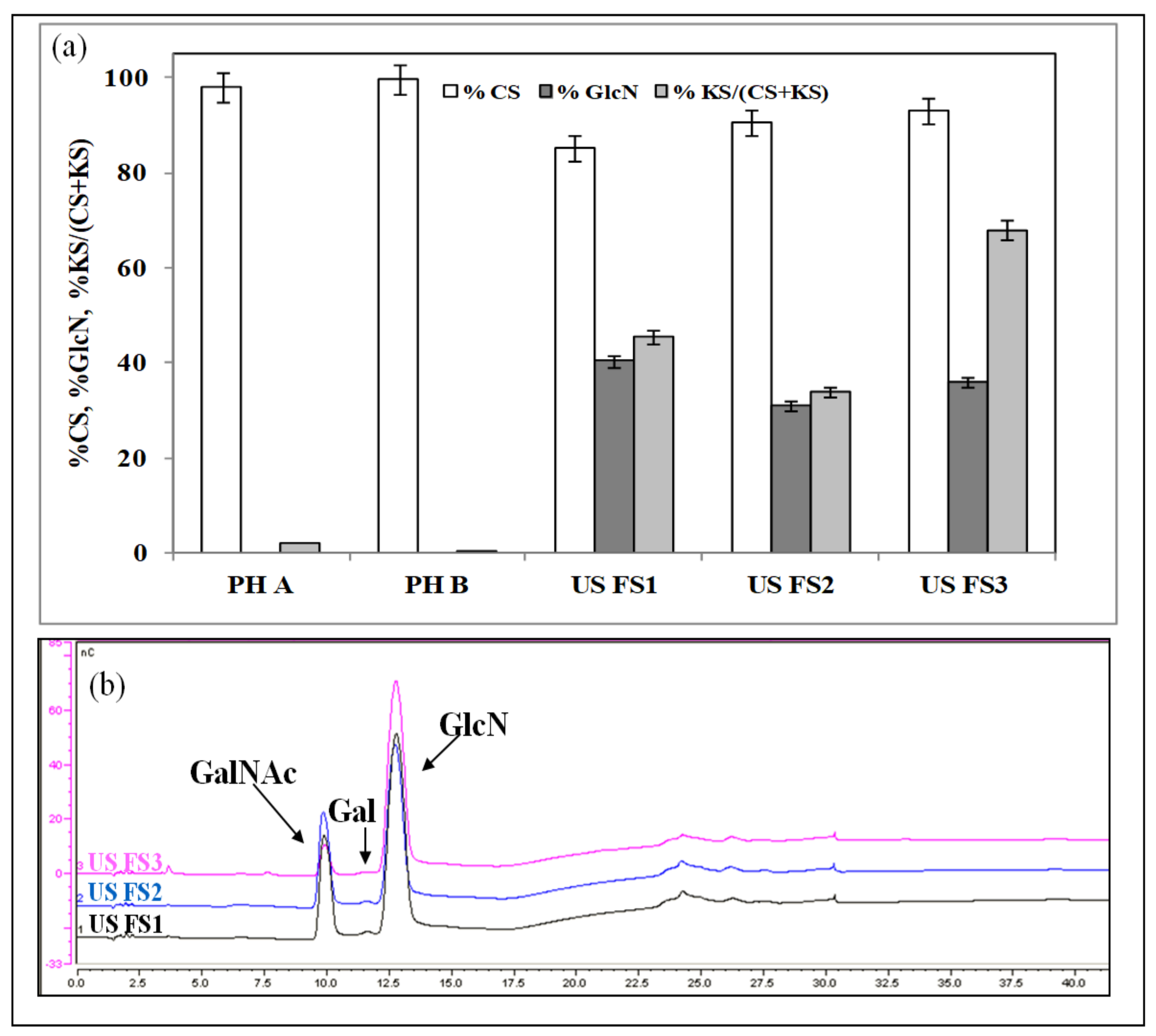
3.2. Monosaccharide Composition by HPAE-PAD, CS Disaccharide Composition by HPCE, CS Average Molecular Weight by SEC-TDA
3.3. CS Analyses after In Vitro Simulated Gastric and Intestinal Digestion
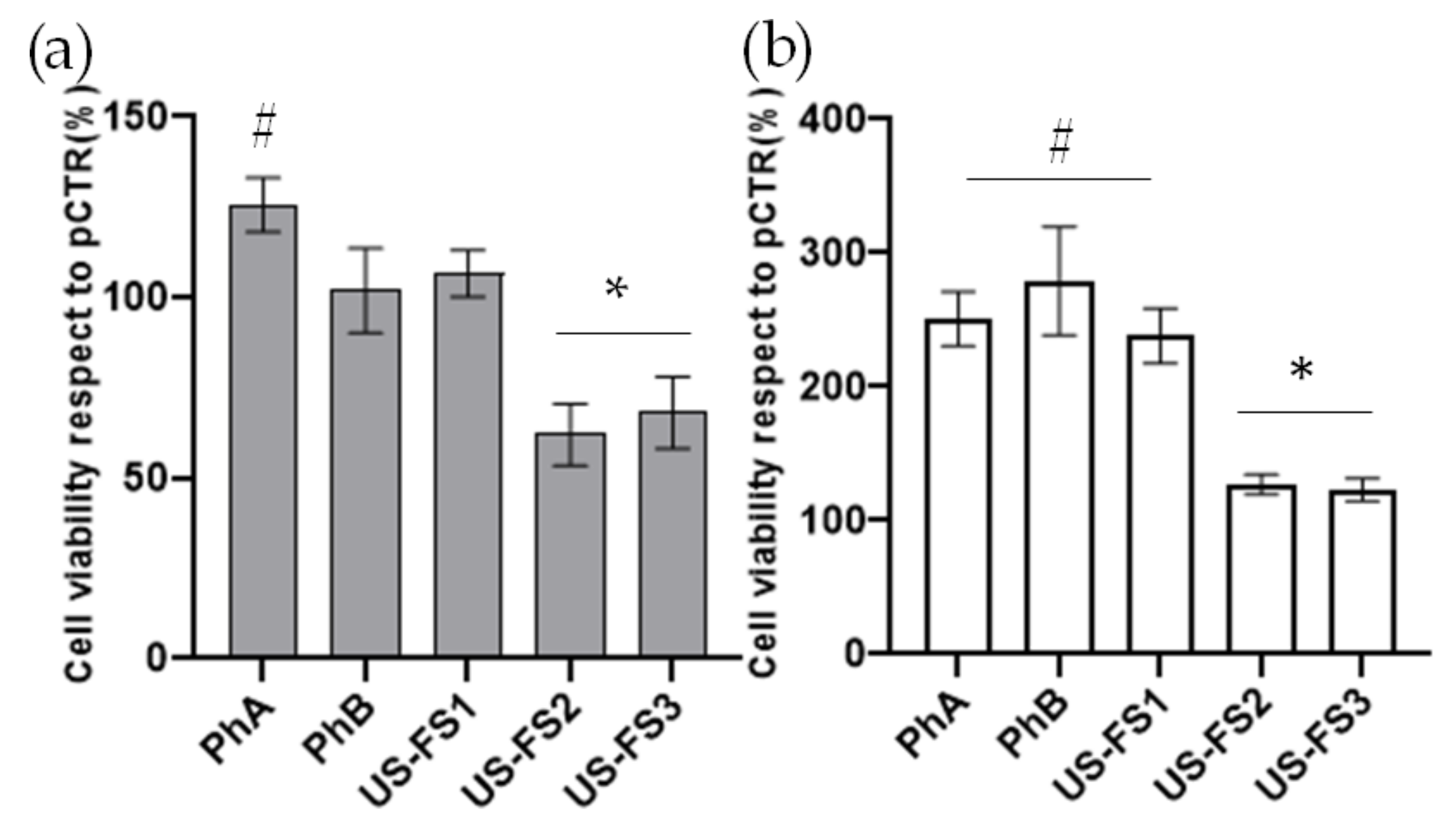
3.4. Cell Viability on Chondrocytes Using MTT Assay
3.5. Cell Growth and Proliferation Evaluation Using Time Lapse Video Microscopy
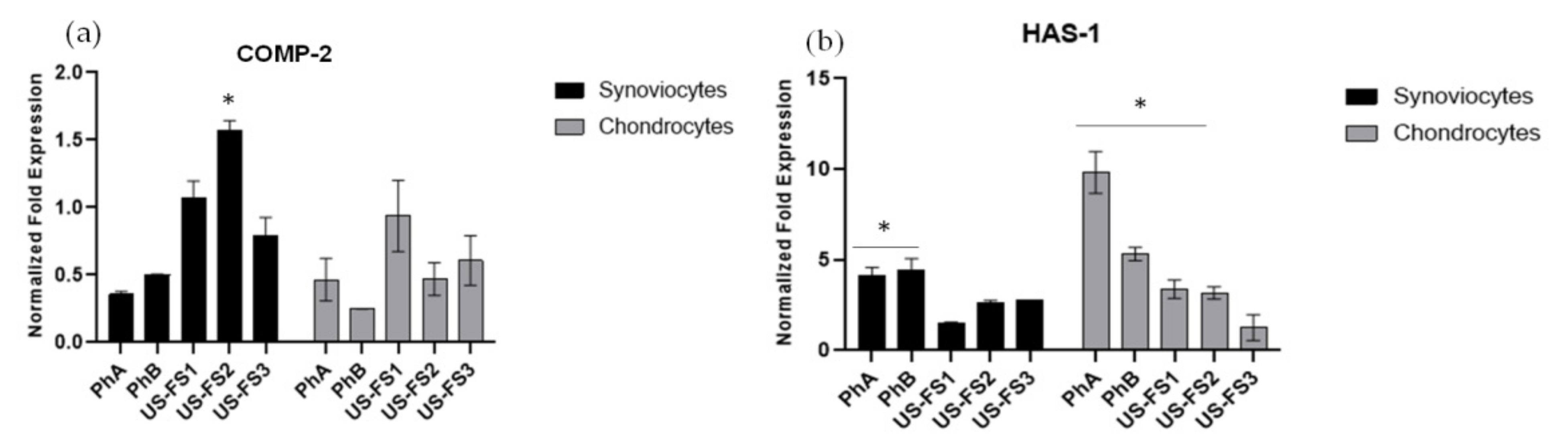
3.6. COMP-2 and HAS-1 Gene Expression Analyses by qRT-PCR
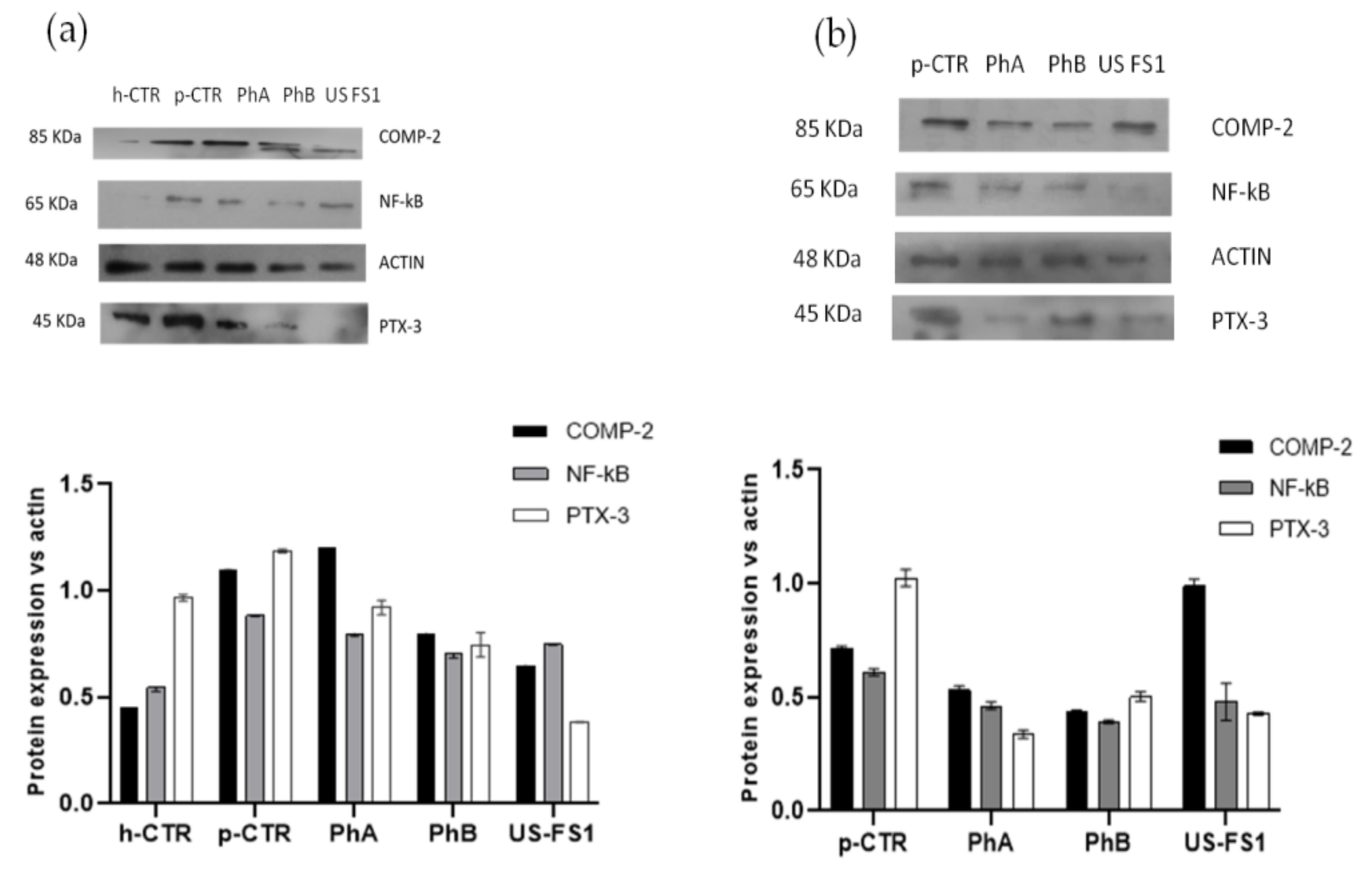
3.7. NF-kB, COMP-2 and PTX-3 Protein Expression in Human OA Chondrocytes and Synoviocytes: WB Analyses
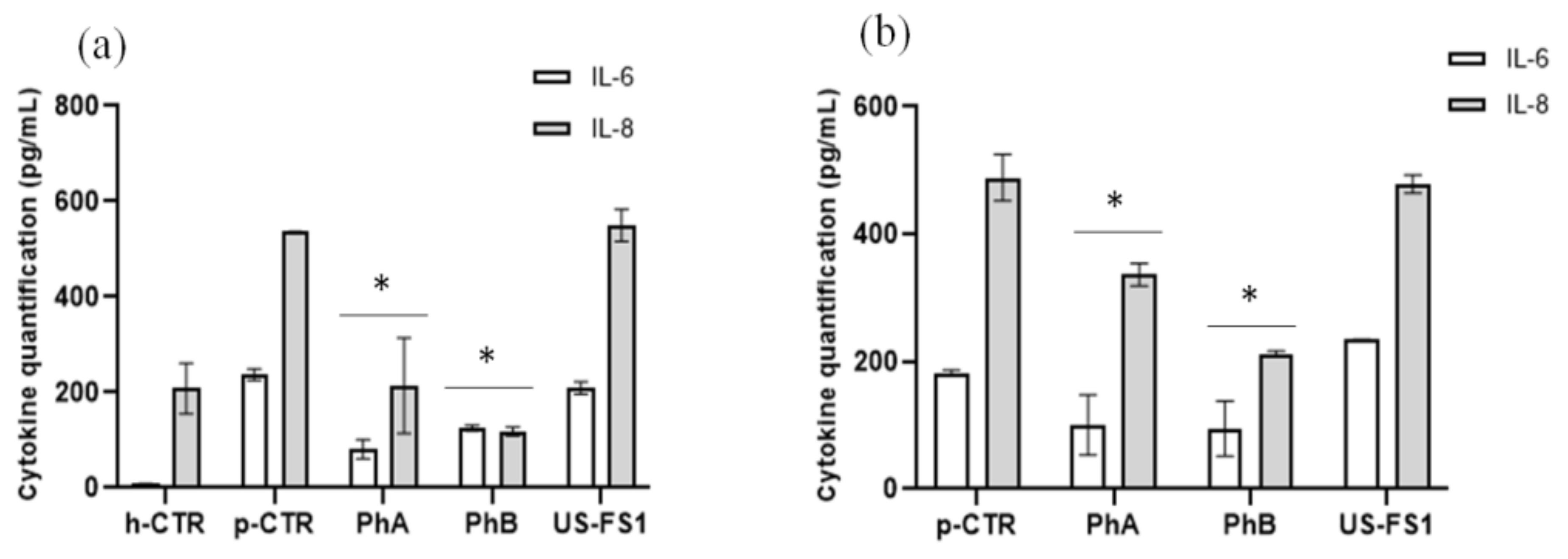
3.8. IL-6 and IL-8 Quantification on Chondrocytes and Synoviocytes Using ELISA Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-kappaB signaling pathways in osteoarthritic cartilage destruction. Cells 2019, 17, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restaino, O.F.; Finamore, R.; Stellavato, A.; Diana, P.; Bedini, E.; Trifuoggi, M.; De Rosa, M.; Schiraldi, C. European chondroitin sulfate and glucosamine food supplements: A systematic quality and quantity assessment compared to pharmaceuticals. Carbohydr. Polym. 2019, 222, 114984. [Google Scholar] [CrossRef] [PubMed]
- Lubis, A.M.T.; Siagian, C.; Wonggokusuma, E.; Marsetyo, A.F.; Setyohadi, B. Comparison of glucosamine-chondroitin sulfate with and without methylsulfonylmethane in grade I-II Knee osteoarthritis: A double blind randomized controlled trial. Acta Med. Indones. 2017, 49, 105–111. [Google Scholar] [PubMed]
- Honvo, G.; Bruyère, O.; Reginster, J.Y. Update on the role of pharmaceutical-grade chondroitin sulfate in the symptomatic management of knee osteoarthritis. Aging Clin. Exp. Res. 2019, 8, 1163–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reginster, J.Y.; Veronese, N. Highly purified chondroitin sulfate: A literature review on clinical efficacy and pharmacoeconomic aspects in osteoarthritis treatment. Aging Clin. Exp. Res. 2021, 33, 37–47. [Google Scholar] [CrossRef]
- Liu, X.; Machado, G.C.; Eyles, J.P.; Ravi, V.; Hunter, D.J. Dietary supplements for treating osteoarthritis: A systematic review and meta-analysis. Br. J. Sports Med. 2018, 52, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Sellam, J.; Courties, A.; Eymard, F.; Ferrero, S.; Latourte, A.; Ornetti, P.; Bannwarth, B.; Baumann, L.; Berenbaum, F.; Chevalier, X.; et al. French society of rheumatology. recommendations of the french society of rheumatology on pharmacological treatment of knee osteoarthritis. Jt. Bone Spine 2020, 87, 548–555. [Google Scholar] [CrossRef]
- Castrogiovanni, P.; Trovato, F.M.; Loreto, C.; Nsir, H.; Szychlinska, M.A.; Musumeci, G. Nutraceutical supplements in the management and prevention of osteoarthritis. Int. J. Mol. Sci. 2016, 17, 2042. [Google Scholar] [CrossRef]
- Dwyer, J.T.; Coates, P.M.; Smith, M.J. Dietary supplements: Regulatory challenges and research resources. Nutrients 2018, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Frankos, V.H.; Street, D.A.; O’Neill, R.K. FDA regulation of dietary supplements and requirements regarding adverse event reporting. Clin Pharmacol Ther. 2010, 87, 239–244. [Google Scholar] [CrossRef]
- Restaino, O.F.; D’ambrosio, S.; Cassese, E.; Barbuto Ferraiuolo, S.; Alfano, A.; Ventriglia, R.; Marrazzo, A.; Schiraldi, C.; Cimini, D. Molecular weight determination of heparosan- and chondroitin-like capsular polysaccharides: Figuring out differences between wild-type and engineered Escherichia coli strains. Appl. Microbiol. Biotechnol. 2019, 103, 6771–6782. [Google Scholar] [CrossRef]
- Stellavato, A.; Restaino, O.F.; Vassallo, V.; Finamore, R.; Ruosi, C.; Cassese, E.; De Rosa, M.; Schiraldi, C. Comparative analyses of pharmaceuticals or food supplements containing chondroitin sulfate: Are their bioactivities equivalent? Adv. Ther. 2019, 36, 3221–3237. [Google Scholar] [CrossRef] [Green Version]
- Volpi, N. Chondroitin sulfate safety and quality. Molecules 2019, 24, 1447. [Google Scholar] [CrossRef] [Green Version]
- Restaino, O.F.; Finamore, R.; Diana, P.; Marseglia, M.; Vitiello, M.; Casillo, A.; Bedini, E.; Parrilli, M.; Corsaro, M.M.; Trifuoggi, M.; et al. A multi-analytical approach to better assess the keratan sulfate contamination in animal origin chondroitin sulfate. Anal. Chim. Acta 2017, 958, 59–70. [Google Scholar] [CrossRef]
- Restaino, O.F.; De Rosa, M.; Schiraldi, C. High performance capillary electrophoresis to determine intact keratan sulfate and hyaluronic acid in animal origin chondroitin sulfate samples and food supplements. Electrophoresis 2020, 41, 1740–1748. [Google Scholar] [CrossRef]
- Stellavato, A.; Vassallo, V.; La Gatta, A. Novel hybrid gels made of high and low molecular weight hyaluronic acid induce proliferation and reduce inflammation in an osteoarthritis in vitro model based on human synoviocytes and chondrocytes. Biomed. Res. Int. 2019, 23, 4328219. [Google Scholar] [CrossRef] [Green Version]
- Calamia, V.; Fernández-Puente, P.; Mateos, J.; Lourido, L.; Rocha, B.; Montell, E.; Vergés, J.; Ruiz-Romero, C.; Blanco, F.J. Pharmacoproteomic study of three different chondroitin sulfate compounds on intracellular and extracellular human chondrocyte proteomes. Mol. Cell. Proteom. 2012, 11, M111.013417. [Google Scholar] [CrossRef] [Green Version]
- United States Pharmacopeia; USP 39-NF34. 2015. Available online: https://www.usp.org/ (accessed on 6 April 2021).
- Volpi, N. Disaccharide mapping of chondroitin sulfate of different origins by high performance capillary electrophoresis and high performance liquid chromatography. Carbohydr. Polym. 2004, 55, 273–281. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Balance, S.; Bohn, B.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014, 5, 1113. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Qin, Y.; Guan, R.; Zheng, W.; Liu, J.; Zhao, J. Digestibility of fucosylated glycosaminoglycan from sea cucumber and its effects on digestive enzymes under simulated salivary and gastrointestinal conditions. Carbohydr. Polym. 2018, 186, 217–255. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods. 1983, 16, 55–63. [Google Scholar] [CrossRef]
- Srivastava, G.K.; Alonso-Alonso, M.L.; Fernandez-Bueno, I.; Garcia-Gutierrez, M.T.; Rull, F.; Medina, J.; Coco, R.M.; Pastor, J.C. Comparison between direct contact and extract exposure methods for PFO cytotoxicity evaluation. Sci. Rep. 2018, 23, 1425. [Google Scholar] [CrossRef]
- D’Agostino, A.; Stellavato, A.; Busico, T.; Papa, A.; Tirino, V.; Papaccio, G.; La Gatta, A.; De Rosa, M.; Schiraldi, C. In vitro analysis of the effects on wound healing of high- and low-molecular weight chains of hyaluronan and their hybrid H-HA/L-HA complexes. BMC Cell Biol. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Stellavato, A.; Tirino, V.; de Novellis, F.; Della Vecchia, A.; Cinquegrani, F.; De Rosa, M.; Papaccio, G.; Schiraldi, C. Biotechnological chondroitin a novel glycosamminoglycan with remarkable biological function on human primary chondrocytes. J. Cell. Biochem. 2016, 117, 2158–2169. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.Q.; Feng, Z.H.; Li, X.B.; Hu, Z.C.; Xuan, J.W.; Wang, X.Y.; Xu, H.C.; Chen, J.X. Downregulating PI3K/Akt/NF-κB signaling with allicin for ameliorating the progression of osteoarthritis: In vitro and vivo studies. Food Funct. 2018, 19, 4865–4875. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N.; Maccari, F.; Linhardt, R.J. Capillary electrophoresis of complex natural polysaccharides. Electrophoresis 2008, 15, 3095–3106. [Google Scholar] [CrossRef] [Green Version]
- Lamari, F.N.; Militsopoulou, M.; Mitropoulou, T.N.; Hjerpe, A.; Karamanos, N.K. Analysis of glycosaminoglycan-derived disaccharides in biologic samples by capillary electrophoresis and protocol for sequencing glycosaminoglycans. Biomed. Chromatogr. 2002, 16, 95–102. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, F.; Linhardt, R.J. Analysis of the glycosaminoglycan chains of proteoglycans. J. Histochem. Cytochem. 2021, 69, 121–135. [Google Scholar] [CrossRef]
- Lacey, D.; Sampey, A.; Mitchell, R.; Bucala, R.; Santos, L.; Leech, M.; Morand, E. Control of fibroblast-like synoviocyte proliferation by macrophage migration inhibitory factor. Arthritis Rheum. 2003, 48, 103–109. [Google Scholar] [CrossRef]
- Yang, P.; Tan, J.; Yuan, Z.; Meng, G.; Bi, L.; Liu, J. Expression profile of cytokines and chemokines in osteoarthritis patients: Proinflammatory roles for CXCL8 and CXCL11 to chondrocytes. Int. Immunopharmacol. 2016, 40, 16–23. [Google Scholar] [CrossRef]
- El-Arman, M.M.; El-Fayoumi, G.; El-Shal, E.; El-Boghdady, I.; El-Ghaweet, A. Aggrecan and cartilage oligomeric matrix protein in serum and synovial fluid of patients with knee osteoarthritis. HSS J. 2010, 6, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Shigaku, S.; Keishi, M.; Takaaki, T.; Naoki, I.; Koji, K.; Katsuyuki, F. Expression analysis of three isoforms of hyaluronan synthase and hyaluronidase in the synovium of knees in osteoarthritis and rheumatoid arthritis by quantitative real-time reverse transcriptase polymerase chain reaction. Arthritis Res. Ther. 2004, 6, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David-Raoudi, M.; Deschrevel, B.; Leclercq, S.; Galéra, P.; Boumediene, K.; Pujol, J.P. Chondroitin sulfate increases hyaluronan production by human synoviocytes through differential regulation of hyaluronan synthases: Role of p38 and Akt. Arthritis Rheum. 2009, 3, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Bishnoi, M.; Jain, A.; Hurkat, P.; Jain, S.K. Chondroitin sulphate: A focus on osteoarthritis. Glycoconj. J. 2016, 33, 693–705. [Google Scholar] [CrossRef]
- Terabe, K.; Ohashi, Y.; Tsuchiya, S.; Ishizuka, S.; Knudson, C.B.; Knudson, W. Chondroprotective effects of 4-methylumbelliferone and hyaluronan synthase-2 overexpression involve changes in chondrocyte energy metabolism. J. Biol. Chem. 2019, 22, 17799–17817. [Google Scholar] [CrossRef]
- Zhao, Q.H.; Lin, L.P.; Guo, Y.X.; Zou, R.; Wang, Z.; Shi, Z.P.; Fu-Qing, L. Matrix metalloproteinase-13, NF-κB p65 and interleukin-1β are associated with the severity of knee osteoarthritis. Exp. Ther. Med. 2020, 19, 3620–3626. [Google Scholar] [CrossRef] [Green Version]
- Yoshihiro, N.; Cheryl, B.K.; Warren, K. Osteogenic Protein-1 inhibits matrix depletion in a hyaluronan hexasaccharide-induced model of osteoarthritis. Osteoarthr. Cartil. 2004, 5, 374–382. [Google Scholar]
- Igarashi, M.; Kaga, I.; Takamori, Y.; Sakamoto, K.; Miyazawa, K.; Nagaoka, I. Effects of glucosamine derivatives and uronic acids on the production of glycosaminoglycans by human synovial cells and chondrocytes. Int. J. Mol. Med. 2011, 27, 821–827. [Google Scholar]
- Ono, Y.; Sakai, T.; Hiraiwa, H.; Hamada, T.; Omachi, T.; Nakashima, M.; Ishizuka, S.; Matsukawa, T.; Knudson, W.; Knudson, C.B.; et al. Chondrogenic capacity and alterations in hyaluronan synthesis of cultured human osteoarthritic chondrocytes. Biochem. Biophys. Res. Commun. 2013, 14, 733–739. [Google Scholar] [CrossRef] [Green Version]
- Hiscock, D.R.; Caterson, B.; Flannery, C.R. Expression of hyaluronan synthases in articular cartilage. Osteoarthr. Cartil. 2000, 8, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Manicourt, D.H.; Pita, J.C. Progressive depletion of hyaluronic acid in early experimental osteoarthritis in dogs. Arthritis Rheum. 1988, 31, 538–544. [Google Scholar] [CrossRef]
- Padeh, S.; Farzam, N.; Chayen, G.; Gerstein, M.; Berkun, Y. Pentraxin 3 is a marker of early joint inflammation in patients with juvenile idiopathic arthritis. Immunol. Res. 2013, 56, 444–450. [Google Scholar] [CrossRef]
- Satomura, K.; Torigoshi, T.; Koga, T.; Maeda, Y.; Izumi, Y.; Jiuchi, Y.; Miyashita, T.; Yamasaki, S.; Kawakami, A.; Aiba, Y.; et al. Serum amyloid A (SAA) induces pentraxin 3 (PTX3) production in rheumatoid synoviocytes. Mod. Rheumatol. 2013, 23, 28–35. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CS (mg) | GlcN (mg) | HA (mg) |
|---|---|---|---|
| PhA | 400.0 | - | - |
| PhB | 400.0 | - | - |
| US FS1 | 400.0 | 500.0 | - |
| US FS2 | 400.0 | 500.0 | - |
| US FS3 | 100.0 | 750.0 | 1.65 |
| Pharmaceuticals | % dis-0S | % dis-6S | % dis-4S | % dis-2S | % dis-2,6S | % dis-4,6S | % dis-2,4S | % dis-4S/6S | CS Animal Origin | CS Mw (KDa) | CS Mw/Mn |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CS | CS | CS | CS | CS | CS | CS | CS | ||||
| Ph A | 2.12 | 15.49 | 82.38 | n.d. | n.d. | n.d. | n.d. | 5.32 | Pig | 19.00 | 1.22 |
| Ph B | n.d. | 64.21 | 26.87 | n.d. | 5.3 | 3.61 | n.d. | 0.42 | Fish | 36.22 | 1.23 |
| Food supplements | % dis-0S | % dis-6S | % dis-4S | % dis-2S | % dis-2,6S | % dis-4,6S | % dis-2,4S CS | % dis-4S/6S CS | CS Animal Origin | CS Mw (KDa) | CS Mw/Mn |
| CS | CS | CS | CS | CS | CS | ||||||
| US FS1 | 4.06 | 18.71 | 71.61 | 1.04 | n.d. | n.d. | 4.56 | 3.83 | Pig | 18.71 | 1.27 |
| US FS2 | 1.81 | 15.55 | 52.99 | 4.93 | 14.6 | 10.11 | n.d. | 3.41 | Mixed Terrestrial/ Marine | 62.06/28.6 | 1.16/1.01 |
| US FS3 | 15.61 * | 15.21 | 53.88 | n.d. | 5.79 | 8.52 | 1.08 | 3.56 | Mixed Terrestrial/ Marine | 375.00 */ 22.8 | 1.49 */1.25 |
| % CS Post Gastric Digestion | % CS Post Intestinal Digestion | % Total CS Post Full Digestion | ||||
|---|---|---|---|---|---|---|
| Pharmaceuticals | % average CS | % std dev CS | % average CS | % std dev CS | % average CS | % std dev CS |
| Ph A | 95.44 | 4.22 | 91.35 | 1.60 | 87.19 | 0.07 |
| Ph B | 94.05 | 5.95 | 96.60 | 3.34 | 90.85 | 0.20 |
| Food supplements | % average CS | % std dev CS | % average CS | % std dev CS | % average CS | % std dev CS |
| US FS1 | 96.17 | 3.34 | 99.60 | 0.31 | 95.78 | 0.01 |
| US FS2 | 95.54 | 4.01 | 94.49 | 2.20 | 90.27 | 0.09 |
| US FS3 | 99.63 | 0.37 | 95.21 | 3.57 | 94.86 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stellavato, A.; Restaino, O.F.; Vassallo, V.; Cassese, E.; Finamore, R.; Ruosi, C.; Schiraldi, C. Chondroitin Sulfate in USA Dietary Supplements in Comparison to Pharma Grade Products: Analytical Fingerprint and Potential Anti-Inflammatory Effect on Human Osteoartritic Chondrocytes and Synoviocytes. Pharmaceutics 2021, 13, 737. https://doi.org/10.3390/pharmaceutics13050737
Stellavato A, Restaino OF, Vassallo V, Cassese E, Finamore R, Ruosi C, Schiraldi C. Chondroitin Sulfate in USA Dietary Supplements in Comparison to Pharma Grade Products: Analytical Fingerprint and Potential Anti-Inflammatory Effect on Human Osteoartritic Chondrocytes and Synoviocytes. Pharmaceutics. 2021; 13(5):737. https://doi.org/10.3390/pharmaceutics13050737
Chicago/Turabian StyleStellavato, Antonietta, Odile Francesca Restaino, Valentina Vassallo, Elisabetta Cassese, Rosario Finamore, Carlo Ruosi, and Chiara Schiraldi. 2021. "Chondroitin Sulfate in USA Dietary Supplements in Comparison to Pharma Grade Products: Analytical Fingerprint and Potential Anti-Inflammatory Effect on Human Osteoartritic Chondrocytes and Synoviocytes" Pharmaceutics 13, no. 5: 737. https://doi.org/10.3390/pharmaceutics13050737