
The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas aeruginosa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Bacterial Strains
2.3. Preparation of Liposomes
2.4. Microbiological Assay for the Measurement of Gentamicin and Erythromycin in Liposomes
2.5. Determination of Encapsulation Efficiency
2.6. Determination of Minimum Inhibitory Concentrations (MICs) and Minimum Bactericidal Concentrations (MBCs)
2.7. Minimum Biofilm Eradication Concentration (MBEC)
2.8. Effects of Sub-Inhibitory Concentrations of Free and Liposomal Gentamicin and Erythromycin on the Growth of P. aeruginosa
2.9. Virulence Factor Assays
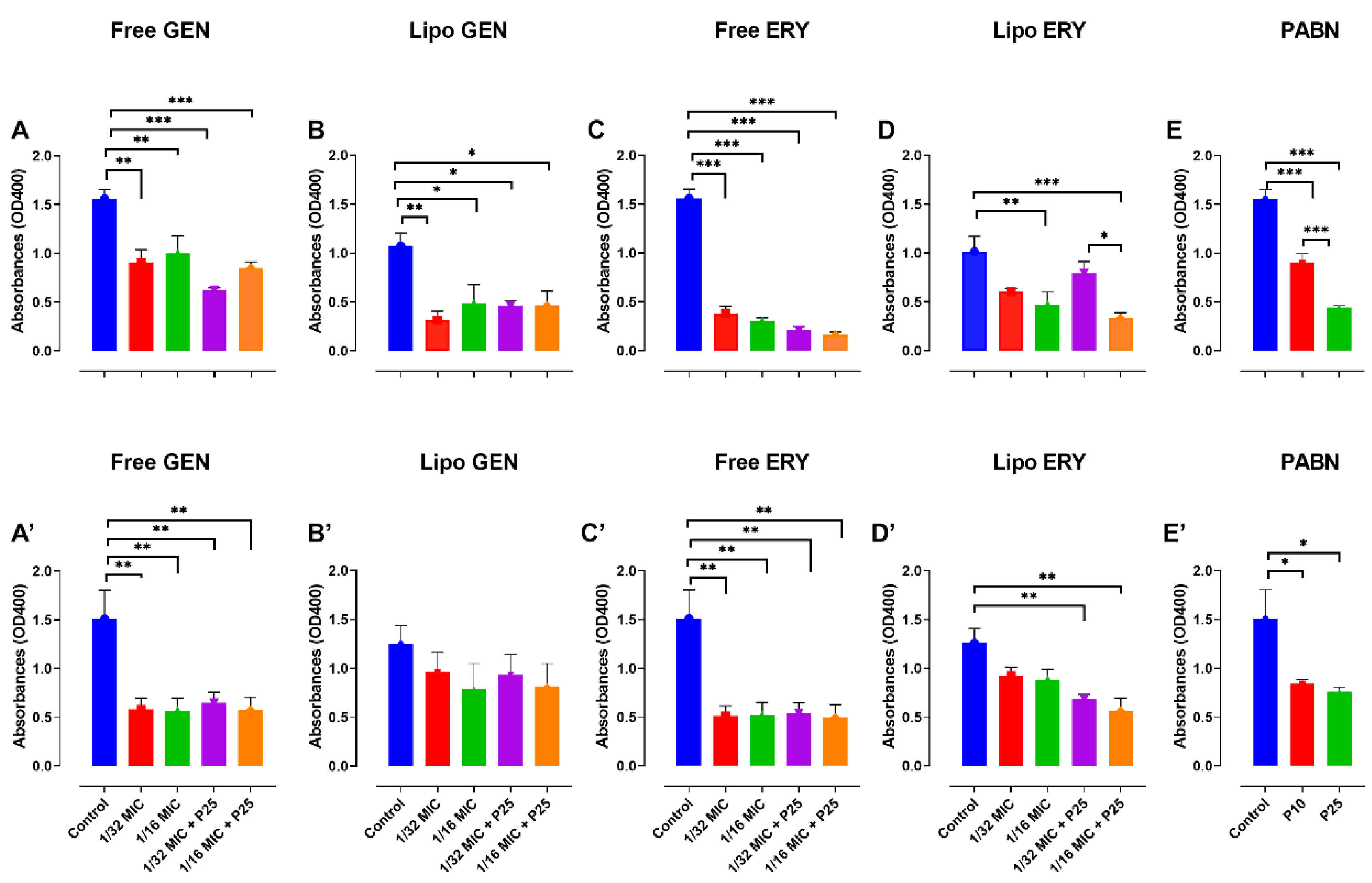
2.9.1. Protease Assay
2.9.2. Pyocyanin and Pyoverdine Assays
2.9.3. Lipase Assay
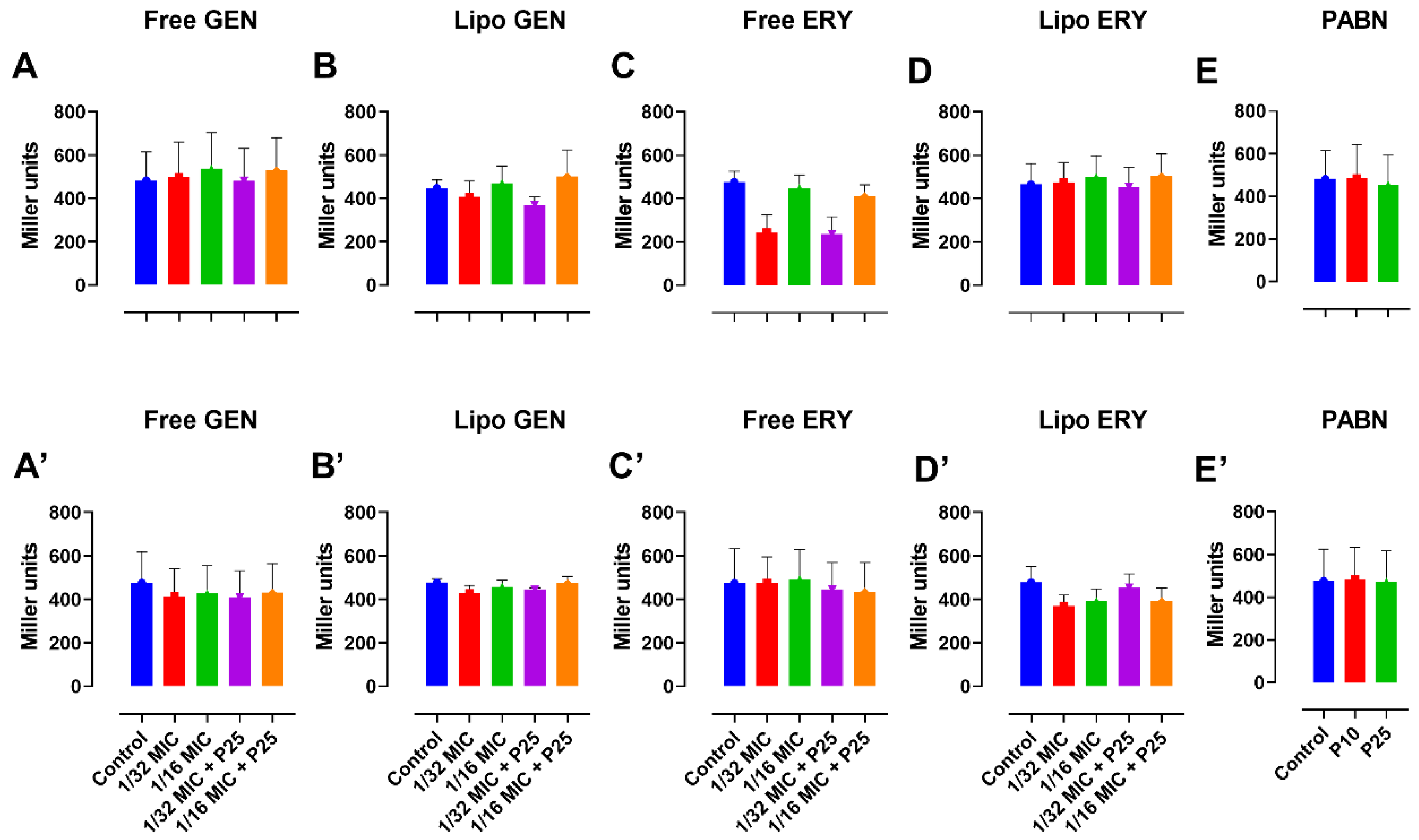
2.10. Beta-Galactosidase Activity Assay
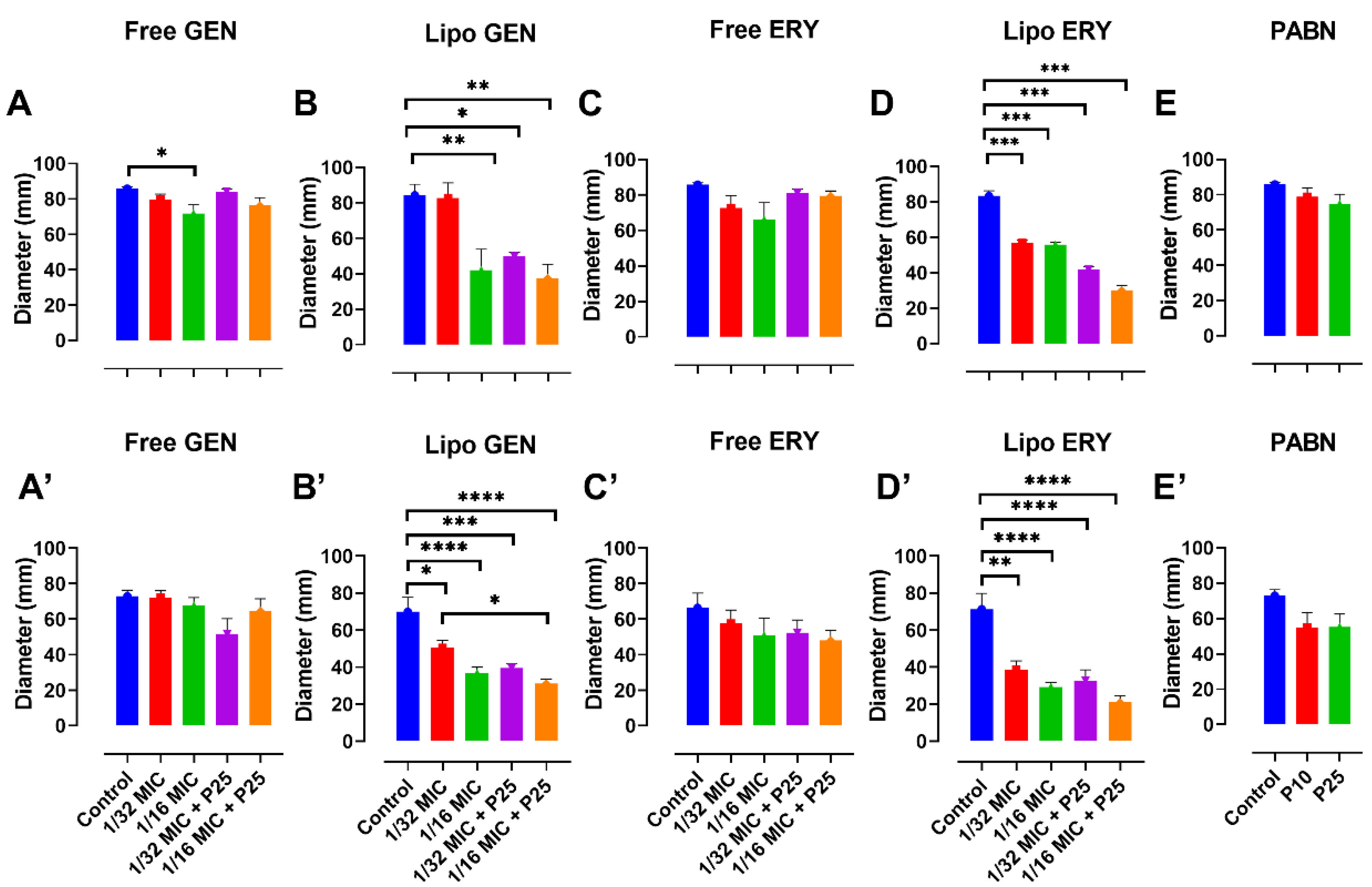
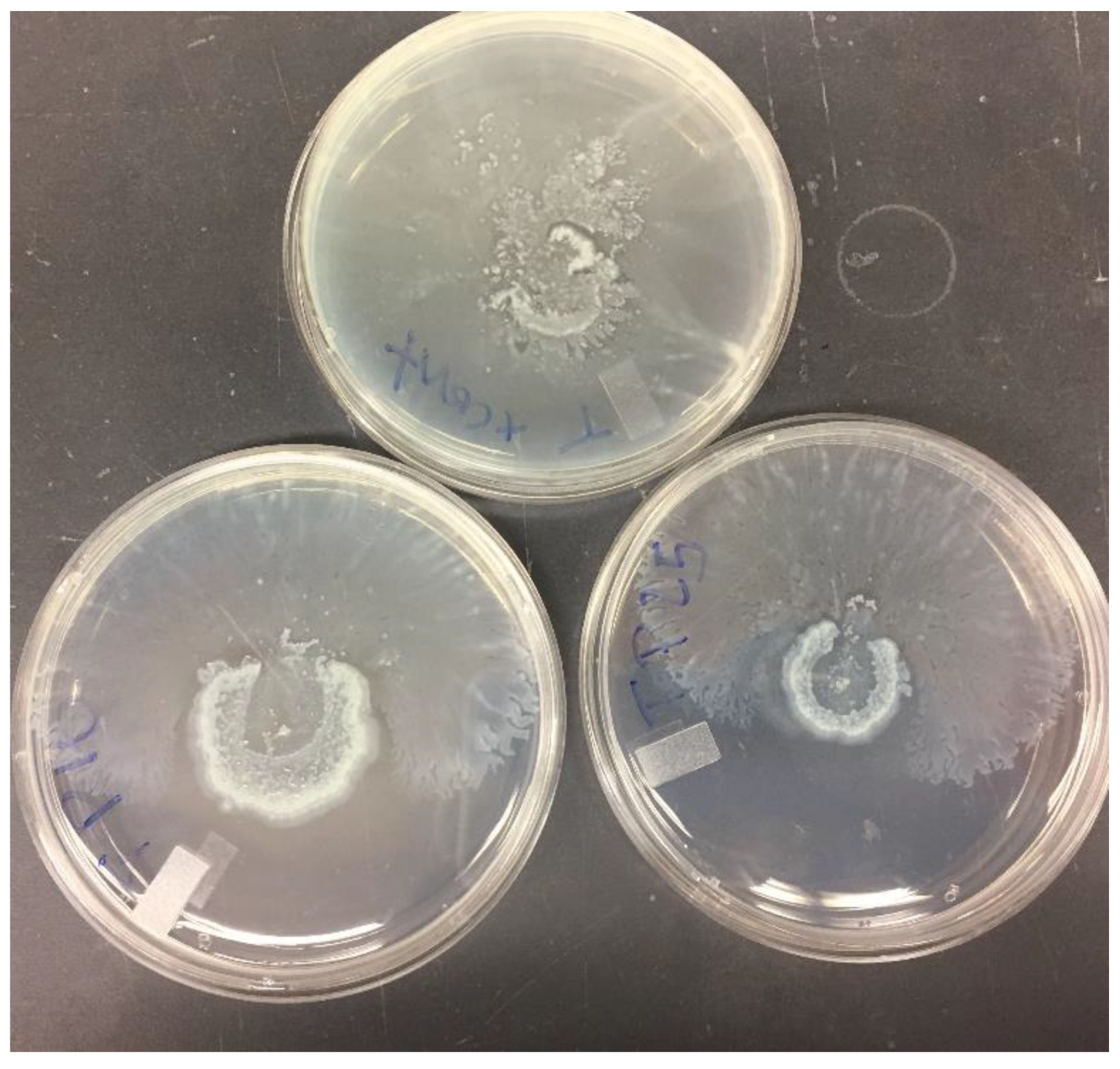
2.11. Motility Studies
2.12. Data Analysis
3. Results
3.1. Liposomal Antibiotics Characterisations
3.2. Determination of MICs, MBCs and MBECs
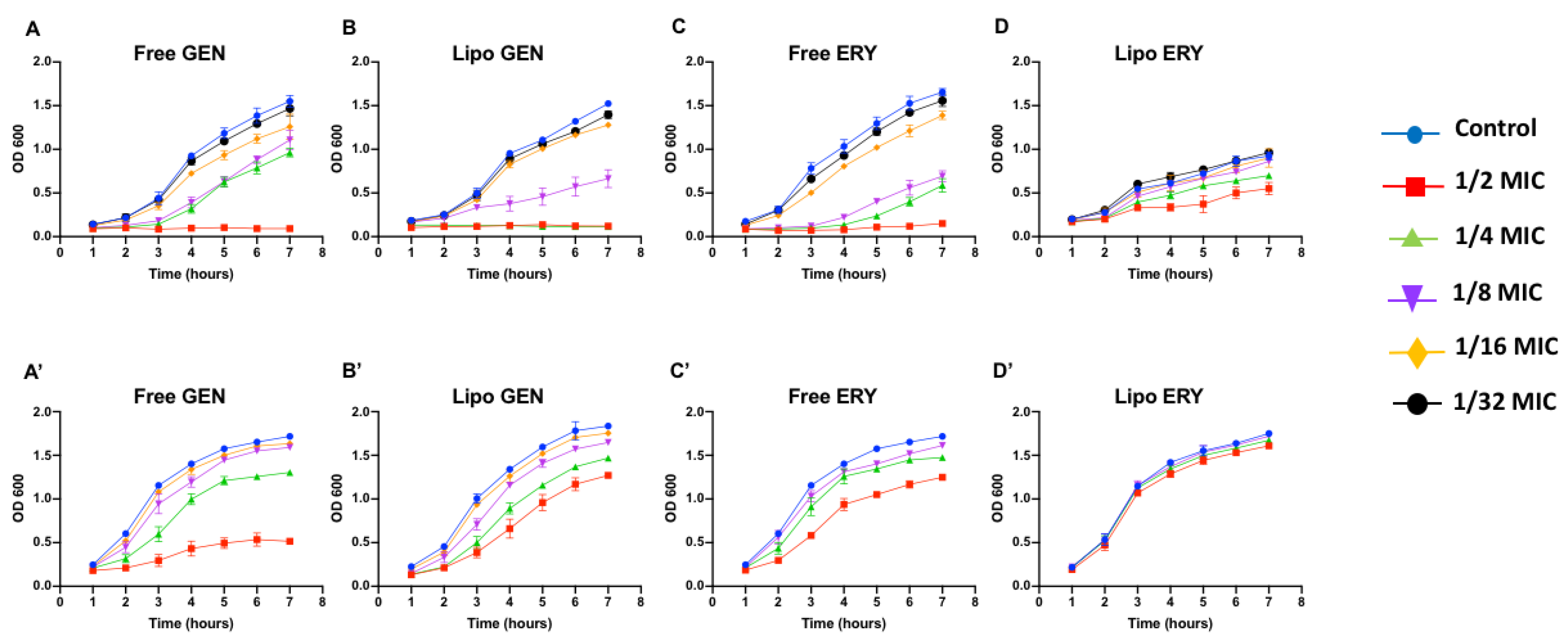
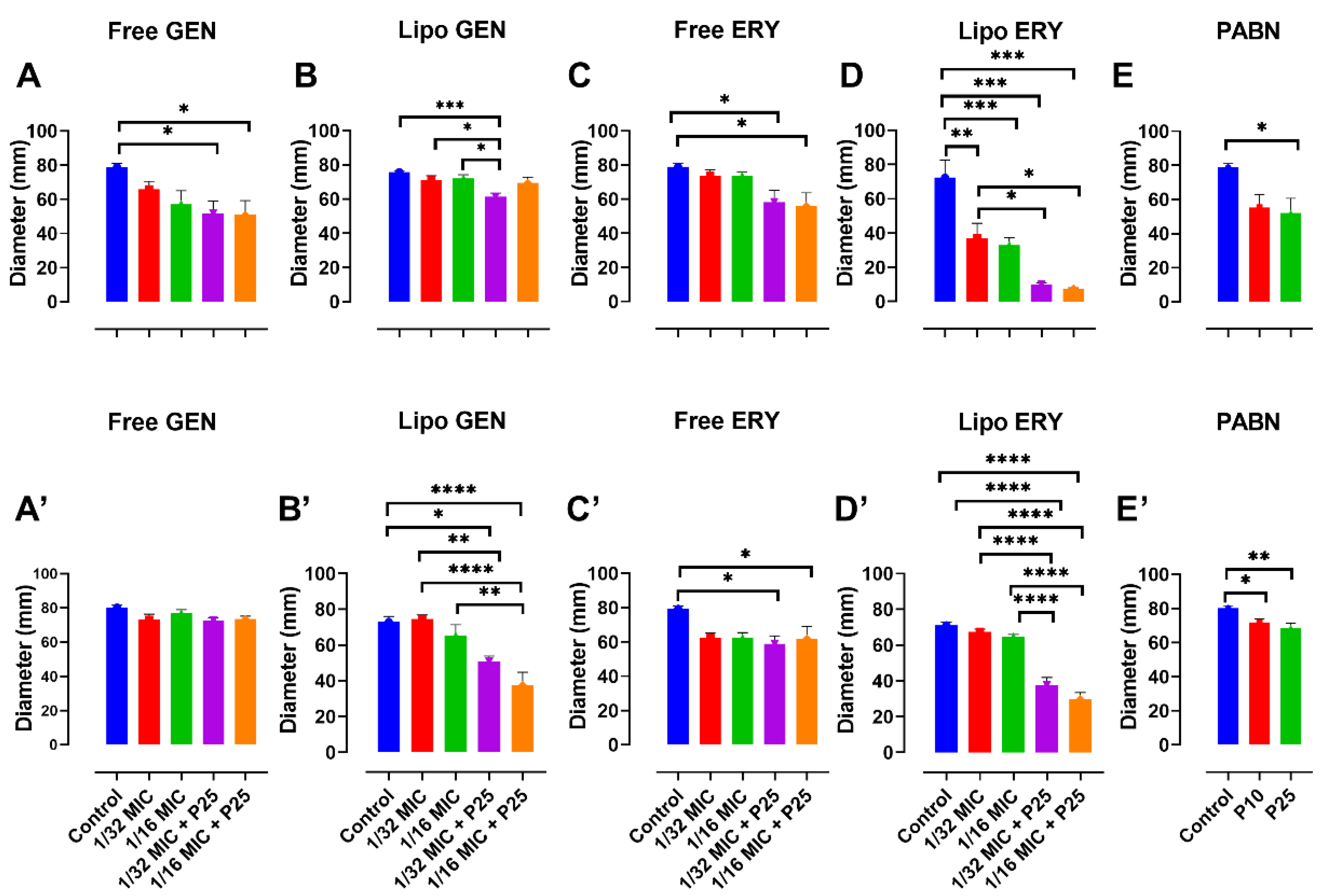
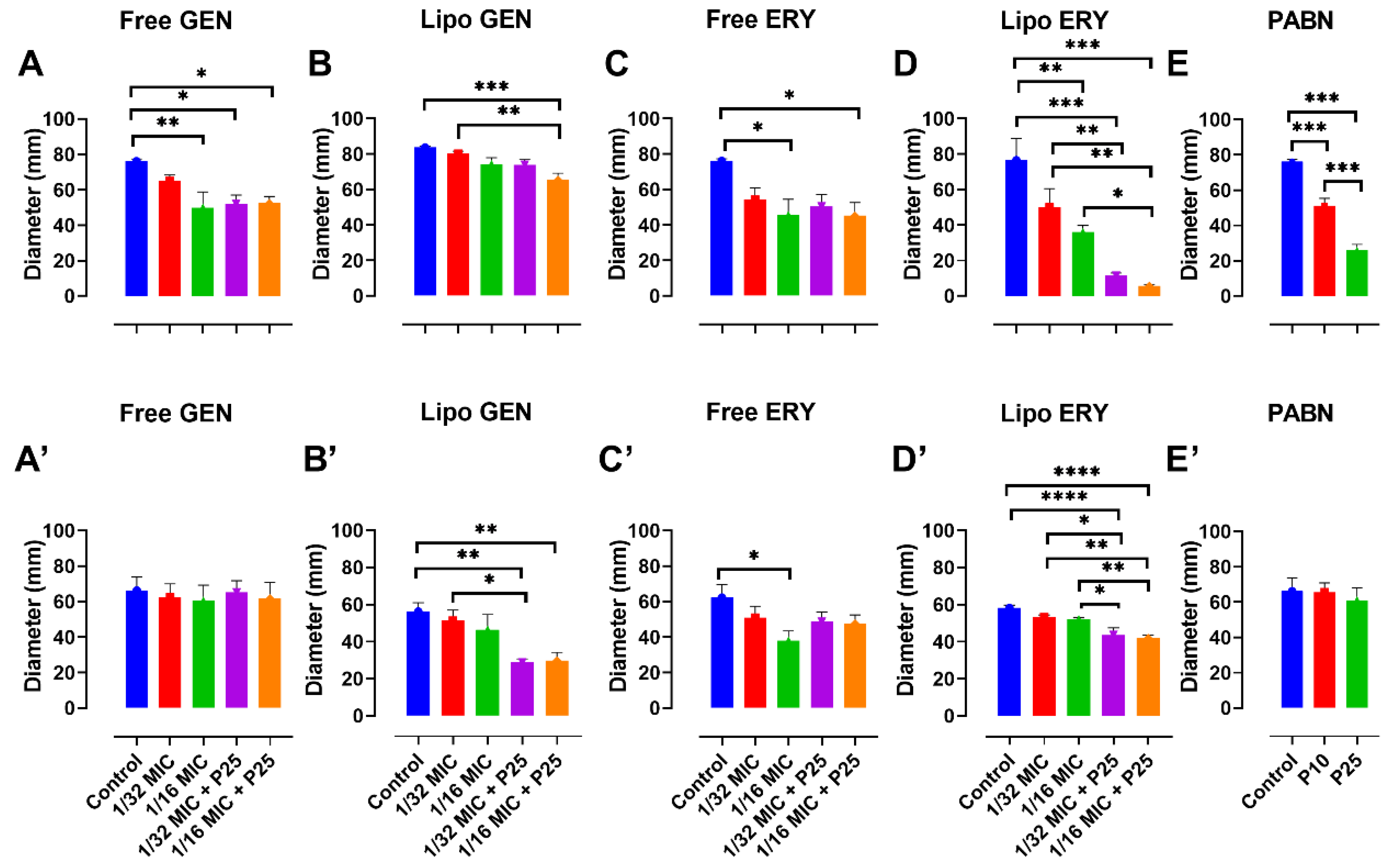
3.3. Effects of Sub-Inhibitory Concentrations of Free and Liposomal Antibiotics on the Growth of P. aeruginosa Strains
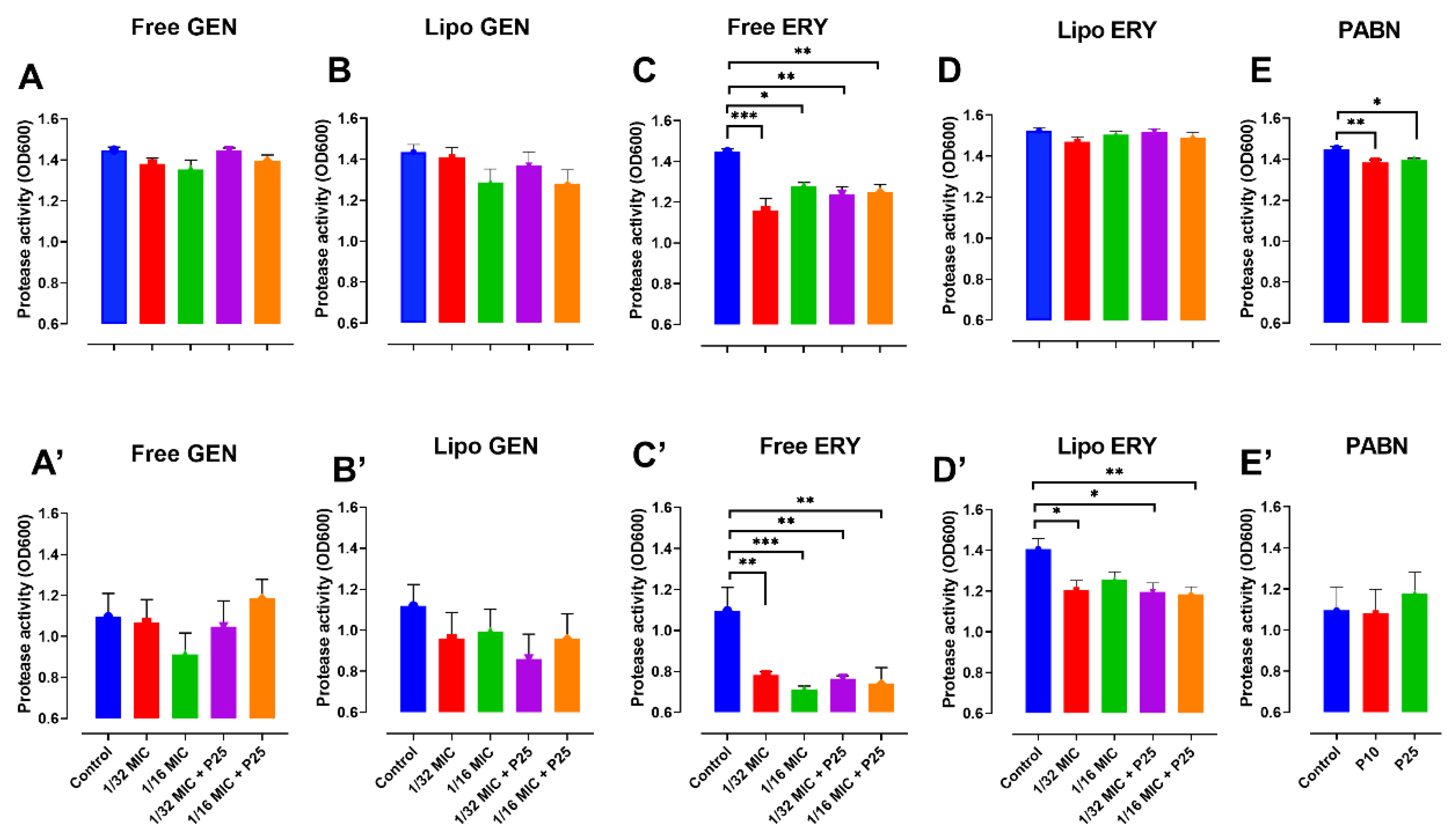
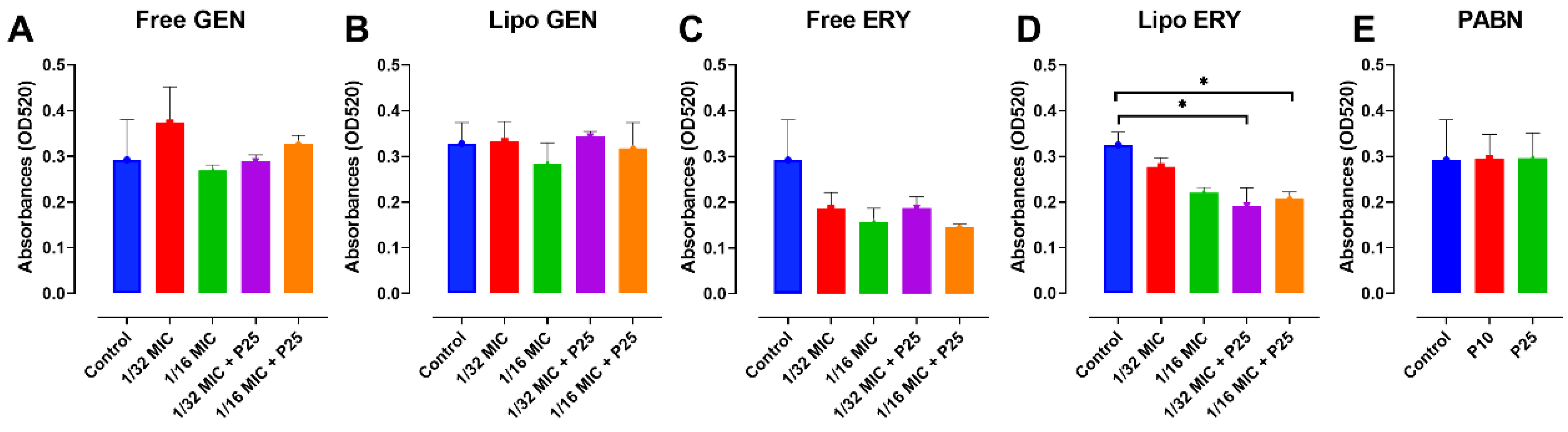
3.4. Effect of Antibiotics and PABN on Bacterial Virulence Factors
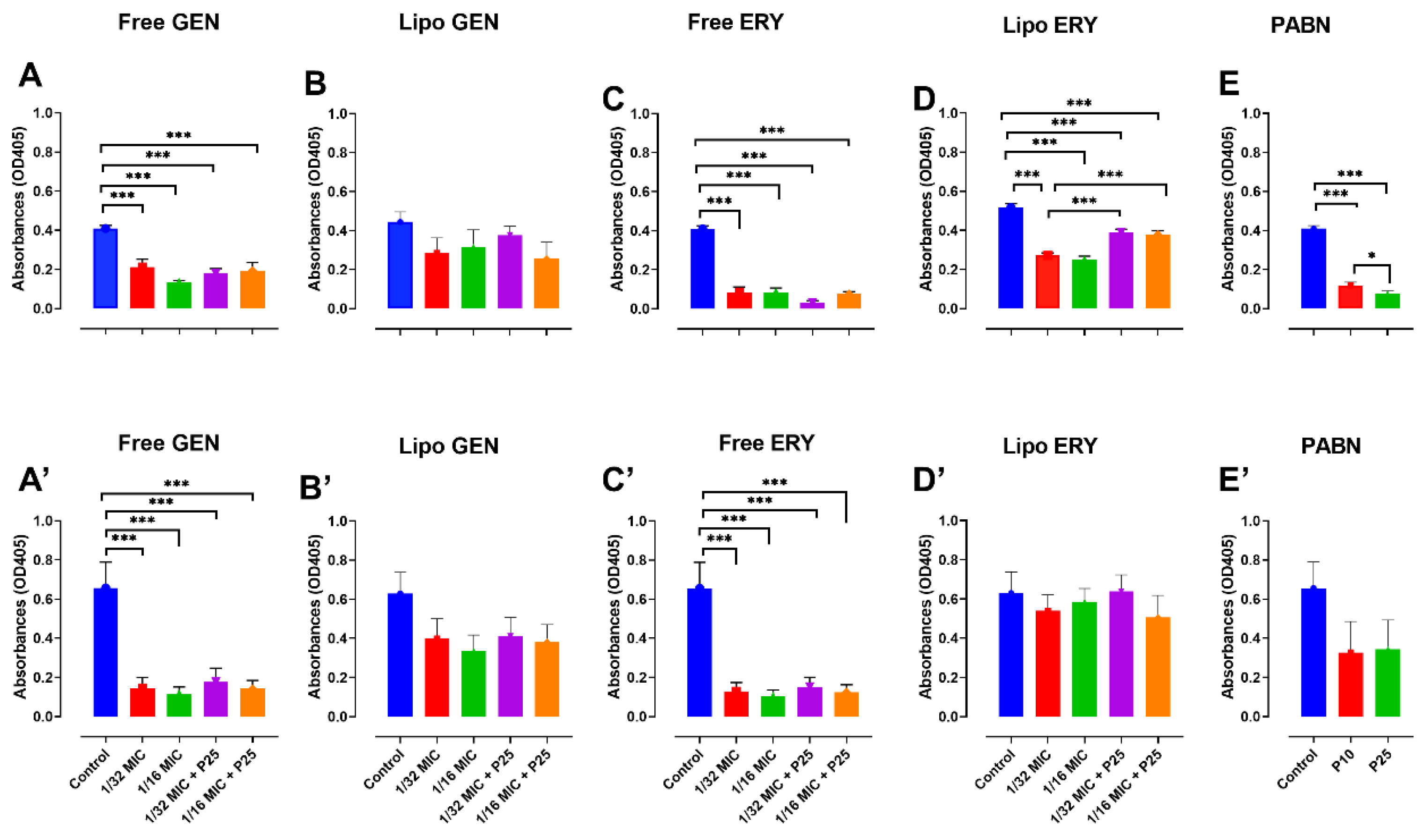
3.5. Assessment of Quorum-Sensing Signal Production through a Beta-Galactosidase Assay
3.6. Effect of Antibiotics and PABN on Bacterial Motility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malhotra, S.; Hayes, D.; Wozniak, D.J. Mucoid Pseudomonas Aeruginosa and Regional Inflammation in the Cystic Fibrosis Lung. J. Cyst. Fibros. 2019, 18, 796–803. [Google Scholar] [CrossRef]
- Abdelghany, S.M.; Quinn, D.J.; Ingram, R.J.; Gilmore, B.F.; Donnelly, R.F.; Taggart, C.C.; Scott, C.J. Gentamicin-Loaded Nanoparticles Show Improved Antimicrobial Effects towards Pseudomonas Aeruginosa Infection. Int. J. Nanomed. 2012, 7, 4053–4063. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, A.P.; Azevedo, N.F.; Pereira, M.O.; Lopes, S.P. The Cystic Fibrosis Microbiome in an Ecological Perspective and Its Impact in Antibiotic Therapy. Appl. Microbiol. Biotechnol. 2016, 100, 1163–1181. [Google Scholar] [CrossRef]
- López-Causapé, C.; Rubio, R.; Cabot, G.; Oliver, A. Evolution of the Pseudomonas Aeruginosa Aminoglycoside Mutational Resistome In Vitro and in the Cystic Fibrosis Setting. Antimicrob. Agents Chemother. 2018, 62, e02583-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassetti, M.; Castaldo, N.; Cattelan, A.; Mussini, C.; Righi, E.; Tascini, C.; Menichetti, F.; Mastroianni, C.M.; Tumbarello, M.; Grossi, P.; et al. Ceftolozane/Tazobactam for the Treatment of Serious Pseudomonas Aeruginosa Infections: A Multicentre Nationwide Clinical Experience. Int. J. Antimicrob. Agents 2019, 53, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; Widlake, E.; Teo, J.; Huseby, D.L.; Tyrrell, J.M.; Polikanov, Y.S.; Ercan, O.; Petersson, A.; Cao, S.; Aboklaish, A.F.; et al. In Vitro Activity of Apramycin against Multidrug-, Carbapenem- and Aminoglycoside-Resistant Enterobacteriaceae and Acinetobacter Baumannii. J. Antimicrob. Chemother. 2019, 74, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.; Lee, J.-W.; Javaid, A.; Park, S.-K.; Kim, Y.-M. Inhibition of Biofilm and Virulence Properties of Pseudomonas Aeruginosa by Sub-Inhibitory Concentrations of Aminoglycosides. Microb. Pathog. 2020, 146, 104249. [Google Scholar] [CrossRef]
- Hoshino, K.; Imai, Y.; Mukai, K.; Hamauzu, R.; Ochi, K.; Hosaka, T. A Putative Mechanism Underlying Secondary Metabolite Overproduction by Streptomyces Strains with a 23S RRNA Mutation Conferring Erythromycin Resistance. Appl. Microbiol. Biotechnol. 2020, 104, 2193–2203. [Google Scholar] [CrossRef]
- Chalmers, J.D.; Boersma, W.; Lonergan, M.; Jayaram, L.; Crichton, M.L.; Karalus, N.; Taylor, S.L.; Martin, M.L.; Burr, L.D.; Wong, C.; et al. Long-Term Macrolide Antibiotics for the Treatment of Bronchiectasis in Adults: An Individual Participant Data Meta-Analysis. Lancet Respir. Med. 2019, 7, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Mayer-Hamblett, N.; Retsch-Bogart, G.; Kloster, M.; Accurso, F.; Rosenfeld, M.; Albers, G.; Black, P.; Brown, P.; Cairns, A.; Davis, S.D.; et al. Azithromycin for Early Pseudomonas Infection in Cystic Fibrosis. The OPTIMIZE Randomized Trial. Am. J. Respir. Crit. Care Med. 2018, 198, 1177–1187. [Google Scholar] [CrossRef]
- Lababidi, N.; Ofosu Kissi, E.; Elgaher, W.A.M.; Sigal, V.; Haupenthal, J.; Schwarz, B.C.; Hirsch, A.K.H.; Rades, T.; Schneider, M. Spray-Drying of Inhalable, Multifunctional Formulations for the Treatment of Biofilms Formed in Cystic Fibrosis. J. Control. Release 2019, 314, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Belotti, S.; Rossi, A.; Colombo, P.; Bettini, R.; Rekkas, D.; Politis, S.; Colombo, G.; Balducci, A.G.; Buttini, F. Spray Dried Amikacin Powder for Inhalation in Cystic Fibrosis Patients: A Quality by Design Approach for Product Construction. Int. J. Pharm. 2014, 471, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Kordes, A.; Preusse, M.; Willger, S.D.; Braubach, P.; Jonigk, D.; Haverich, A.; Warnecke, G.; Häussler, S. Genetically Diverse Pseudomonas Aeruginosa Populations Display Similar Transcriptomic Profiles in a Cystic Fibrosis Explanted Lung. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sonnleitner, E.; Pusic, P.; Wolfinger, M.T.; Bläsi, U. Distinctive Regulation of Carbapenem Susceptibility in Pseudomonas Aeruginosa by Hfq. Front. Microbiol. 2020, 11, 1001. [Google Scholar] [CrossRef]
- Abbasi, F.; Yusefi, S.; Yavar, S.A. Minimum Inhibitory Concentration of Ciprofloxacin against Pseudomonas Aeruginosa in the Presence of the Efflux Inhibitor Phenylalanine-Arginine Beta-Naphthylamide. Intern. Med. Med. Investig. J. 2018, 3. [Google Scholar] [CrossRef]
- Pu, Y.; Ke, Y.; Bai, F. Active Efflux in Dormant Bacterial Cells – New Insights into Antibiotic Persistence. Drug Resist. Updat. 2017, 30, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas Aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Sans-Serramitjana, E.; Jorba, M.; Fusté, E.; Pedraz, J.L.; Vinuesa, T.; Viñas, M. Free and Nanoencapsulated Tobramycin: Effects on Planktonic and Biofilm Forms of Pseudomonas. Microorganisms 2017, 5, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetard, A.; Zedet, A.; Girard, C.; Plésiat, P.; Llanes, C. Cinnamaldehyde Induces Expression of Efflux Pumps and Multidrug Resistance in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e01081-19. [Google Scholar] [CrossRef] [Green Version]
- Tafti, F.A.; Eslami, G.; Zandi, H.; Barzegar, K. Mutations in Nalc Gene of Mex AB-OprM Efflux Pump in Carbapenem Resistant Pseudomonas Aeruginosa Isolated from Burn Wounds in Yazd, Iran. Iran J. Microbiol. 2020, 12, 32–36. [Google Scholar]
- Minagawa, S.; Inami, H.; Kato, T.; Sawada, S.; Yasuki, T.; Miyairi, S.; Horikawa, M.; Okuda, J.; Gotoh, N. RND Type Efflux Pump System MexAB-OprM of Pseudomonas Aeruginosa Selects Bacterial Languages, 3-Oxo-Acyl-Homoserine Lactones, for Cell-to-Cell Communication. BMC Microbiol. 2012, 12, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Sykes, E.M.; Li, Y.; Kumar, A. MexXY RND Pump of Pseudomonas Aeruginosa PA7 Effluxes Bi-Anionic β-Lactams Carbenicillin and Sulbenicillin When It Partners with the Outer Membrane Factor OprA but Not with OprM. Microbiology 2020, 166, 1095–1106. [Google Scholar] [CrossRef]
- Seupt, A.; Schniederjans, M.; Tomasch, J.; Häussler, S. Expression of the MexXY Aminoglycoside Efflux Pump and Presence of an Aminoglycoside-Modifying Enzyme in Clinical Pseudomonas Aeruginosa Isolates Are Highly Correlated. Antimicrob. Agents Chemother. 2020, 65, e01166-20. [Google Scholar] [CrossRef] [PubMed]
- Puja, H.; Bolard, A.; Noguès, A.; Plésiat, P.; Jeannot, K. The Efflux Pump MexXY/OprM Contributes to the Tolerance and Acquired Resistance of Pseudomonas Aeruginosa to Colistin. Antimicrob. Agents Chemother. 2020, 64, e02033-19. [Google Scholar] [CrossRef]
- Cruz, R.L.; Asfahl, K.L.; Van den Bossche, S.; Coenye, T.; Crabbé, A.; Dandekar, A.A. RhlR-Regulated Acyl-Homoserine Lactone Quorum Sensing in a Cystic Fibrosis Isolate of Pseudomonas Aeruginosa. mBio 2020, 11, e00532-20. [Google Scholar] [CrossRef] [Green Version]
- Gökalsın, B.; Aksoydan, B.; Erman, B.; Sesal, N.C. Reducing Virulence and Biofilm of Pseudomonas Aeruginosa by Potential Quorum Sensing Inhibitor Carotenoid: Zeaxanthin. Microb. Ecol. 2017, 74, 466–473. [Google Scholar] [CrossRef]
- Karthick Raja Namasivayam, S.; Angel, J.; Bharani, R.S.A.; Nachiyar, C.V. Terminalia Chebula and Ficus Racemosa Principles Mediated Repression of Novel Drug Target Las R—The Transcriptional Regulator and Its Controlled Virulence Factors Produced by Multiple Drug Resistant Pseudomonas Aeruginosa—Biocompatible Formulation against Drug Resistant Bacteria. Microb. Pathog. 2020, 148, 104412. [Google Scholar] [CrossRef]
- Ahmed, T.; Pattnaik, S.; Khan, M.B.; Ampasala, D.R.; Busi, S.; Sarma, V.V. Inhibition of Quorum Sensing–Associated Virulence Factors and Biofilm Formation in Pseudomonas Aeruginosa PAO1 by Mycoleptodiscus Indicus PUTY1. Braz. J. Microbiol. 2020, 51, 467–487. [Google Scholar] [CrossRef]
- Mishra, R.; Kushveer, J.S.; Khan, M.I.K.; Pagal, S.; Meena, C.K.; Murali, A.; Dhayalan, A.; Venkateswara Sarma, V. 2,4-Di-Tert-Butylphenol Isolated From an Endophytic Fungus, Daldinia Eschscholtzii, Reduces Virulence and Quorum Sensing in Pseudomonas Aeruginosa. Front. Microbiol. 2020, 11, 1668. [Google Scholar] [CrossRef]
- Armijo, L.M.; Wawrzyniec, S.J.; Kopciuch, M.; Brandt, Y.I.; Rivera, A.C.; Withers, N.J.; Cook, N.C.; Huber, D.L.; Monson, T.C.; Smyth, H.D.C.; et al. Antibacterial Activity of Iron Oxide, Iron Nitride, and Tobramycin Conjugated Nanoparticles against Pseudomonas Aeruginosa Biofilms. J. Nanobiotechnol. 2020, 18, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, A.; Roche, B.; Schalk, I.J. Iron Acquisition in Pseudomonas Aeruginosa by the Siderophore Pyoverdine: An Intricate Interacting Network Including Periplasmic and Membrane Proteins. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Harrington, N.E.; Sweeney, E.; Harrison, F. Building a Better Biofilm - Formation of in Vivo-like Biofilm Structures by Pseudomonas Aeruginosa in a Porcine Model of Cystic Fibrosis Lung Infection. Biofilm 2020, 2, 100024. [Google Scholar] [CrossRef]
- Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Social Behavior of Antibiotic Resistant Mutants Within Pseudomonas Aeruginosa Biofilm Communities. Front. Microbiol. 2019, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A Quorum-Sensing Inhibitor Blocks Pseudomonas Aeruginosa Virulence and Biofilm Formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [Green Version]
- Malgaonkar, A.; Nair, M. Quorum Sensing in Pseudomonas Aeruginosa Mediated by RhlR Is Regulated by a Small RNA PhrD. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sankar Ganesh, P.; Ravishankar Rai, V. Attenuation of Quorum-Sensing-Dependent Virulence Factors and Biofilm Formation by Medicinal Plants against Antibiotic Resistant Pseudomonas Aeruginosa. J. Tradit. Complement. Med. 2018, 8, 170–177. [Google Scholar] [CrossRef]
- Caldwell, C.C.; Chen, Y.; Goetzmann, H.S.; Hao, Y.; Borchers, M.T.; Hassett, D.J.; Young, L.R.; Mavrodi, D.; Thomashow, L.; Lau, G.W. Pseudomonas Aeruginosa Exotoxin Pyocyanin Causes Cystic Fibrosis Airway Pathogenesis. Am. J. Pathol. 2009, 175, 2473–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Managò, A.; Becker, K.A.; Carpinteiro, A.; Wilker, B.; Soddemann, M.; Seitz, A.P.; Edwards, M.J.; Grassmé, H.; Szabò, I.; Gulbins, E. Pseudomonas Aeruginosa Pyocyanin Induces Neutrophil Death via Mitochondrial Reactive Oxygen Species and Mitochondrial Acid Sphingomyelinase. Antioxid. Redox Signal. 2015, 22, 1097–1110. [Google Scholar] [CrossRef] [Green Version]
- Du, D.; Wang-Kan, X.; Neuberger, A.; Van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug Efflux Pumps: Structure, Function and Regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Kirienko, D.R.; Kang, D.; Kirienko, N.V. Novel Pyoverdine Inhibitors Mitigate Pseudomonas Aeruginosa Pathogenesis. Front. Microbiol. 2019, 9, 3317. [Google Scholar] [CrossRef] [Green Version]
- Kishk, R.M.; Abdalla, M.O.; Hashish, A.A.; Nemr, N.A.; El Nahhas, N.; Alkahtani, S.; Abdel-Daim, M.M.; Kishk, S.M. Efflux MexAB-Mediated Resistance in P. Aeruginosa Isolated from Patients with Healthcare Associated Infections. Pathogens 2020, 9, 471. [Google Scholar] [CrossRef]
- Ferrer-Espada, R.; Shahrour, H.; Pitts, B.; Stewart, P.S.; Sánchez-Gómez, S.; Martínez-de-Tejada, G. A Permeability-Increasing Drug Synergizes with Bacterial Efflux Pump Inhibitors and Restores Susceptibility to Antibiotics in Multi-Drug Resistant Pseudomonas Aeruginosa Strains. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- El-Shaer, S.; Shaaban, M.; Barwa, R.; Hassan, R. Control of Quorum Sensing and Virulence Factors of Pseudomonas Aeruginosa Using Phenylalanine Arginyl β-Naphthylamide. J. Med. Microbiol. 2016, 65, 1194–1204. [Google Scholar] [CrossRef]
- Lamers, R.P.; Cavallari, J.F.; Burrows, L.L. The Efflux Inhibitor Phenylalanine-Arginine Beta-Naphthylamide (PAβN) Permeabilizes the Outer Membrane of Gram-Negative Bacteria. PLoS ONE 2013, 8, e60666. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, M.; Kalhapure, R.S.; Rambharose, S.; Mocktar, C.; Singh, S.; Kodama, T.; Govender, T. Novel Lipids with Three C18-Fatty Acid Chains and an Amino Acid Head Group for PH-Responsive and Sustained Antibiotic Delivery. Chem. Phys. Lipids 2018, 212, 12–25. [Google Scholar] [CrossRef]
- Obuobi, S.; Julin, K.; Fredheim, E.G.A.; Johannessen, M.; Škalko-Basnet, N. Liposomal Delivery of Antibiotic Loaded Nucleic Acid Nanogels with Enhanced Drug Loading and Synergistic Anti-Inflammatory Activity against S. aureus Intracellular Infections. J. Control. Release 2020, 324, 620–632. [Google Scholar] [CrossRef]
- Alhariri, M.; Omri, A. Efficacy of Liposomal Bismuth-Ethanedithiol-Loaded Tobramycin after Intratracheal Administration in Rats with Pulmonary Pseudomonas Aeruginosa Infection. Antimicrob. Agents Chemother. 2013, 57, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Leifer, F.; Rose, S.; Chun, D.Y.; Thaisz, J.; Herr, T.; Nashed, M.; Joseph, J.; Perkins, W.R.; DiPetrillo, K. Amikacin Liposome Inhalation Suspension (ALIS) Penetrates Non-Tuberculous Mycobacterial Biofilms and Enhances Amikacin Uptake Into Macrophages. Front. Microbiol. 2018, 9, 915. [Google Scholar] [CrossRef] [Green Version]
- Cipolla, D.; Blanchard, J.; Gonda, I. Development of Liposomal Ciprofloxacin to Treat Lung Infections. Pharmaceutics 2016, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Rukholm, G.; Mugabe, C.; Azghani, A.O.; Omri, A. Antibacterial Activity of Liposomal Gentamicin against Pseudomonas Aeruginosa: A Time–Kill Study. Int. J. Pharm. 2006, 27, 247–252. [Google Scholar] [CrossRef]
- Alhajlan, M.; Alhariri, M.; Omri, A. Efficacy and Safety of Liposomal Clarithromycin and Its Effect on Pseudomonas Aeruginosa Virulence Factors. Antimicrob. Agents Chemother. 2013, 57, 2694–2704. [Google Scholar] [CrossRef] [Green Version]
- Solleti, V.S.; Alhariri, M.; Halwani, M.; Omri, A. Antimicrobial Properties of Liposomal Azithromycin for Pseudomonas Infections in Cystic Fibrosis Patients. J. Antimicrob. Chemother. 2015, 70, 784–796. [Google Scholar] [CrossRef] [Green Version]
- Mugabe, C.; Azghani, A.O.; Omri, A. Preparation and Characterization of Dehydration–Rehydration Vesicles Loaded with Aminoglycoside and Macrolide Antibiotics. Int. J. Pharm. 2006, 307, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Halwani, M.; Yebio, B.; Suntres, Z.E.; Alipour, M.; Azghani, A.O.; Omri, A. Co-Encapsulation of Gallium with Gentamicin in Liposomes Enhances Antimicrobial Activity of Gentamicin against Pseudomonas Aeruginosa. J. Antimicrob. Chemother. 2008, 62, 1291–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innovotech MBEC Assay®. Available online: https://www.innovotech.ca/products/mbec-assays/ (accessed on 17 April 2021).
- Alipour, M.; Omri, A.; Suntres, Z.E. Ginseng Aqueous Extract Attenuates the Production of Virulence Factors, Stimulates Twitching and Adhesion, and Eradicates Biofilms of Pseudomonas Aeruginosa. Can. J. Physiol. Pharmacol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M.; Suntres, Z.E.; Lafrenie, R.M.; Omri, A. Attenuation of Pseudomonas Aeruginosa Virulence Factors and Biofilms by Co-Encapsulation of Bismuth–Ethanedithiol with Tobramycin in Liposomes. J. Antimicrob. Chemother. 2010, 65, 684–693. [Google Scholar] [CrossRef]
- Vasavi, H.S.; Sudeep, H.V.; Lingaraju, H.B.; Shyam Prasad, K. Bioavailability-Enhanced ResveramaxTM Modulates Quorum Sensing and Inhibits Biofilm Formation in Pseudomonas Aeruginosa PAO1. Microb. Pathog. 2017, 104, 64–71. [Google Scholar] [CrossRef]
- El-Mowafy, S.A.; Abd El Galil, K.H.; El-Messery, S.M.; Shaaban, M.I. Aspirin Is an Efficient Inhibitor of Quorum Sensing, Virulence and Toxins in Pseudomonas Aeruginosa. Microb. Pathog. 2014, 74, 25–32. [Google Scholar] [CrossRef]
- Miller, J.H.; Miller, J.B. Experiments in Molecular Genetics; Bacterial genetics-E. coli; Cold Spring Harbor Laboratory: New York, NY, USA, 1972; ISBN 978-0-87969-106-6. [Google Scholar]
- Mugabe, C.; Halwani, M.; Azghani, A.O.; Lafrenie, R.M.; Omri, A. Mechanism of Enhanced Activity of Liposome-Entrapped Aminoglycosides against Resistant Strains of Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 2016–2022. [Google Scholar] [CrossRef] [Green Version]
- Alhariri, M.; Majrashi, M.A.; Bahkali, A.H.; Almajed, F.S.; Azghani, A.O.; Khiyami, M.A.; Alyamani, E.J.; Aljohani, S.M.; Halwani, M.A. Efficacy of Neutral and Negatively Charged Liposome-Loaded Gentamicin on Planktonic Bacteria and Biofilm Communities. Int. J. Nanomed. 2017, 12, 6949–6961. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Luo, L.; Liang, S.; Long, M.; Xu, H. Influence of Probe-Sonication Process on Drug Entrapment Efficiency of Liposomes Loaded with a Hydrophobic Drug. Int. J. Polym. Mater. 2019, 68, 193–197. [Google Scholar] [CrossRef]
- Jia, Y.; Joly, H.; Omri, A. Characterization of the Interaction between Liposomal Formulations and Pseudomonas Aeruginosa. J. Liposome Res. 2010, 20, 134–146. [Google Scholar] [CrossRef]
- Ye, T.; Sun, S.; Sugianto, T.D.; Tang, P.; Parumasivam, T.; Chang, Y.K.; Astudillo, A.; Wang, S.; Chan, H.-K. Novel Combination Proliposomes Containing Tobramycin and Clarithromycin Effective against Pseudomonas Aeruginosa Biofilms. Int. J. Pharm. 2018, 552, 130–138. [Google Scholar] [CrossRef]
- Bandara, H.M.H.N.; Herpin, M.J.; Kolacny, D.; Harb, A.; Romanovicz, D.; Smyth, H.D.C. Incorporation of Farnesol Significantly Increases the Efficacy of Liposomal Ciprofloxacin against Pseudomonas Aeruginosa Biofilms in Vitro. Mol. Pharm. 2016, 13, 2760–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieme, L.; Hartung, A.; Tramm, K.; Klinger-Strobel, M.; Jandt, K.D.; Makarewicz, O.; Pletz, M.W. MBEC versus MBIC: The Lack of Differentiation between Biofilm Reducing and Inhibitory Effects as a Current Problem in Biofilm Methodology. Biol. Proced. Online 2019, 21, 1–5. [Google Scholar] [CrossRef]
- Sandri, A.; Ortombina, A.; Boschi, F.; Cremonini, E.; Boaretti, M.; Sorio, C.; Melotti, P.; Bergamini, G.; Lleo, M. Inhibition of Pseudomonas Aeruginosa Secreted Virulence Factors Reduces Lung Inflammation in CF Mice. Virulence 2018, 9, 1008–1018. [Google Scholar] [CrossRef] [Green Version]
- Burr, L.D.; Rogers, G.B.; Chen, A.C.-H.; Hamilton, B.R.; Pool, G.F.; Taylor, S.L.; Venter, D.; Bowler, S.D.; Biga, S.; McGuckin, M.A. Macrolide Treatment Inhibits Pseudomonas Aeruginosa Quorum Sensing in Non–Cystic Fibrosis Bronchiectasis. An Analysis from the Bronchiectasis and Low-Dose Erythromycin Study Trial. Ann. Am. Thorac. Soc. 2016, 13, 1697–1703. [Google Scholar] [PubMed]
- Khan, F.; Lee, J.-W.; Pham, D.T.N.; Lee, J.-H.; Kim, H.-W.; Kim, Y.-K.; Kim, Y.-M. Streptomycin Mediated Biofilm Inhibition and Suppression of Virulence Properties in Pseudomonas Aeruginosa PAO1. Appl. Microbiol. Biotechnol. 2020, 104, 799–816. [Google Scholar] [CrossRef]
- Rampioni, G.; Pillai, C.R.; Longo, F.; Bondì, R.; Baldelli, V.; Messina, M.; Imperi, F.; Visca, P.; Leoni, L. Effect of Efflux Pump Inhibition on Pseudomonas Aeruginosa Transcriptome and Virulence. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Gupta, P.; Chhibber, S.; Harjai, K. Subinhibitory Concentration of Ciprofloxacin Targets Quorum Sensing System of Pseudomonas Aeruginosa Causing Inhibition of Biofilm Formation & Reduction of Virulence. Indian J. Med. Res. 2016, 143, 643–651. [Google Scholar] [CrossRef]
- Kiymaci, M.E.; Altanlar, N.; Gumustas, M.; Ozkan, S.A.; Akin, A. Quorum Sensing Signals and Related Virulence Inhibition of Pseudomonas Aeruginosa by a Potential Probiotic Strain’s Organic Acid. Microb. Pathog. 2018, 121, 190–197. [Google Scholar] [CrossRef]
- El-Mowafy, S.A.; Galil, K.H.A.E.; Habib, E.-S.E.; Shaaban, M.I. Quorum Sensing Inhibitory Activity of Sub-Inhibitory Concentrations of β-Lactams. Afr. Health Sci. 2017, 17, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Nalca, Y.; Jänsch, L.; Bredenbruch, F.; Geffers, R.; Buer, J.; Häussler, S. Quorum-Sensing Antagonistic Activities of Azithromycin in Pseudomonas Aeruginosa PAO1: A Global Approach. Antimicrob. Agents Chemother. 2006, 50, 1680–1688. [Google Scholar] [CrossRef] [Green Version]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Cámara, M.; Martínez, J.L. Role of the Multidrug Resistance Efflux Pump MexCD-OprJ in the Pseudomonas Aeruginosa Quorum Sensing Response. Front. Microbiol. 2018, 9, 2757. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Déziel, E. MexEF-OprN Efflux Pump Exports the Pseudomonas Quinolone Signal (PQS) Precursor HHQ (4-Hydroxy-2-Heptylquinoline). PLoS ONE 2011, 6, e24310. [Google Scholar] [CrossRef] [PubMed]
- Aendekerk, S.; Diggle, S.P.; Song, Z.; Høiby, N.; Cornelis, P.; Williams, P.; Cámara, M. The MexGHI-OpmD Multidrug Efflux Pump Controls Growth, Antibiotic Susceptibility and Virulence in Pseudomonas Aeruginosa via 4-Quinolone-Dependent Cell-to-Cell Communication. Microbiology 2005, 151, 1113–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, A.; Zhang, M.; Du, W.; Wang, D.; Ma, L.Z. A Molecular Mechanism for How Sigma Factor AlgT and Transcriptional Regulator AmrZ Inhibit Twitching Motility in Pseudomonas Aeruginosa. Environ. Microbiol. 2020, 23, 572–587. [Google Scholar] [CrossRef] [PubMed]
- Dave, A.; Samarth, A.; Karolia, R.; Sharma, S.; Karunakaran, E.; Partridge, L.; MacNeil, S.; Monk, P.N.; Garg, P.; Roy, S. Characterization of Ocular Clinical Isolates of Pseudomonas Aeruginosa from Non-Contact Lens Related Keratitis Patients from South India. Microorganisms 2020, 8, 260. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, D.; Harikrishnan, A.; Jyoti, K.; Ali, M.I.; Jeevaratnam, K. A Compound Isolated from Alpinia Officinarum Hance. Inhibits Swarming Motility of Pseudomonas Aeruginosa and down Regulates Virulence Genes. J. Appl. Microbiol. 2020, 128, 1355–1365. [Google Scholar] [CrossRef]
- Molinari, G.; Paglia, P.; Schito, G.C. Inhibition of Motility of Pseudomonas Aeruginosa and Proteus Mirabilis by Subinhibitory Concentrations of Azithromycin. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 469–471. [Google Scholar] [CrossRef]
- Bahari, S.; Zeighami, H.; Mirshahabi, H.; Roudashti, S.; Haghi, F. Inhibition of Pseudomonas Aeruginosa Quorum Sensing by Subinhibitory Concentrations of Curcumin with Gentamicin and Azithromycin. J. Glob. Antimicrob. Resist. 2017, 10, 21–28. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Efflux Pumps | Substrates | QS Systems (Molecules) | Function | Virulence Factors | Function |
|---|---|---|---|---|---|
| MexAB-OprM | Beta-lactams, macrolides, tetracyclines [20], and 3-oxo-dodecanoyl homoserine lactone (3OC12-HSL, QS signal) [21] | LasI/R (3OC12-HSL) | Regulates elastase, protease, exotoxin A, biofilm formation and induces PQS and Rhl systems [34,35] | Protease | Immune invasion and host tissue damage [7,29] |
| MexXY-OprM | Aminoglycosides, fluoroquinolones, tetracyclines and macrolides [22,23,24] | RhlI/R (C4-HSL) | Regulates the production of pyocyanin, rhamnolipids, elastase and hydrogen cyanide [36] | Pyocyanin | Induces oxidative stress, neutrophil apoptosis, inhibits ciliary beating in the airways, and causes cytotoxicity [37,38] |
| MexCD-OprJ | Macrolides, cephalosporins, fluoroquinolones, tetracyclines and organic solvents [39] | PQS 1 (PQS signal) | Regulates the expression of pyoverdine, pyocyanin, rhamnolipids and the RhlI/R system [35] | Pyoverdine | Iron scavenging, vital for pathogenesis. Sequestrates iron from host, which is used for biofilm formation [40] |
| MexEF-OprN | Chloramphenicol, tetracycline, fluoroquinolones, HHQ 2 (QS signal) [39] | Lipase | Degrades lipids in the host [29] |
| Liposomal Antibiotics | Size (nm) | Polydispersity Index | Encapsulation Efficiency (%) | Concentration (mg/mL) |
|---|---|---|---|---|
| Gentamicin | 562.67 ± 33.74 | 0.7 ± 0.11 | 13.89 ± 1.545 | 0.42 ± 0.046 |
| Erythromycin | 3086.35 ± 553.95 | 0.6 ± 0.12 | 51.58 ± 2.846 | 10.32 ± 0.571 |
| Strains | MIC (mg/L) (MBC *(mg/L)) | MIC (mg/L) (MBC * (mg/L)) | ||||||
|---|---|---|---|---|---|---|---|---|
| Free GEN | Lipo GEN | Free ERY | Lipo ERY | |||||
| −PABN | +PABN | −PABN | +PABN | −PABN | +PABN | −PABN | +PABN | |
| PA 01 | 8 (32) | 8 (16) | 2 (4) | 1 (2) | 512 (512) | 256 (256) | 128 (128) | 128 (128) |
| PA 11 | 256 (1024) | 32 (128) | 32 (128) | 8 (32) | 1024 (1024) | 512 (512) | 128 (256) | 64(256) |
| Strains | MBEC (mg/L) | |||||||
|---|---|---|---|---|---|---|---|---|
| Free GEN | Lipo GEN | Free ERY | Lipo ERY | |||||
| −PABN | +PABN | −PABN | +PABN | −PABN | +PABN | −PABN | +PABN | |
| PA 01 | 64 | 16 | 4 | 2 | 1024 | 1024 | 128 | 64 |
| PA 11 | 1024 | 128 | 256 | 128 | 1024 | 512 | 512 | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gbian, D.L.; Omri, A. The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas aeruginosa. Pharmaceutics 2021, 13, 577. https://doi.org/10.3390/pharmaceutics13040577
Gbian DL, Omri A. The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas aeruginosa. Pharmaceutics. 2021; 13(4):577. https://doi.org/10.3390/pharmaceutics13040577
Chicago/Turabian StyleGbian, Douweh Leyla, and Abdelwahab Omri. 2021. "The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas aeruginosa" Pharmaceutics 13, no. 4: 577. https://doi.org/10.3390/pharmaceutics13040577