
Sulfated Lactosyl Archaeol Archaeosomes Synergize with Poly(I:C) to Enhance the Immunogenicity and Efficacy of a Synthetic Long Peptide-Based Vaccine in a Melanoma Tumor Model

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Vaccine Preparation and Immunization
2.2. Therapeutic Tumor Challenge Model
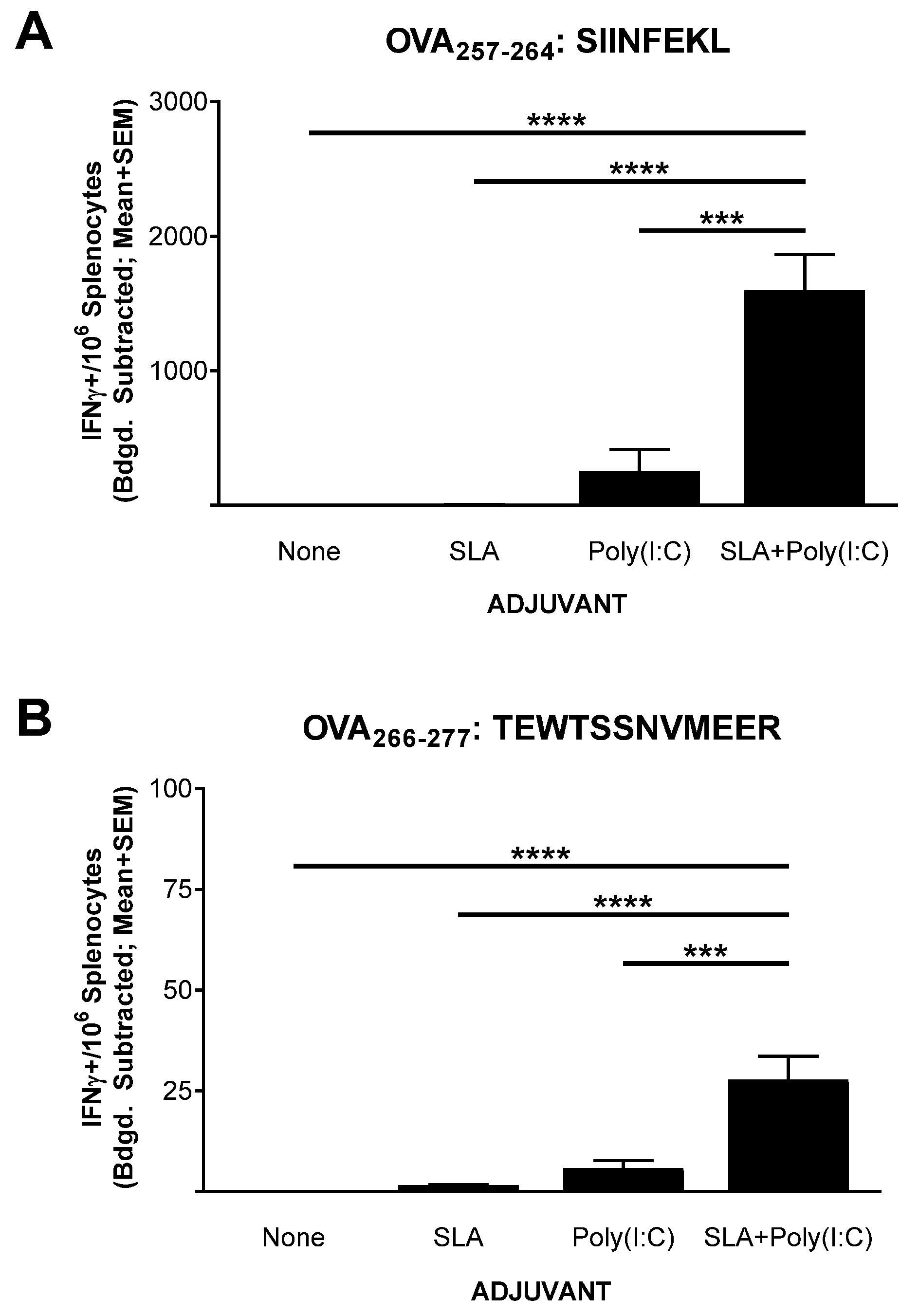
2.3. ELISpot
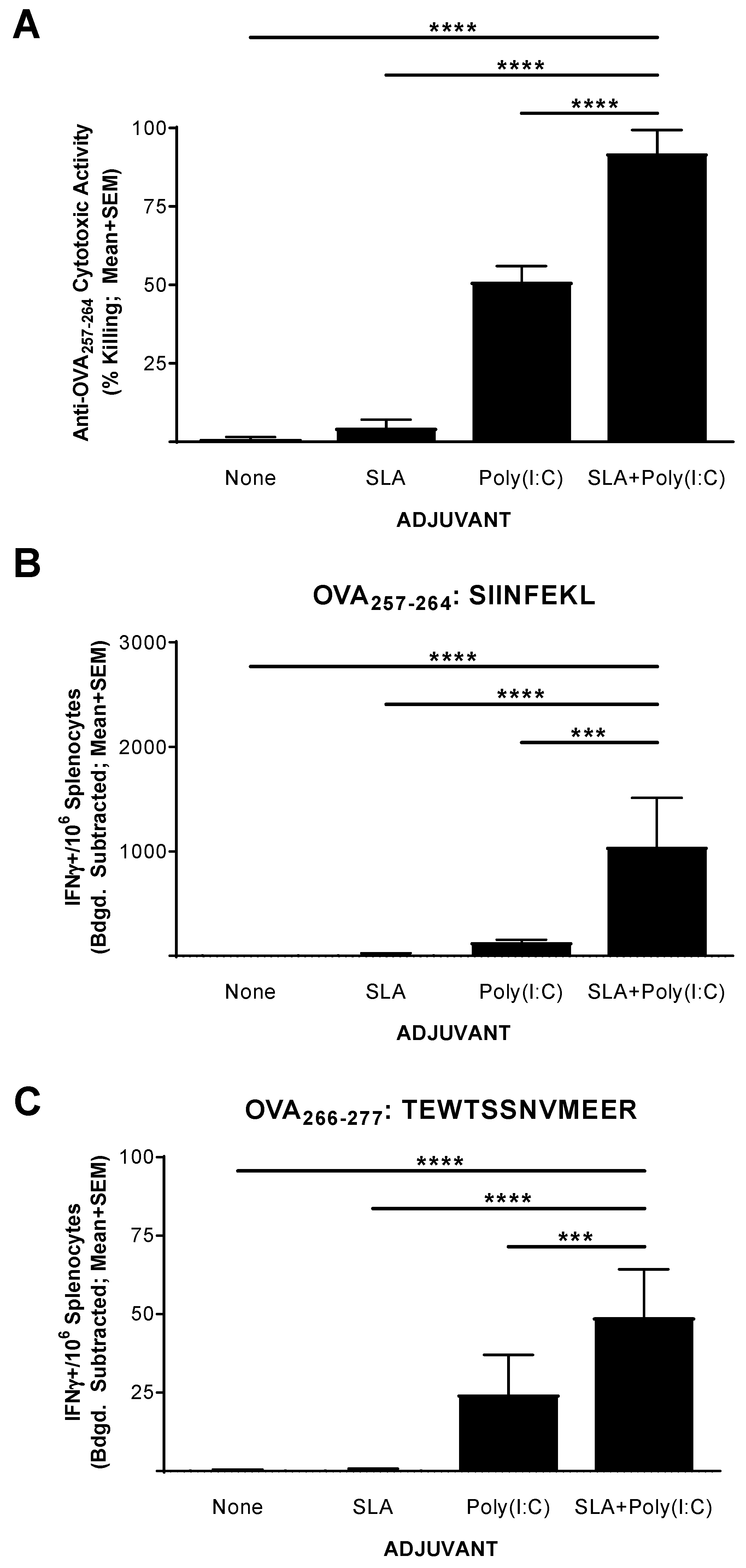
2.4. In Vivo Cytolytic Activity
2.5. Statistical Analysis
3. Results and Discussion
3.1. IFN-γ+ T Cell Response to OVA SLP Vaccine Formulations in Mice
3.2. Functionality of CD8+ T Cell Response to OVA SLP Vaccine Formulations in Mice
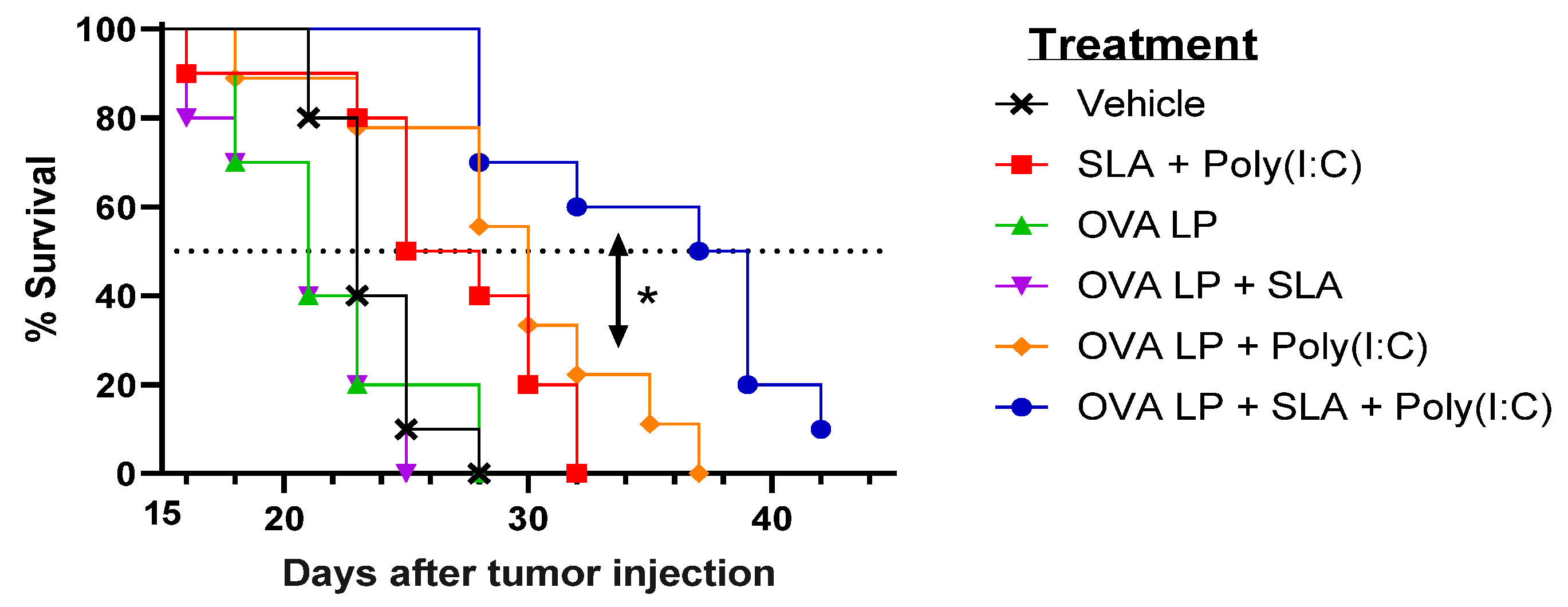
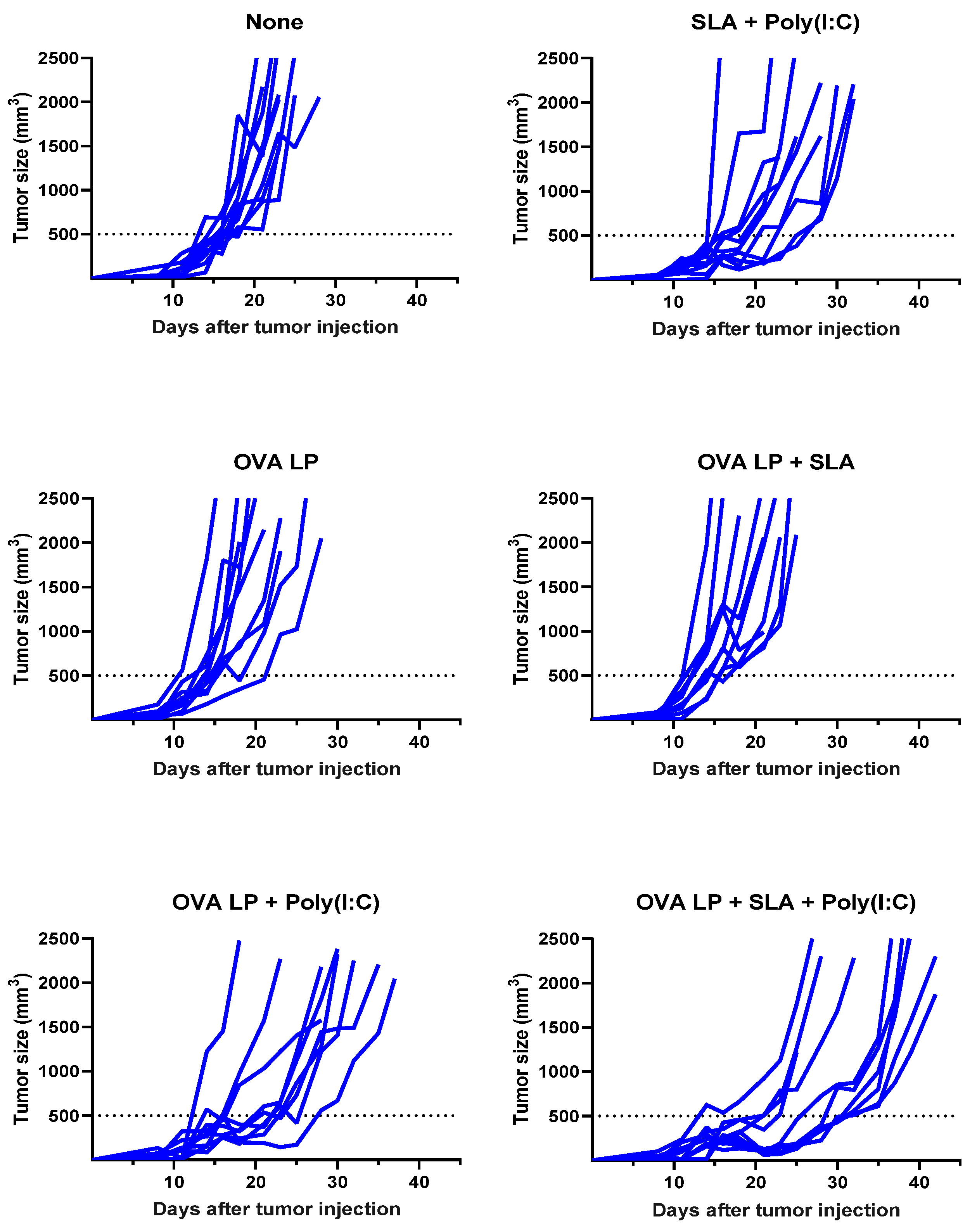
3.3. Anti-Tumor Activity of OVA SLP Vaccine Formulations in a Therapeutic B16-OVA Tumor Challenge Model
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaddepally, R.K.; Kharel, P.; Pandey, R.; Garje, R.; Chandra, A.B. Review of Indications of FDA-Approved Immune Checkpoint Inhibitors per NCCN Guidelines with the Level of Evidence. Cancers 2020, 12, 738. [Google Scholar] [CrossRef] [Green Version]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune Checkpoint Inhibitors: Recent Progress and Potential Biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; Türeci, Ö. Personalized Vaccines for Cancer Immunotherapy. Science 2018, 359, 1355–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumai, T.; Kobayashi, H.; Harabuchi, Y.; Celis, E. Peptide Vaccines in Cancer-Old Concept Revisited. Curr. Opin. Immunol. 2017, 45, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezu, L.; Kepp, O.; Cerrato, G.; Pol, J.; Fucikova, J.; Spisek, R.; Zitvogel, L.; Kroemer, G.; Galluzzi, L. Trial Watch: Peptide-Based Vaccines in Anticancer Therapy. Oncoimmunology 2018, 7. [Google Scholar] [CrossRef]
- Bijker, M.S.; Melief, C.J.M.; Offringa, R.; van der Burg, S.H. Design and Development of Synthetic Peptide Vaccines: Past, Present and Future. Expert Rev. Vaccines 2007, 6, 591–603. [Google Scholar] [CrossRef]
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An Immunogenic Personal Neoantigen Vaccine for Patients with Melanoma. Nature 2017, 547, 217–221. [Google Scholar] [CrossRef]
- Hu, Z.; Ott, P.A.; Wu, C.J. Towards Personalized, Tumour-Specific, Therapeutic Vaccines for Cancer. Nat. Rev. Immunol. 2018, 18, 168–182. [Google Scholar] [CrossRef]
- Jou, J.; Harrington, K.J.; Zocca, M.-B.; Ehrnrooth, E.; Cohen, E.E.W. The Changing Landscape of Therapeutic Cancer Vaccines - Novel Platforms and Neoantigen Identification. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020. [Google Scholar] [CrossRef]
- Slingluff, C.L. The Present and Future of Peptide Vaccines for Cancer: Single or Multiple, Long or Short, Alone or in Combination? Cancer J. Sudbury Mass 2011, 17, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pol, J.; Bloy, N.; Buqué, A.; Eggermont, A.; Cremer, I.; Sautès-Fridman, C.; Galon, J.; Tartour, E.; Zitvogel, L.; Kroemer, G.; et al. Trial Watch: Peptide-Based Anticancer Vaccines. Oncoimmunology 2015, 4, e974411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, K.; Jia, Y.; Krishnan, L. Archaeal Lipid Vaccine Adjuvants for Induction of Cell-Mediated Immunity. Expert Rev. Vaccines 2016, 15, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Akache, B.; Deschatelets, L.; Qian, H.; Dudani, R.; Harrison, B.A.; Stark, F.C.; Chandan, V.; Jamshidi, M.P.; Krishnan, L.; et al. A Comparison of the Immune Responses Induced by Antigens in Three Different Archaeosome-Based Vaccine Formulations. Int. J. Pharm. 2019, 561, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Akache, B.; Deschatelets, L.; Harrison, B.A.; Dudani, R.; Stark, F.C.; Jia, Y.; Landi, A.; Law, J.L.M.; Logan, M.; Hockman, D.; et al. Effect of Different Adjuvants on the Longevity and Strength of Humoral and Cellular Immune Responses to the HCV Envelope Glycoproteins. Vaccines 2019, 7, 204. [Google Scholar] [CrossRef] [Green Version]
- Stark, F.C.; Akache, B.; Ponce, A.; Dudani, R.; Deschatelets, L.; Jia, Y.; Sauvageau, J.; Williams, D.; Jamshidi, M.P.; Agbayani, G.; et al. Archaeal Glycolipid Adjuvanted Vaccines Induce Strong Influenza-Specific Immune Responses through Direct Immunization in Young and Aged Mice or through Passive Maternal Immunization. Vaccine 2019, 37, 7108–7116. [Google Scholar] [CrossRef]
- McCluskie, M.J.; Deschatelets, L.; Krishnan, L. Sulfated Archaeal Glycolipid Archaeosomes as a Safe and Effective Vaccine Adjuvant for Induction of Cell-Mediated Immunity. Hum. Vaccines Immunother. 2017, 13, 2772–2779. [Google Scholar] [CrossRef]
- Akache, B.; Stark, F.C.; Jia, Y.; Deschatelets, L.; Dudani, R.; Harrison, B.A.; Agbayani, G.; Williams, D.; Jamshidi, M.P.; Krishnan, L.; et al. Sulfated Archaeol Glycolipids: Comparison with Other Immunological Adjuvants in Mice. PLoS ONE 2018, 13, e0208067. [Google Scholar] [CrossRef]
- Agbayani, G.; Jia, Y.; Akache, B.; Chandan, V.; Iqbal, U.; Stark, F.C.; Deschatelets, L.; Lam, E.; Hemraz, U.D.; Régnier, S.; et al. Mechanistic Insight into the Induction of Cellular Immune Responses by Encapsulated and Admixed Archaeosome-Based Vaccine Formulations. Hum. Vaccines Immunother. 2020, 16, 2183–2195. [Google Scholar] [CrossRef]
- Akache, B.; Stark, F.C.; Iqbal, U.; Chen, W.; Jia, Y.; Krishnan, L.; McCluskie, M.J. Safety and Biodistribution of Sulfated Archaeal Glycolipid Archaeosomes as Vaccine Adjuvants. Hum. Vaccines Immunother. 2018, 14, 1746–1759. [Google Scholar] [CrossRef]
- Stark, F.C.; Agbayani, G.; Sandhu, J.K.; Akache, B.; McPherson, C.; Deschatelets, L.; Dudani, R.; Hewitt, M.; Jia, Y.; Krishnan, L.; et al. Simplified Admix Archaeal Glycolipid Adjuvanted Vaccine and Checkpoint Inhibitor Therapy Combination Enhances Protection from Murine Melanoma. Biomedicines 2019, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonam, S.R.; Bhunia, D.; Muller, S.; Nerella, S.G.; Alvala, M.; Halmuthur Mahabalarao, S.K. Novel Trisaccharide Based Phospholipids as Immunomodulators. Int. Immunopharmacol. 2019, 74, 105684. [Google Scholar] [CrossRef]
- Nalla, N.; Pallavi, P.; Reddy, B.S.; Miryala, S.; Naveen Kumar, V.; Mahboob, M.; Halmuthur, M.S.K. Design, Synthesis and Immunological Evaluation of 1,2,3-Triazole-Tethered Carbohydrate-Pam3Cys Conjugates as TLR2 Agonists. Bioorg Med Chem 2015, 23, 5846–5855. [Google Scholar] [CrossRef]
- Kumai, T.; Fan, A.; Harabuchi, Y.; Celis, E. Cancer Immunotherapy: Moving Forward with Peptide T Cell Vaccines. Curr. Opin. Immunol. 2017, 47, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, D.M.; Sprott, G.D.; Krishnan, L. Sulfated-Glycolipids as Adjuvants for Vaccines. Patent No. WO/2016/004512, 14 January 2016. [Google Scholar]
- Akache, B.; McCluskie, M.J. The Quantification of Antigen-Specific T Cells by IFN-γ ELISpot. Methods Mol. Biol. Clifton NJ 2021, 2183, 525–536. [Google Scholar] [CrossRef]
- Stark, F.C.; Dudani, R.; Agbayani, G.; McCluskie, M.J. A Method to Evaluate In Vivo CD8+ T Cell Cytotoxicity in a Murine Model. Methods Mol. Biol. Clifton NJ 2021, 2183, 549–558. [Google Scholar] [CrossRef]
- Conlan, J.W.; Krishnan, L.; Willick, G.E.; Patel, G.B.; Sprott, G.D. Immunization of Mice with Lipopeptide Antigens Encapsulated in Novel Liposomes Prepared from the Polar Lipids of Various Archaeobacteria Elicits Rapid and Prolonged Specific Protective Immunity against Infection with the Facultative Intracellular Pathogen, Listeria Monocytogenes. Vaccine 2001, 19, 3509–3517. [Google Scholar] [CrossRef] [PubMed]
- Levast, B.; Awate, S.; Babiuk, L.; Mutwiri, G.; Gerdts, V.; van Drunen Littel-van den Hurk, S. Vaccine Potentiation by Combination Adjuvants. Vaccines 2014, 2, 297. [Google Scholar] [CrossRef] [Green Version]
- Nanishi, E.; Dowling, D.J.; Levy, O. Toward Precision Adjuvants: Optimizing Science and Safety. Curr. Opin. Pediatr. 2020, 32, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Garçon, N.; Di Pasquale, A. From Discovery to Licensure, the Adjuvant System Story. Hum. Vaccines Immunother. 2017, 13, 19–33. [Google Scholar] [CrossRef]
- Miyake, T.; Kumagai, Y.; Kato, H.; Guo, Z.; Matsushita, K.; Satoh, T.; Kawagoe, T.; Kumar, H.; Jang, M.H.; Kawai, T.; et al. Poly I:C-Induced Activation of NK Cells by CD8 Alpha+ Dendritic Cells via the IPS-1 and TRIF-Dependent Pathways. J. Immunol. Baltim. Md 1950 2009, 183, 2522–2528. [Google Scholar] [CrossRef] [Green Version]
- Speiser, D.E.; Liénard, D.; Rufer, N.; Rubio-Godoy, V.; Rimoldi, D.; Lejeune, F.; Krieg, A.M.; Cerottini, J.-C.; Romero, P. Rapid and Strong Human CD8+ T Cell Responses to Vaccination with Peptide, IFA, and CpG Oligodeoxynucleotide 7909. J. Clin. Investig. 2005, 115, 739–746. [Google Scholar] [CrossRef]
- Karbach, J.; Gnjatic, S.; Bender, A.; Neumann, A.; Weidmann, E.; Yuan, J.; Ferrara, C.A.; Hoffmann, E.; Old, L.J.; Altorki, N.K.; et al. Tumor-Reactive CD8+ T-Cell Responses after Vaccination with NY-ESO-1 Peptide, CpG 7909 and Montanide ISA-51: Association with Survival. Int. J. Cancer 2010, 126, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Fourcade, J.; Kudela, P.; Andrade Filho, P.A.; Janjic, B.; Land, S.R.; Sander, C.; Krieg, A.; Donnenberg, A.; Shen, H.; Kirkwood, J.M.; et al. Immunization with Analog Peptide in Combination with CpG and Montanide Expands Tumor Antigen-Specific CD8+ T Cells in Melanoma Patients. J. Immunother. Hagerstown Md 1997 2008, 31, 781–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailemichael, Y.; Dai, Z.; Jaffarzad, N.; Ye, Y.; Medina, M.A.; Huang, X.-F.; Dorta-Estremera, S.M.; Greeley, N.R.; Nitti, G.; Peng, W.; et al. Persistent Antigen at Vaccination Sites Induces Tumor-Specific CD8+ T Cell Sequestration, Dysfunction and Deletion. Nat. Med. 2013, 19, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Treatment | Median Survival (Days) |
|---|---|
| Vehicle | 23 |
| Sulfated Lactosyl Archaeol (SLA) + Poly (I:C) (No Antigen) | 26.5 |
| Ovalbumin synthetic long peptide (OVA SLP) | 21 |
| OVA SLP + SLA | 21 |
| OVA SLP + Poly (I:C) | 30 |
| OVA SLP + SLA + Poly (I:C) | 38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akache, B.; Agbayani, G.; Stark, F.C.; Jia, Y.; Dudani, R.; Harrison, B.A.; Deschatelets, L.; Chandan, V.; Lam, E.; Hemraz, U.D.; et al. Sulfated Lactosyl Archaeol Archaeosomes Synergize with Poly(I:C) to Enhance the Immunogenicity and Efficacy of a Synthetic Long Peptide-Based Vaccine in a Melanoma Tumor Model. Pharmaceutics 2021, 13, 257. https://doi.org/10.3390/pharmaceutics13020257
Akache B, Agbayani G, Stark FC, Jia Y, Dudani R, Harrison BA, Deschatelets L, Chandan V, Lam E, Hemraz UD, et al. Sulfated Lactosyl Archaeol Archaeosomes Synergize with Poly(I:C) to Enhance the Immunogenicity and Efficacy of a Synthetic Long Peptide-Based Vaccine in a Melanoma Tumor Model. Pharmaceutics. 2021; 13(2):257. https://doi.org/10.3390/pharmaceutics13020257
Chicago/Turabian StyleAkache, Bassel, Gerard Agbayani, Felicity C. Stark, Yimei Jia, Renu Dudani, Blair A. Harrison, Lise Deschatelets, Vandana Chandan, Edmond Lam, Usha D. Hemraz, and et al. 2021. "Sulfated Lactosyl Archaeol Archaeosomes Synergize with Poly(I:C) to Enhance the Immunogenicity and Efficacy of a Synthetic Long Peptide-Based Vaccine in a Melanoma Tumor Model" Pharmaceutics 13, no. 2: 257. https://doi.org/10.3390/pharmaceutics13020257