
Identification of Bioactive Natural Product from the Stems and Stem Barks of Cornus walteri: Benzyl Salicylate Shows Potential Anti-Inflammatory Activity in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Extraction and Isolation of Compounds 1–15
2.2. RAW 264.7 Cells Culture
2.3. Measurement of Viability of RAW 264.7 Cells
2.4. Measurement of NO Produced by RAW 264.7 Cells
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
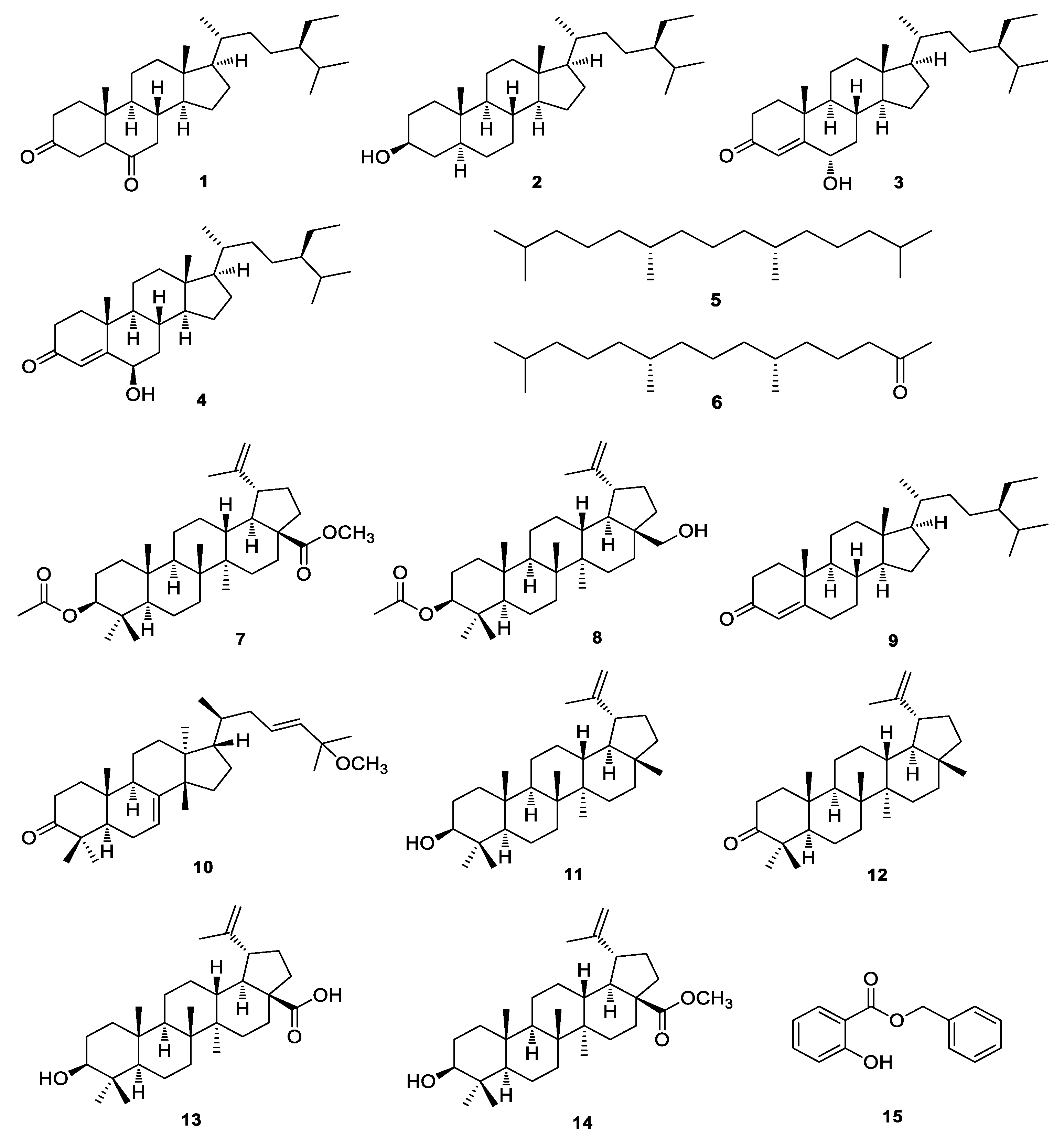
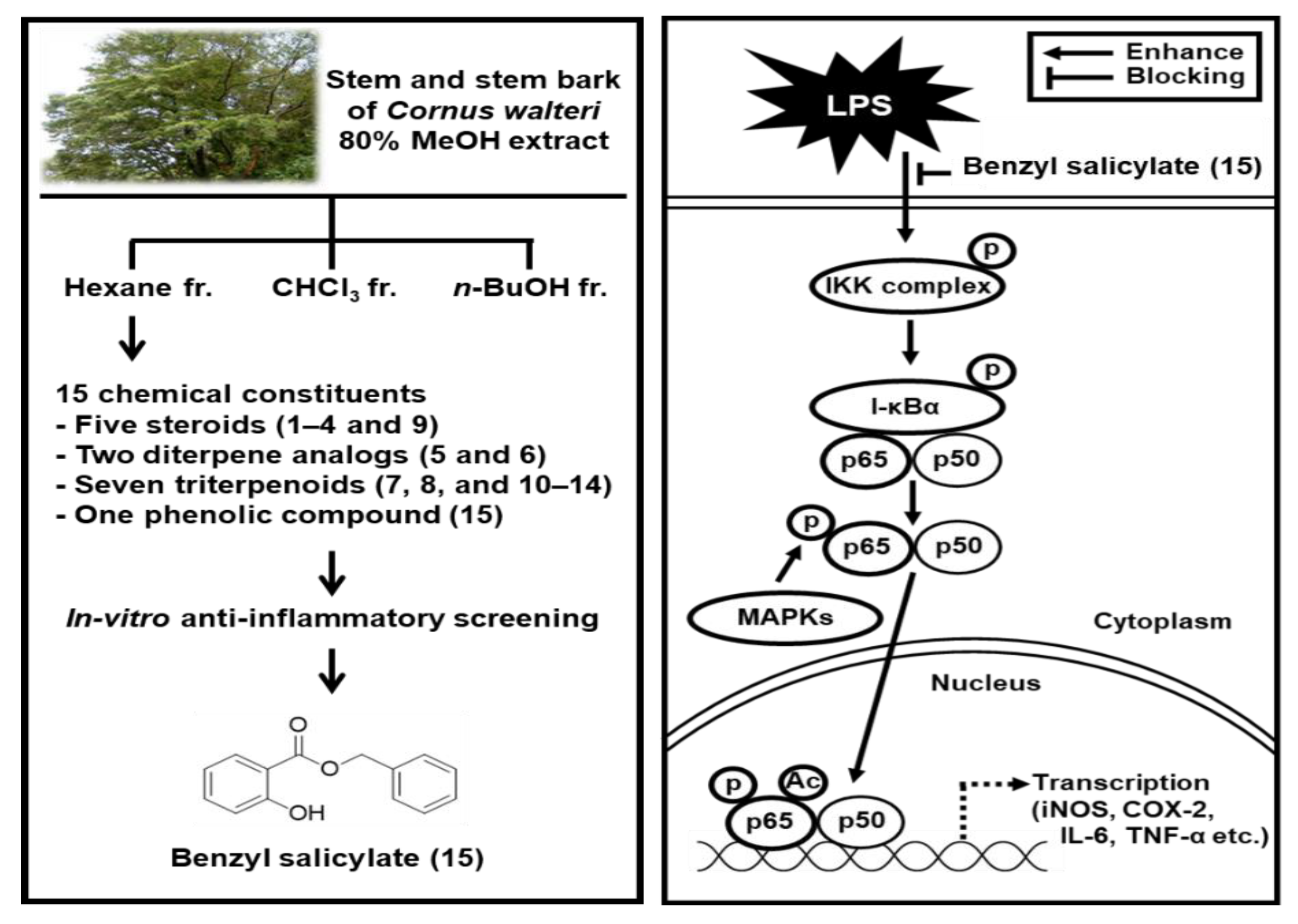
3.1. Isolation and Identification of the Compounds
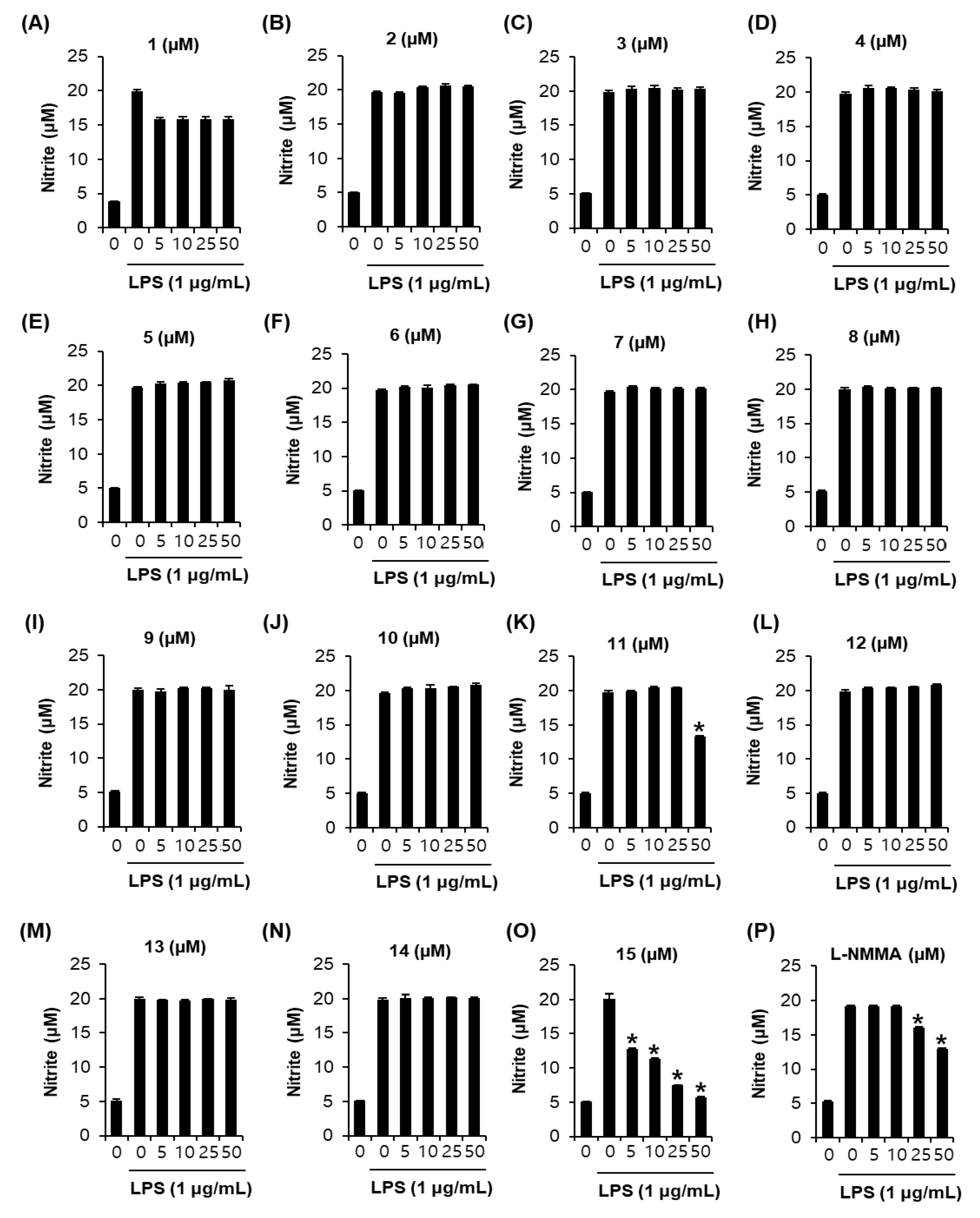
3.2. Effects of Compounds 1–15 on Nitric Oxide (NO) Production
3.3. Effects of Benzyl Salicylate (15) on the LPS-Induced Expression of IKKα/β, I-κBα, and NF-κB in RAW 264.7 Mouse Macrophages
3.4. Effects of Benzyl Salicylate (15) on the LPS-Induced Expression of iNOS and COX-2 in RAW 264.7 Mouse Macrophages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Muniandy, K.; Gothai, S.; Badran, K.M.H.; Kumar, S.S.; Esa, N.M.; Arulselvan, P. Suppression of Proinflammatory Cytokines and Mediators in LPS-Induced RAW 264.7 Macrophages by Stem extract of Alternanthera sessilis via the Inhibition of the NF-κB Pathway. J. Immunol. Res. 2018, 2018, 3430684. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Ryoo, R.; Kim, J.C.; Park, H.B.; Kang, K.S.; Kim, K.H. Calvatianone, a Sterol Possessing a 6/5/6/5-Fused Ring System with a Contracted Tetrahydrofuran B-Ring, from the Fruiting Bodies of Calvatia nipponica. J. Nat. Prod. 2020, 83, 2737–2742. [Google Scholar] [CrossRef]
- Lee, S.R.; Kang, H.S.; Yoo, M.J.; Yi, S.A.; Beemelmanns, C.; Lee, J.C.; Kim, K.H. Anti-adipogenic Pregnane Steroid from a Hydractinia-associated Fungus, Cladosporium sphaerospermum SW67. Nat. Prod. Sci. 2020, 26, 230–235. [Google Scholar]
- Lee, S.; Ryoo, R.; Choi, J.H.; Kim, J.H.; Kim, S.H.; Kim, K.H. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity. Arch. Pharm. Res. 2020, 43, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.A.; Park, E.J.; Lee, D.; Song, J.H.; Lee, H.L.; Kim, K.H.; Kim, Y.; Jung, K.; Kang, K.S.; Yoo, J.E. Estrogenic Activity of Sanguiin H-6 through Activation of Estrogen Receptor α Coactivator-binding Site. Nat. Prod. Sci. 2019, 25, 28–33. [Google Scholar] [CrossRef]
- Ha, J.W.; Kim, J.; Kim, H.; Jang, W.; Kim, K.H. Mushrooms: An Important Source of Natural Bioactive Compounds. Nat. Prod. Sci. 2020, 26, 118–131. [Google Scholar]
- Yu, J.S.; Li, C.; Kwon, M.; Oh, T.; Lee, T.H.; Kim, D.H.; Ahn, J.S.; Ko, S.K.; Kim, C.S.; Cao, S.; et al. Herqueilenone A, a unique rearranged benzoquinone-chromanone from the hawaiian volcanic soil-associated fungal strain Penicillium herquei FT729. Bioorg. Chem. 2020, 105, 104397. [Google Scholar] [CrossRef]
- Yu, J.S.; Park, M.; Pang, C.; Rashan, L.; Jung, W.H.; Kim, K.H. Antifungal Phenols from Woodfordia uniflora Collected in Oman. J. Nat. Prod. 2020, 83, 2261–2268. [Google Scholar] [CrossRef] [PubMed]
- Park, H.C.; Jung, T.K.; Kim, M.J.; Yoon, K.S. Protective effect of Cornus walteri Wangerin leaf against UVB irradiation induced photoaging in human reconstituted skin. J. Ethnopharmacol. 2016, 193, 445–449. [Google Scholar] [CrossRef]
- Choi, W.H.; Park, W.Y.; Hwang, B.Y.; Oh, G.J.; Kang, S.J.; Lee, K.S.; Ro, J.S. Phenolic Compounds from the Stem Bark of Cornus walteri Wanger. Korean J. Pharmacogn. 1998, 29, 217–224. [Google Scholar]
- Lee, S.H.; Yoon, K.R.; Lee, E.; Cha, Y.Y. Anti-Inflammatory Effect of Cornus walteri. J. Physiol. Pathol. Korean Med. 2011, 25, 982–988. [Google Scholar]
- Park, W.H.; Cha, Y.Y. Effects of Stem Bark Extracts of Cornus walteri Wanger on the Lipid Lowering, Anti-oxidative Activity and Concentration of Proinflammatory Cytokines in Rat Fed High Fat Diet. J. Korean Med. Rehabi. 2009, 19, 59–78. [Google Scholar]
- Lee, D.Y.; Yoo, K.H.; Chung, I.S.; Kim, J.Y.; Chung, D.K.; Kim, D.K.; Kim, S.H.; Baek, N.I. A new lignan glycoside from the fruits of Cornus kousa Burg. Arch. Pharm. Res. 2008, 31, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Vareed, S.K.; Schutzki, R.E.; Nair, M.G. Lipid peroxidation, cyclooxygenase enzyme and tumor cell proliferation inhibitory compounds in Cornus kousa fruits. Phytomedicine 2007, 14, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Choi, S.U.; Kim, Y.C.; Lee, K.R. Tirucallane Triterpenoids from Cornus walteri. J. Nat. Prod. 2011, 74, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Shin, Y.J.; Choi, S.U.; Lee, K.R. New Cytotoxic δ-Valerolactones from Cornus walteri. Bull. Korean Chem. Soc. 2011, 32, 2443–2445. [Google Scholar] [CrossRef]
- Lee, S.R.; Nam, J.W.; Kim, K.H. New Triterpenoids from the stems of Cornus walteri. Chem. Pharm. Bull. 2017, 65, 683–686. [Google Scholar] [CrossRef]
- Lee, D.; Lee, S.R.; Kang, K.S.; Ko, Y.; Pang, C.; Yamabe, N.; Kim, K.H. Betulinic Acid Suppresses Ovarian Cancer Cell Proliferation through Induction of Apoptosis. Biomolecules 2019, 9, 257. [Google Scholar] [CrossRef]
- Lee, S.R.; Choi, E.; Jeon, S.H.; Zhi, X.Y.; Yu, J.S.; Kim, S.H.; Lee, J.; Park, K.M.; Kim, K.H. Tirucallane Triterpenoids from the Stems and Stem Bark of Cornus walteri that Control Adipocyte and Osteoblast Differentiations. Molecules 2018, 23, 2732. [Google Scholar] [CrossRef]
- Lee, D.; Lee, S.R.; Kang, K.S.; Kim, K.H. Benzyl salicylate from the stems and stem barks of Cornus walteri as a nephroprotective agent against cisplatin-induced apoptotic cell death in LLC-PK1 cells. RSC Adv. 2020, 10, 5777–5784. [Google Scholar] [CrossRef]
- Yang, E.J.; Yim, E.Y.; Song, G.; Kim, G.O.; Hyun, C.G. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Interdiscipl. Toxicol. 2009, 2, 245–249. [Google Scholar] [CrossRef]
- Zhang, D.D.; Yang, J.; Luo, J.F.; Li, X.N.; Long, C.L.; Wang, Y.H. New aporphine alkaloids from the aerial parts of Piper semiimmersum. J. Asian Nat. Prod. Res. 2018, 20, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Hattori, T.; Hamaguchi, N.; Masuda, K.; Takano, A.; Shiojima, K. Fern Constituents: Dryocrassyl Formate, Sitostanyl Formate and 12α-Hydroxyfern-9(11)-ene from Cyathea podophylla. Chem. Pharm. Bull. 2003, 51, 1311–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.K.; Zheng, C.J.; Li, X.B.; Chen, G.Y.; Han, C.R.; Chen, W.H.; Song, X.P. Two New Lanostane Triterpenoids from the Branches and Leaves of Polyalthia obliqua. Molecules 2014, 19, 7621–7628. [Google Scholar] [CrossRef]
- Della Greca, M.; Monaco, P.; Previtera, L. Stigmasterols from Typha latifolia. J. Nat. Prod. 1990, 53, 1430–1435. [Google Scholar] [CrossRef]
- Dalling, D.K.; Pugmire, R.J.; Grant, D.M.; Hull, W.E. The use of high-field carbon-13 NMR spectroscopy to characterize chiral centers in isopranes. Magn. Reson. Chem. 1986, 24, 191–198. [Google Scholar] [CrossRef]
- Zhao, L.; Jin, C.; Mao, Z.; Gopinathan, M.B.; Rehder, K.; Brinton, R.D. Design, Synthesis, and Estrogenic Activity of a Novel Estrogen Receptor Modulator-A Hybrid Structure of 17β-Estradiol and Vitamin E in Hippocampal Neurons. J. Med. Chem. 2007, 50, 4471–4481. [Google Scholar] [CrossRef]
- Urban, M.; Sarek, J.; Klinot, J.; Hajduch, M. Synthesis of A-Seco Derivatives of Betulinic Acid with Cytotoxic Activity. J. Nat. Prod. 2004, 67, 1100–1105. [Google Scholar] [CrossRef]
- Santos, R.C.; Salvador, J.A.R.; Marin, S.; Cascante, M. Novel semisynthetic derivatives of betulin and betulinic acid with cytotoxic activity. Bioorg. Med. Chem. 2009, 17, 6241–6250. [Google Scholar] [CrossRef]
- Prachayasittikul, S.; Suphapong, S.; Worachartcheewan, A.; Lawung, R.; Ruchirawat, S.; Prachayasittikul, V. Bioactive Metabolites from Spilanthes acmella Murr. Molecules 2009, 14, 850–867. [Google Scholar] [CrossRef]
- Fotie, J.; Bohle, D.S.; Leimanis, M.L.; Georges, E.; Rukunga, G.; Nkengfack, A.E. Lupeol Long-Chain Fatty Acid Esters with Antimalarial Activity from Holarrhena floribunda. J. Nat. Prod. 2006, 69, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Puapairoj, P.; Naengchomnong, W.; Kijjoa, A.; Pinto, M.M.; Pedro, M.; Nascimento, M.S.J.; Silva, A.M.S.; Herz, W. Cytotoxic Activity of Lupane-Type Triterpenes from Glochidion sphaerogynum and Glochidion eriocarpum Two of which Induce Apoptosis. Planta Med. 2005, 71, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Sholichin, M.; Yamasaki, K.; Kasai, R.; Tanaka, O. Carbon-13 Nuclear Magnetic Resonance of Lupane-Type Triterpenes, Lupeol, Betulin and Betulinic Acid. Chem. Pharm. Bull. 1980, 28, 1006–1008. [Google Scholar] [CrossRef]
- Pohjala, L.; Alakurtti, S.; Ahola, T.; Yli-Kauhaluoma, J.; Tammela, P. Betulin-Derived Compounds as Inhibitors of Alphavirus Replication. J. Nat. Prod. 2009, 72, 1917–1926. [Google Scholar] [CrossRef]
- Talzi, V.P. A 13C and 1H NMR analysis of perfumes. Russ. J. Appl. Chem. 2006, 79, 107–116. [Google Scholar] [CrossRef]
- Li, F.; Cao, Y.; Luo, Y.; Liu, T.; Yan, G.; Chen, L.; Ji, L.; Wang, L.; Chen, B.; Yaseen, A.; et al. Two new triterpenoid saponins derived from the leaves of Panax ginseng and their antiinflammatory activity. J. Ginseng Res. 2019, 43, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Saba, E.; Lee, Y.Y.; Kim, M.; Hyun, S.H.; Park, C.K.; Son, E.; Kim, D.S.; Kim, S.D.; Rhee, M.H. A novel herbal formulation consisting of red ginseng extract and Epimedium koreanum Nakai-attenuated dextran sulfate sodium-induced colitis in mice. J. Ginseng Res. 2020, 44, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Xin, C.; Quan, H.; Kim, J.M.; Hur, Y.H.; Shin, J.Y.; Bae, H.B.; Choi, J.I. Ginsenoside Rb1 increases macrophage phagocytosis through p38 mitogen-activated protein kinase/Akt pathway. J. Ginseng Res. 2019, 43, 394–401. [Google Scholar] [CrossRef]
- Li, J.; Wang, R.F.; Zhou, Y.; Hu, H.J.; Yang, Y.B.; Yang, L.; Wang, Z.T. Dammarane-type triterpene oligoglycosides from the leaves and stems of Panax notoginseng and their antiinflammatory activities. J. Ginseng Res. 2019, 43, 377–384. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Jo, M.S.; Lee, S.; Yu, J.S.; Baek, S.C.; Cho, Y.C.; Kim, K.H. Megastigmane Derivatives from the Cladodes of Opuntia humifusa and Their Nitric Oxide Inhibitory Activities in Macrophages. J. Nat. Prod. 2020, 83, 684–692. [Google Scholar] [CrossRef]
- Charles, A.K.; Darbre, P.D. Oestrogenic activity of benzyl salicylate, benzyl benzoate and butylphenylmethylpropional (Lilial) in MCF7 human breast cancer cells in vitro. J. Appl. Toxicol. 2009, 29, 422–434. [Google Scholar] [CrossRef]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. Regulation of NF-κB Activation by MAP Kinase Cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Liu, Y.; Zhang, X.; Liu, X.; Qi, J. Regulation of iNOS Expression by NF-κB in Human Lens Epithelial Cells Treated with High Levels of Glucose. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5070–5077. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Rahman, M.A.; Dhar, D.K.; Yamaguchi, E.; Maruyama, S.; Sato, T.; Hayashi, H.; Ono, T.; Yamanoi, A.; Kohno, H.; Nagasue, N. Coexpression of Inducible Nitric Oxide Synthase and COX-2 in Hepatocellular Carcinoma and Surrounding Liver: Possible Involvement of COX-2 in the Angiogenesis of Hepatitis C Virus-positive Cases. Clin. Cancer Res. 2001, 7, 1325–1332. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Alishir, A.; Jang, T.S.; Kim, K.H. Identification of Bioactive Natural Product from the Stems and Stem Barks of Cornus walteri: Benzyl Salicylate Shows Potential Anti-Inflammatory Activity in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Pharmaceutics 2021, 13, 443. https://doi.org/10.3390/pharmaceutics13040443
Lee D, Alishir A, Jang TS, Kim KH. Identification of Bioactive Natural Product from the Stems and Stem Barks of Cornus walteri: Benzyl Salicylate Shows Potential Anti-Inflammatory Activity in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Pharmaceutics. 2021; 13(4):443. https://doi.org/10.3390/pharmaceutics13040443
Chicago/Turabian StyleLee, Dahae, Akida Alishir, Tae Su Jang, and Ki Hyun Kim. 2021. "Identification of Bioactive Natural Product from the Stems and Stem Barks of Cornus walteri: Benzyl Salicylate Shows Potential Anti-Inflammatory Activity in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages" Pharmaceutics 13, no. 4: 443. https://doi.org/10.3390/pharmaceutics13040443