
Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review



Abstract
:1. Introduction
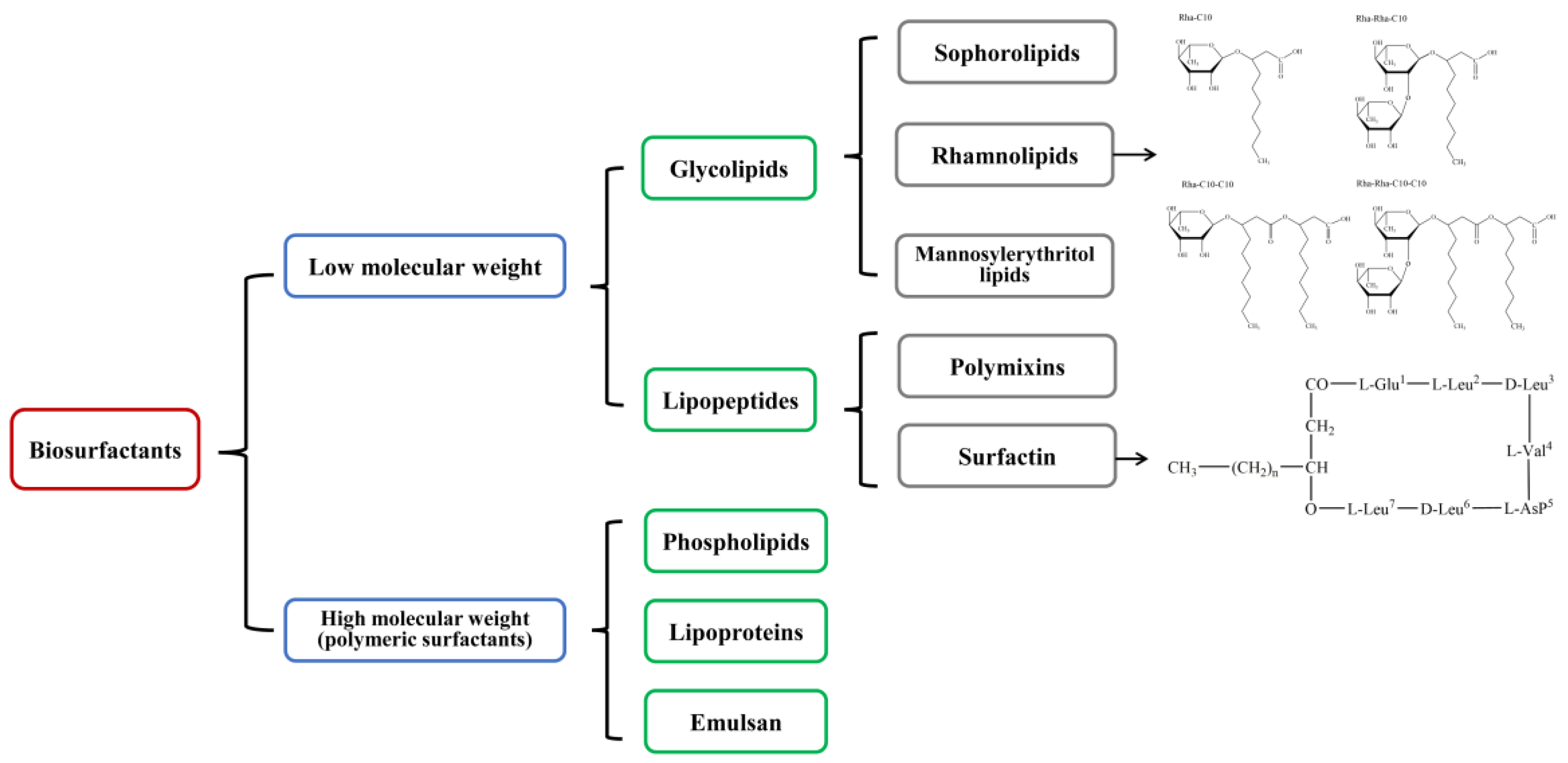
2. Glycolipids
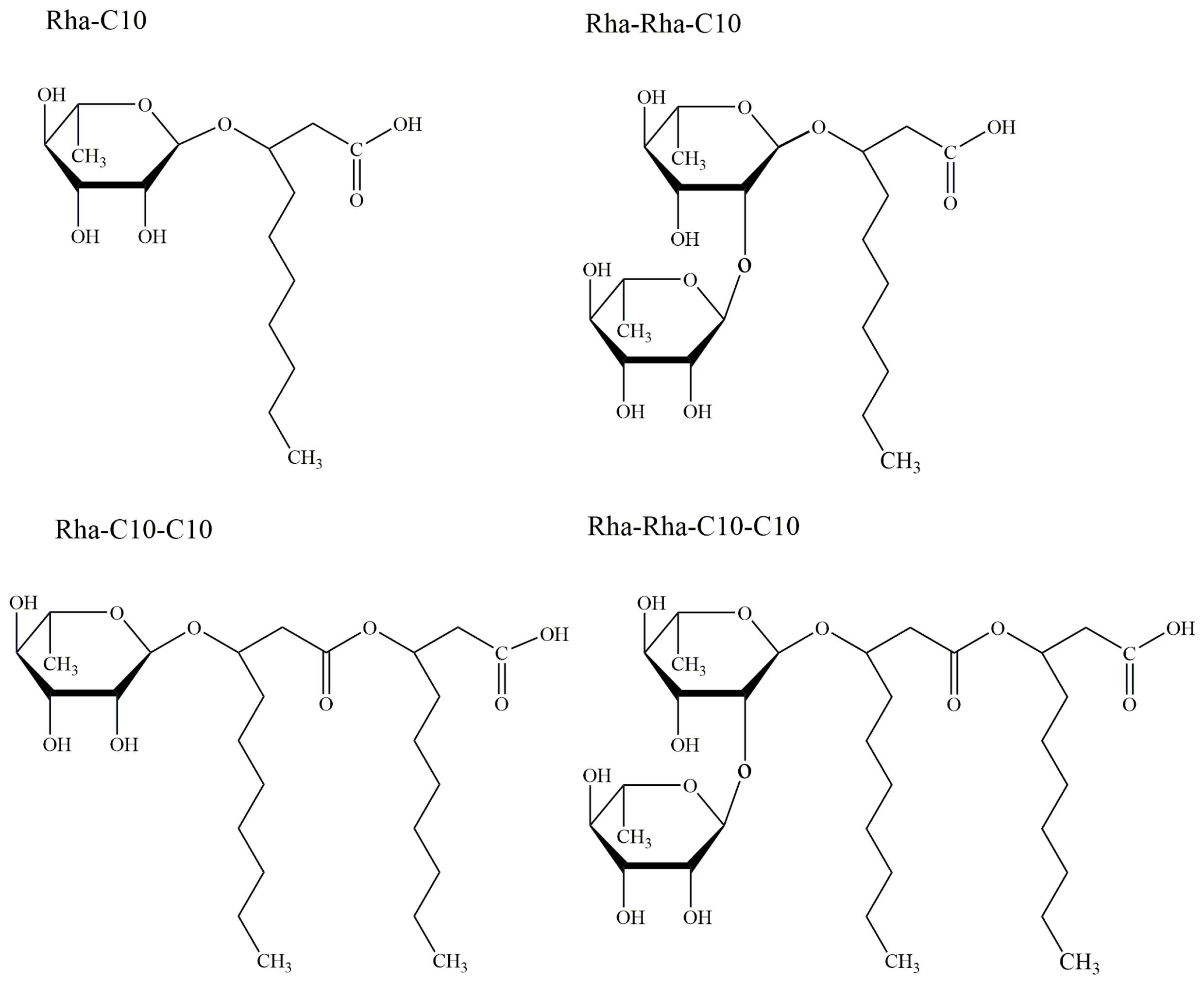
2.1. Rhamnolipids
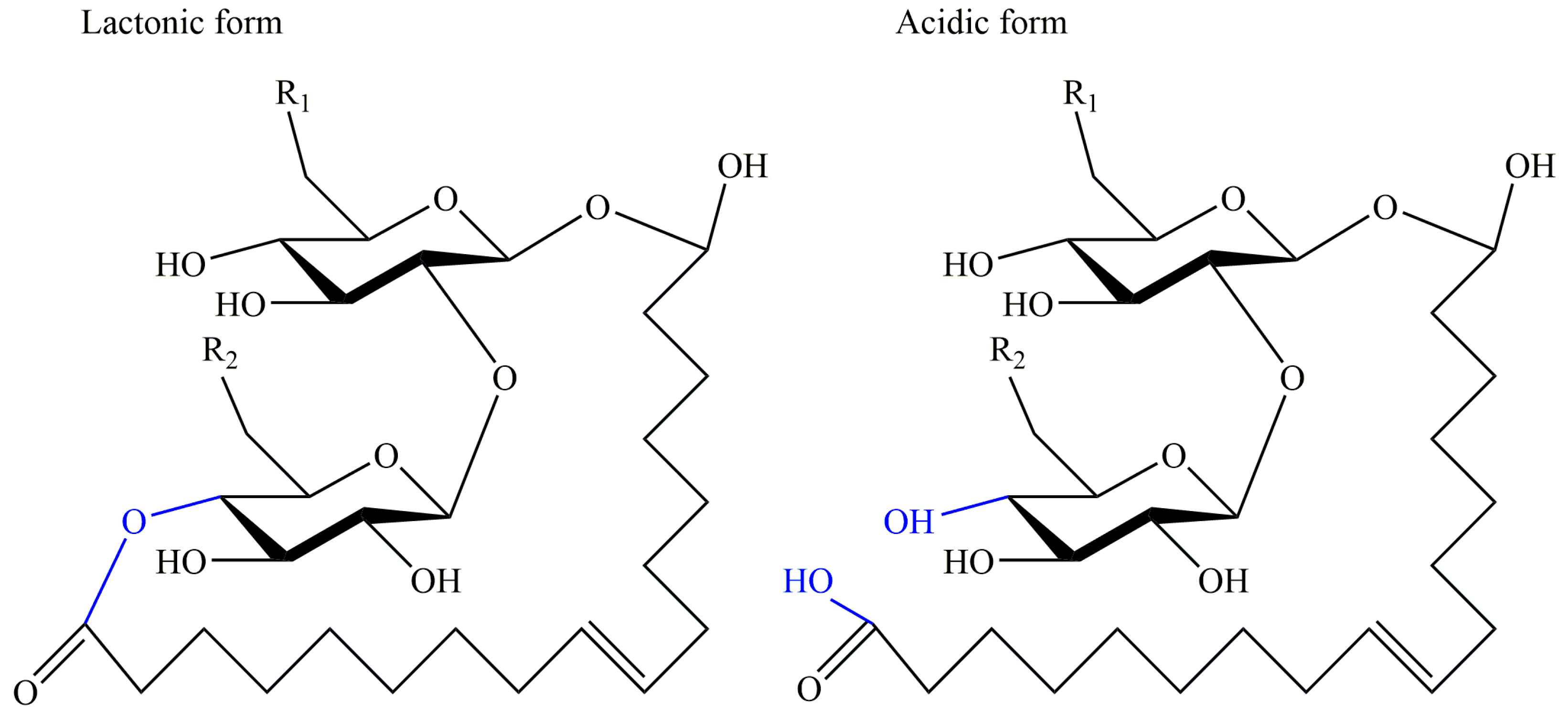
2.2. Sophorolipids
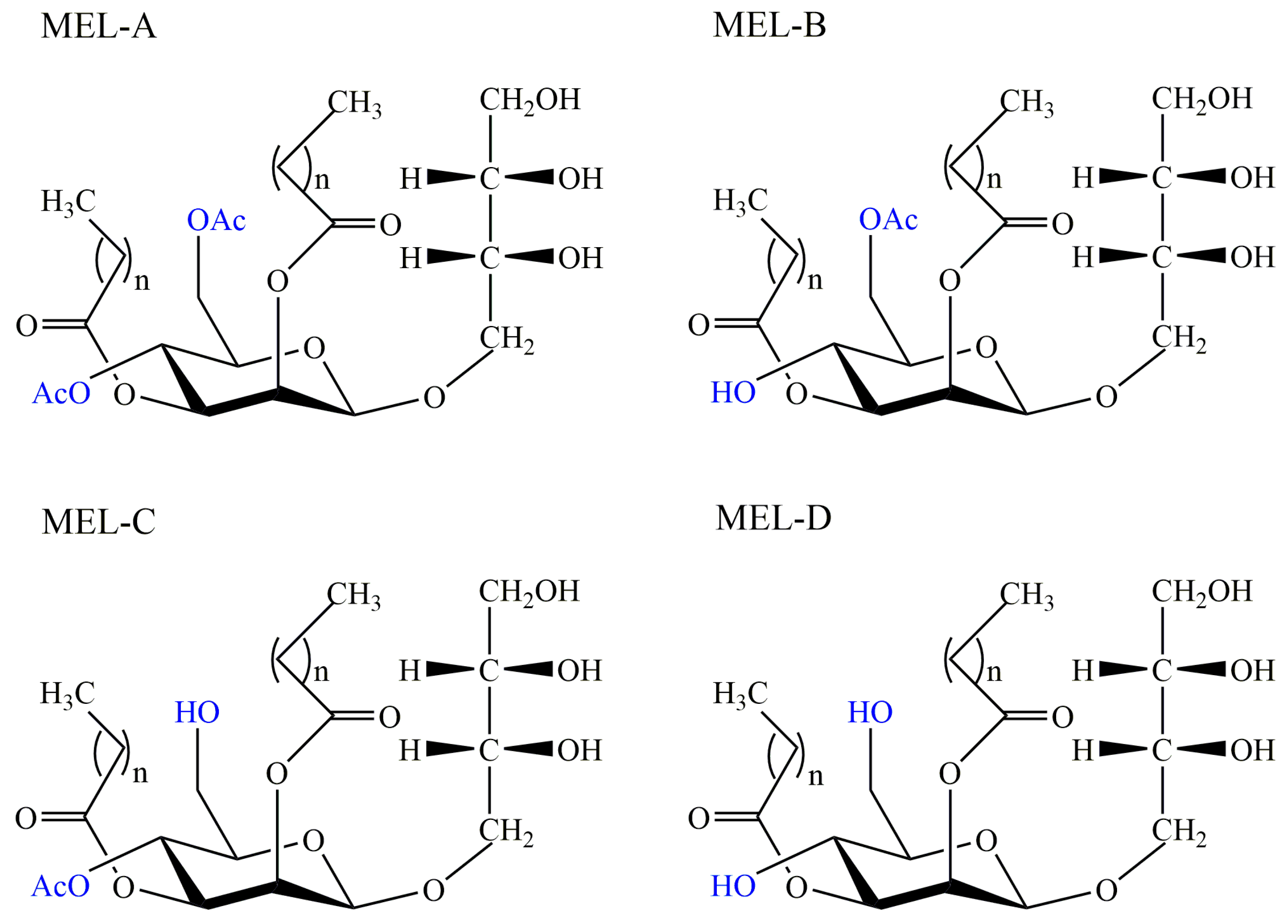
2.3. Mannosylerythritol Lipids
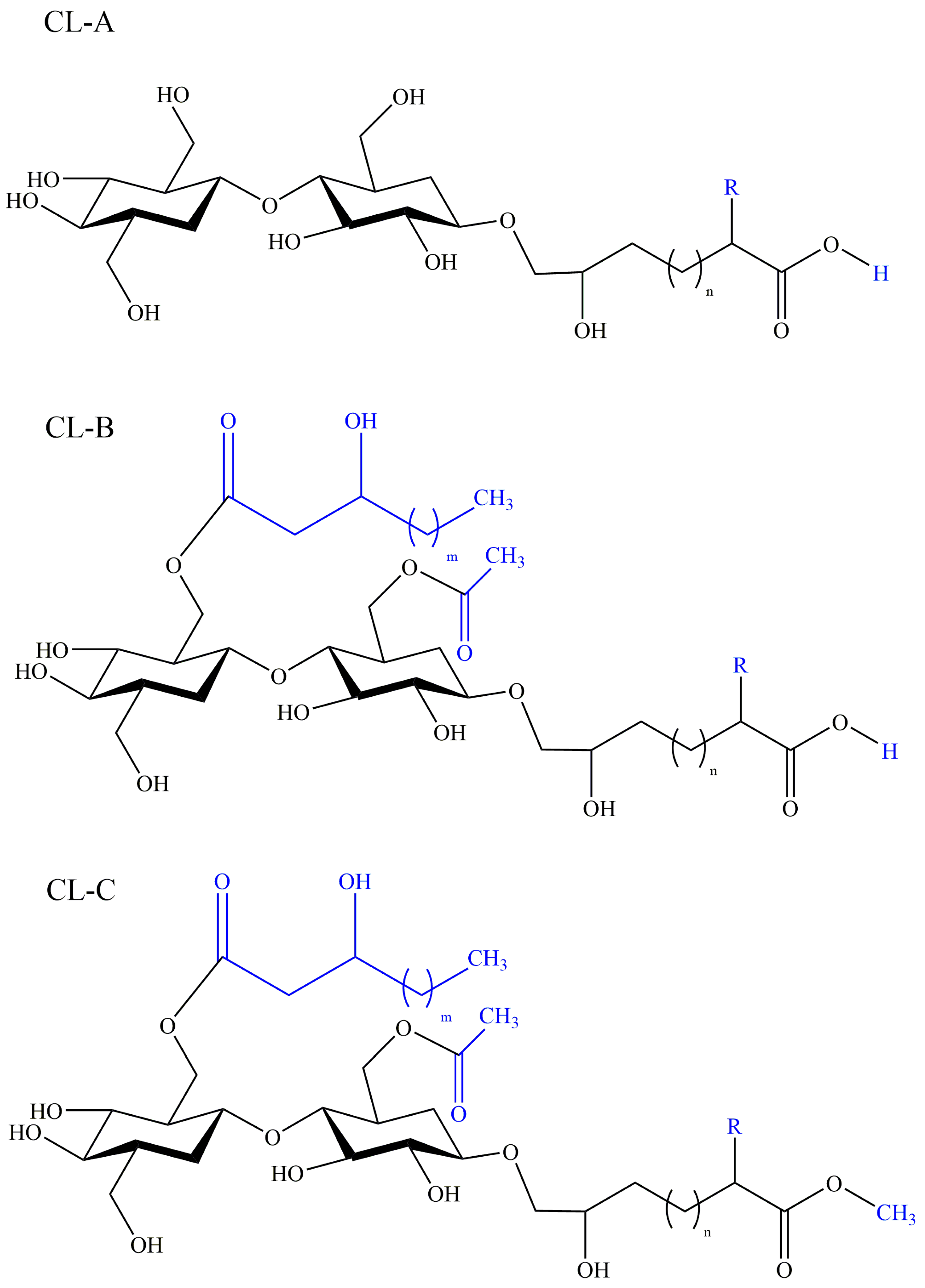
2.4. Cellobiose Lipids
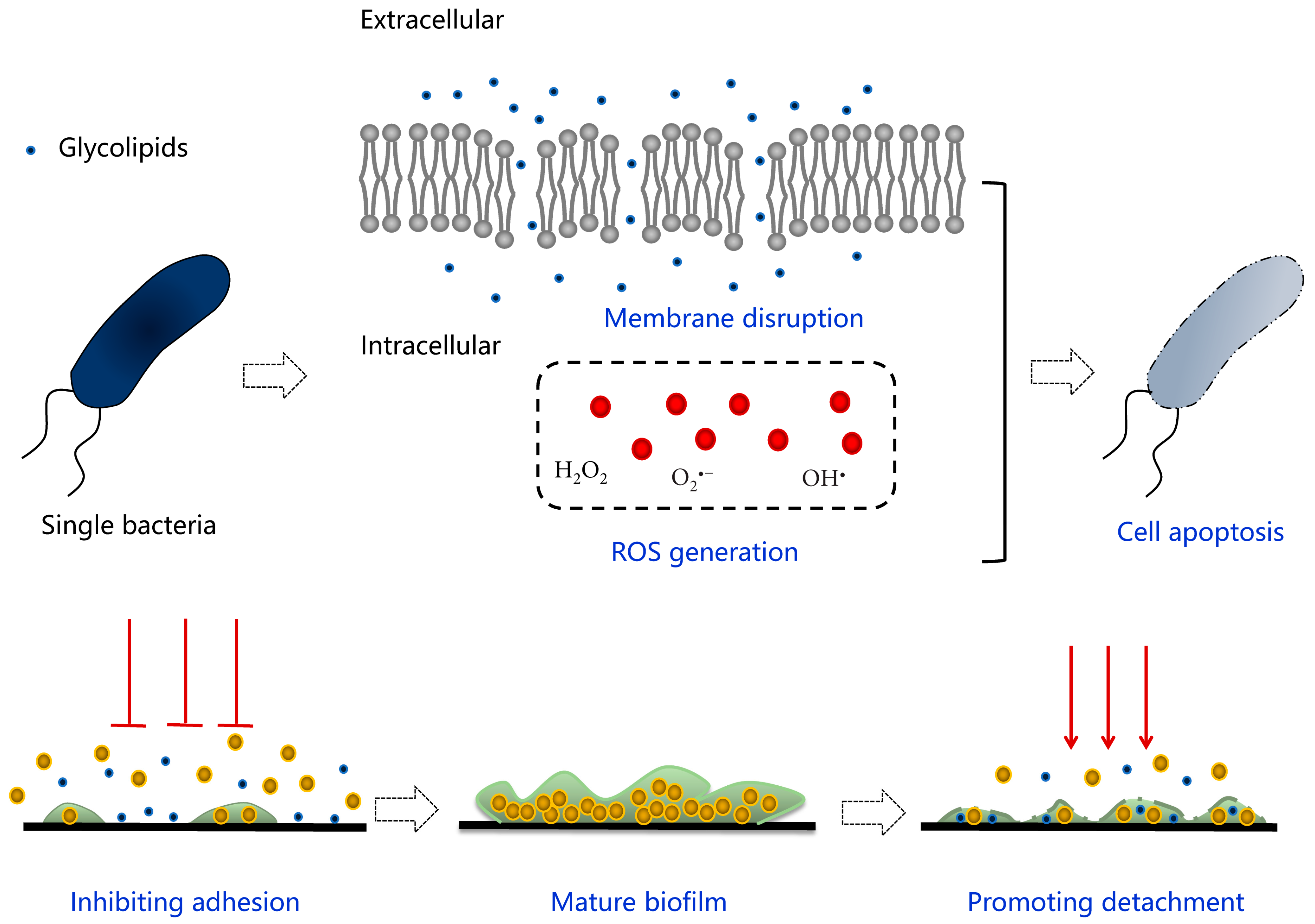
3. Glycolipid-Modified Materials
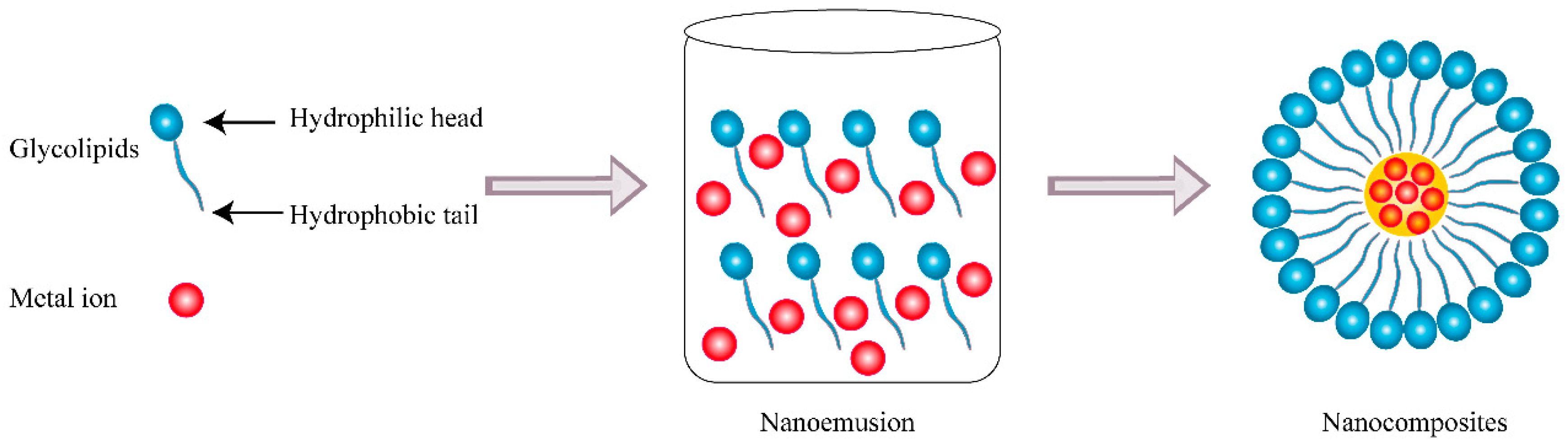
3.1. Nanocomposites
3.2. Liposomes
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Paulino, B.N.; Pessoa, M.G.; Mano, M.C.; Molina, G.; Neri-Numa, I.A.; Pastore, G.M. Current status in biotechnological production and applications of glycolipid biosurfactants. Appl. Microbiol. Biotechnol. 2016, 100, 10265–10293. [Google Scholar] [CrossRef]
- Varvaresou, A.; Iakovou, K. Biosurfactants in cosmetics and biopharmaceuticals. Lett. Appl. Microbiol. 2015, 61, 214–223. [Google Scholar] [CrossRef]
- Jahan, R.; Bodratti, A.M.; Tsianou, M.; Alexandridis, P. Biosurfactants, natural alternatives to synthetic surfactants: Physicochemical properties and applications. Adv. Colloid Interface Sci. 2019, 275, 102061. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Sanchez Ade, J.; Hernandez-Sanchez, H.; Jaramillo-Flores, M.E. Biological activity of glycolipids produced by microorganisms: New trends and possible therapeutic alternatives. Microbiol. Res. 2013, 168, 22–32. [Google Scholar] [CrossRef]
- Shekhar, S.; Sundaramanickam, A.; Balasubramanian, T. Biosurfactant producing microbes and their potential applications: A review. Crit. Rev. Environ. Sci. Technol. 2014, 45, 1522–1554. [Google Scholar] [CrossRef]
- Chen, J.; Wu, Q.; Hua, Y.; Chen, J.; Zhang, H.; Wang, H. Potential applications of biosurfactant rhamnolipids in agriculture and biomedicine. Appl. Microbiol. Biotechnol. 2017, 101, 8309–8319. [Google Scholar] [CrossRef]
- Chen, W.C.; Juang, R.S.; Wei, Y.H. Applications of a lipopeptide biosurfactant, surfactin, produced by microorganisms. Biochem. Eng. J. 2015, 103, 158–169. [Google Scholar] [CrossRef]
- McClements, D.J.; Gumus, C.E. Natural emulsifiers-Biosurfactants, phospholipids, biopolymers, and colloidal particles: Molecular and physicochemical basis of functional performance. Adv. Colloid Interface Sci. 2016, 234, 3–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramani, K.; Jain, S.C.; Mandal, A.B.; Sekaran, G. Microbial induced lipoprotein biosurfactant from slaughterhouse lipid waste and its application to the removal of metal ions from aqueous solution. Colloids Surf. B Biointerfaces 2012, 97, 254–263. [Google Scholar] [CrossRef]
- Mnif, I.; Ellouz-Chaabouni, S.; Ghribi, D. Glycolipid biosurfactants, main classes, functional properties and related potential applications in environmental biotechnology. J. Polym. Environ. 2017, 26, 2192–2206. [Google Scholar] [CrossRef]
- Rahimi, K.; Lotfabad, T.B.; Jabeen, F.; Mohammad Ganji, S. Cytotoxic effects of mono- and di-rhamnolipids from Pseudomonas aeruginosa MR01 on MCF-7 human breast cancer cells. Colloids Surfaces B Biointerfaces 2019, 181, 943–952. [Google Scholar] [CrossRef]
- Hardin, R.; Pierre, J.; Schulze, R.; Mueller, C.M.; Fu, S.L.; Wallner, S.R.; Stanek, A.; Shah, V.; Gross, R.A.; Weedon, J.; et al. Sophorolipids improve sepsis survival: Effects of dosing and derivatives. J. Surg. Res. 2007, 142, 314–319. [Google Scholar] [CrossRef]
- Banat, I.M.; De Rienzo, M.A.; Quinn, G.A. Microbial biofilms: Biosurfactants as antibiofilm agents. Appl. Microbiol. Biotechnol. 2014, 98, 9915–9929. [Google Scholar] [CrossRef] [PubMed]
- Borsanyiova, M.; Patil, A.; Mukherji, R.; Prabhune, A.; Bopegamage, S. Biological activity of sophorolipids and their possible use as antiviral agents. Folia Microbiol. 2016, 61, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Ghribi, D. Glycolipid biosurfactants: Main properties and potential applications in agriculture and food industry. J. Sci. Food Agric. 2016, 96, 4310–4320. [Google Scholar] [CrossRef]
- Jimoh, A.A.; Lin, J. Biosurfactant: A new frontier for greener technology and environmental sustainability. Ecotoxicol. Environ. Saf. 2019, 184, 109607. [Google Scholar] [CrossRef] [PubMed]
- Eslami, P.; Hajfarajollah, H.; Bazsefidpar, S. Recent advancements in the production of rhamnolipid biosurfactants by Pseudomonas aeruginosa. RSC Adv. 2020, 10, 34014–34032. [Google Scholar] [CrossRef]
- Bergstrom, S.; Theorell, H.; Davide, H. On a metabolic product of ps pyocyanea, pyolipic acid, active against mycobact tuberculosis. Arkiv Kemi Mineral. Och Geologi 1947, 23, 1–12. [Google Scholar]
- Bergstrom, S.; Theorell, H.; Davide, H. Pyolipic acid, a metabolic product of Pseudomonas pyocyanea, active against Mycobacterium tuberculosis. Arch. Biochem. 1946, 10, 165–166. [Google Scholar]
- Jarvis, F.G.; Johnson, M.J. A glyco-lipide produced by Pseudomonas aeruginosa. J. Am. Chem. Soc. 1949, 71, 4124–4126. [Google Scholar] [CrossRef]
- Edwards, J.R.; Hayashi, J.A. Structure of a rhamnolipid from Pseudomonas aeruginosa. Arch. Biochem. Biophys. 1965, 111, 415–421. [Google Scholar] [CrossRef]
- Geys, R.; Soetaert, W.; Van Bogaert, I. Biotechnological opportunities in biosurfactant production. Curr. Opin Biotechnol. 2014, 30, 66–72. [Google Scholar] [CrossRef]
- Dubeau, D.; Deziel, E.; Woods, D.E.; Lepine, F. Burkholderia thailandensis harbors two identical rhl gene clusters responsible for the biosynthesis of rhamnolipids. BMC Microbiol. 2009, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Andrä, J.; Rademann, J.; Howe, J.; Koch, M.H.J.; Heine, H.; Zähringer, U.; Brandenburg, K. Endotoxin-like properties of a rhamnolipid exotoxin from Burkholderia (Pseudomonas) plantarii: Immune cell stimulation and biophysical characterization. Biol. Chem. 2006, 387, 301–310. [Google Scholar] [CrossRef]
- Howe, J.; Bauer, J.; Andra, J.; Schromm, A.B.; Ernst, M.; Rossle, M.; Zahringer, U.; Rademann, J.; Brandenburg, K. Biophysical characterization of synthetic rhamnolipids. FEBS J. 2006, 273, 5101–5112. [Google Scholar] [CrossRef] [Green Version]
- Itoh, S.H.; Tomita, F.; Suzuki, T. Rhamnolipids produced by Pseudomonas aeruginosa grown on n-paraffin (mixture of C 12, C 13 and C 14 fractions). J. Antibiot. 1971, 24, 855–859. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Maniglio, D.; Tambone, E.; Carmagnola, I.; Fedeli, E.; Caola, I.; Nollo, G.; Chiono, V.; Allegrone, G.; et al. Medical-grade silicone coated with rhamnolipid R89 is effective against Staphylococcus spp. biofilms. Molecules 2019, 24, 3843. [Google Scholar] [CrossRef] [Green Version]
- De Freitas Ferreira, J.; Vieira, E.A.; Nitschke, M. The antibacterial activity of rhamnolipid biosurfactant is pH dependent. Food Res. Int. 2019, 116, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.; Teruel, J.A.; Espuny, M.J.; Marques, A.; Aranda, F.J.; Manresa, A.; Ortiz, A. Modulation of the physical properties of dielaidoylphosphatidylethanolamine membranes by a dirhamnolipid biosurfactant produced by Pseudomonas aeruginosa. Chem. Phys. Lipids 2006, 142, 118–127. [Google Scholar] [CrossRef]
- Araujo, L.V.D.; Guimarães, C.R.; Marquita, R.L.D.S.; Santiago, V.M.J.; De Souza, M.P.; Nitschke, M.; Freire, D.M.G. Rhamnolipid and surfactin: Anti-adhesion/antibiofilm and antimicrobial effects. Food Control. 2016, 63, 171–178. [Google Scholar] [CrossRef]
- Abdollahi, S.; Tofighi, Z.; Babaee, T.; Shamsi, M.; Rahimzadeh, G.; Rezvanifar, H.; Saeidi, E.; Mohajeri Amiri, M.; Saffari Ashtiani, Y.; Samadi, N. Evaluation of anti-oxidant and anti-biofilm activities of biogenic surfactants derived from Bacillus amyloliquefaciens and Pseudomonas aeruginosa. Iran J. Pharm. Res. 2020, 19, 115–126. [Google Scholar]
- Elshikh, M.; Funston, S.; Chebbi, A.; Ahmed, S.; Marchant, R.; Banat, I.M. Rhamnolipids from non-pathogenic Burkholderia thailandensis E264: Physicochemical characterization, antimicrobial and antibiofilm efficacy against oral hygiene related pathogens. New Biotechnol. 2017, 36, 26–36. [Google Scholar] [CrossRef]
- Kim, L.H.; Jung, Y.; Yu, H.W.; Chae, K.J.; Kim, I.S. Physicochemical interactions between rhamnolipids and Pseudomonas aeruginosa biofilm layers. Environ. Sci. Technol. 2015, 49, 3718–3726. [Google Scholar] [CrossRef]
- Kim, B.S.; Lee, J.Y.; Hwang, B.K. In vivo control and in vitro antifungal activity of rhamnolipid B, a glycolipid antibiotic, against Phytophthora capsici and Colletotrichum orbiculare. Pest. Manag. Sci. 2000, 56, 1029–1035. [Google Scholar] [CrossRef]
- Goswami, D.; Borah, S.N.; Lahkar, J.; Handique, P.J.; Deka, S. Antifungal properties of rhamnolipid produced by Pseudomonas aeruginosa DS9 against Colletotrichum falcatum. J. Basic Microbiol. 2015, 55, 1265–1274. [Google Scholar] [CrossRef]
- Sen, S.; Borah, S.N.; Kandimalla, R.; Bora, A.; Deka, S. Efficacy of a rhamnolipid biosurfactant to inhibit Trichophyton rubrum in vitro and in a mice model of dermatophytosis. Exp. Dermatol. 2019, 28, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Pemmaraju, S.C.; Pruthi, P.A.; Cameotra, S.S.; Pruthi, V. Candida biofilm disrupting ability of di-rhamnolipid (RL-2) produced from Pseudomonas aeruginosa DSVP20. Appl. Biochem. Biotechnol. 2013, 169, 2374–2391. [Google Scholar] [CrossRef] [PubMed]
- Devendra, H.; Dusane, S.D. Yarlagadda V Nancharaiah, Ameeta Ravi Kumar, Vayalam P. Venugopalan and Smita S Zinjarde*, Disruption of Yarrowia lipolytica biofilms by rhamnolipid biosurfactant. Aquat. Biosyst. 2012, 8, 1–7. [Google Scholar]
- Rodrigues, L.R.; Banat, I.M.; Van der Mei, H.C.; Teixeira, J.A.; Oliveira, R. Interference in adhesion of bacteria and yeasts isolated from explanted voice prostheses to silicone rubber by rhamnolipid biosurfactants. J. Appl. Microbiol. 2006, 100, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid-biosurfactant permeabilizing effects on gram-positive and gram-negative bacterial strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef]
- Sotirova, A.; Avramova, T.; Stoitsova, S.; Lazarkevich, I.; Lubenets, V.; Karpenko, E.; Galabova, D. The importance of rhamnolipid-biosurfactant-induced changes in bacterial membrane lipids of Bacillus subtilis for the antimicrobial activity of thiosulfonates. Curr. Microbiol. 2012, 65, 534–541. [Google Scholar] [CrossRef]
- Chen, X.; Li, P.; Shen, Y.; Zou, Y.; Yuan, G.; Hu, H. Rhamnolipid-involved antibiotics combinations improve the eradication of Helicobacter pylori biofilm in vitro: A comparison with conventional triple therapy. Microb. Pathog. 2019, 131, 112–119. [Google Scholar] [CrossRef]
- Jezierska, S.; Claus, S.; Van Bogaert, I. Yeast glycolipid biosurfactants. FEBS Lett. 2018, 592, 1312–1329. [Google Scholar] [CrossRef] [Green Version]
- Gorin, P.A.J.; Spencer, J.F.T.; Tulloch, A.P. Hydroxy Fatty acid glycosides of sophorose from Torulopsis magnoliae. Can. J. Chem. 1961, 39, 846–855. [Google Scholar] [CrossRef] [Green Version]
- Tulloch, A.P.; Spencer, J.F.T.; Deinema, M.H. A new hydroxy fatty acid sophoroside from Candida bogoriensis. Can. J. Chem. 1968, 46, 345–348. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Price, N.P.J.; Ray, K.J.; Kuo, T.-M. Production of sophorolipid biosurfactants by multiple species of the Starmerella (Candida) bombicola yeast clade. FEMS Microbiol. Lett. 2010, 311, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, T.H.G.M.; Lee, J.; Chen, J.; Song, X.; Zhang, H.; Qu, Y. Production, structure elucidation and anticancer properties of sophorolipid from Wickerhamiella domercqiae. Enzyme Microb. Technol. 2006, 39, 501–506. [Google Scholar]
- Van Bogaert, I.N.; Saerens, K.; De Muynck, C.; Develter, D.; Soetaert, W.; Vandamme, E.J. Microbial production and application of sophorolipids. Appl. Microbiol. Biotechnol. 2007, 76, 23–34. [Google Scholar] [CrossRef]
- Shin, J.D.; Lee, J.; Kim, Y.B.; Han, I.-S.; Kim, E.-K. Production and characterization of methyl ester sophorolipids with 22-carbon-fatty acids. Bioresour. Technol. 2010, 101, 3170–3174. [Google Scholar] [CrossRef] [PubMed]
- Van Bogaert, I.N.A.; Zhang, J.; Soetaert, W. Microbial synthesis of sophorolipids. Process Biochem. 2011, 46, 821–833. [Google Scholar] [CrossRef]
- Ankulkar, R.; Chavan, M. Characterisation and application studies of sophorolipid biosurfactant by Candida tropicalis RA1. J. Pure Appl. Microbiol. 2019, 13, 1653–1665. [Google Scholar] [CrossRef] [Green Version]
- Fontoura, I.C.C.D.; Saikawa, G.I.A.; Silveira, V.A.I.; Pan, N.C.; Amador, I.R.; Baldo, C.; Rocha, S.P.D.D.; Celligoi, M.A.P.C. Antibacterial activity of sophorolipids from Candida bombicola against human pathogens. Braz. Arch. Biol. Technol. 2020, 63, e20180568. [Google Scholar] [CrossRef]
- Gaur, V.K.; Regar, R.K.; Dhiman, N.; Gautam, K.; Srivastava, J.K.; Patnaik, S.; Kamthan, M.; Manickam, N. Biosynthesis and characterization of sophorolipid biosurfactant by Candida spp.: Application as food emulsifier and antibacterial agent. Bioresour. Technol. 2019, 285, 121314. [Google Scholar] [CrossRef] [PubMed]
- Solaiman, D.K.Y.; Ashby, R.D.; Uknalis, J. Characterization of growth inhibition of oral bacteria by sophorolipid using a microplate-format assay. J. Microbiol. Methods 2017, 136, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Silveira, V.A.I.; Nishio, E.K.; Freitas, C.A.U.Q.; Amador, I.R.; Kobayashi, R.K.T.; Caretta, T.; Macedo, F.; Celligoi, M.A.P.C. Production and antimicrobial activity of sophorolipid against Clostridium perfringens and Campylobacter jejuni and their additive interaction with lactic acid. Biocatal. Agric. Biotechnol. 2019, 21, 101287. [Google Scholar] [CrossRef]
- Ceresa, C.; Fracchia, L.; Williams, M.; Banat, I.M.; Díaz De Rienzo, M.A. The effect of sophorolipids against microbial biofilms on medical-grade silicone. J. Biotechnol. 2020, 309, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Borah, S.N.; Bora, A.; Deka, S. Production, characterization, and antifungal activity of a biosurfactant produced by Rhodotorula babjevae YS3. Microb. Cell Factories 2017, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Haque, F.; Alfatah, M.; Ganesan, K.; Bhattacharyya, M.S. Inhibitory effect of sophorolipid on Candida albicans biofilm formation and hyphal growth. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Haque, F.; Verma, N.K.; Alfatah, M.; Bijlani, S.; Bhattacharyya, M.S. Sophorolipid exhibits antifungal activity by ROS mediated endoplasmic reticulum stress and mitochondrial dysfunction pathways in Candida albicans. RSC Adv. 2019, 9, 41639–41648. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, X.; Fu, S.; An, Z.; Feng, Y.; Wang, R.; Ji, P. Effects of sophorolipids on fungal and oomycete pathogens in relation to pH solubility. J. Appl. Microbiol. 2020, 128, 1754–1763. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, X.; Solaiman, D.K.Y.; Ashby, R.D.; Liu, Z.; Mukhopadhyay, S.; Yan, R. Inactivation of Escherichia coli O157:H7 in vitro and on the surface of spinach leaves by biobased antimicrobial surfactants. Food Control 2016, 60, 158–165. [Google Scholar] [CrossRef]
- Sen, S.; Borah, S.N.; Kandimalla, R.; Bora, A.; Deka, S. Sophorolipid biosurfactant can control cutaneous dermatophytosis caused by Trichophyton mentagrophytes. Front Microbiol. 2020, 11, 329. [Google Scholar] [CrossRef] [Green Version]
- Arutchelvi, J.I.; Bhaduri, S.; Uppara, P.V.; Doble, M. Mannosylerythritol lipids: A review. J. Ind. Microbiol. Biotechnol. 2008, 35, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Haskins, R.H.; Thorn, J.A.; Boothroyd, B. Biochemistry of the ustilaginales: XI. metabolic products of Ustilago zeae in submerged culture. Can. J. Microbiol. 1955, 1, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Koike, H.; Koyama, Y.; Hagiwara, H.; Ito, E.; Fukuoka, T.; Imura, T.; Machida, M.; Kitamoto, D. Genome Sequence of the Basidiomycetous yeast Pseudozyma antarctica T-34, a producer of the glycolipid biosurfactants mannosylerythritol lipids. Genome Announc. 2013, 1, e0006413. [Google Scholar] [CrossRef] [Green Version]
- Saika, A.; Koike, H.; Fukuoka, T.; Yamamoto, S.; Kishimoto, T.; Morita, T. A Gene cluster for biosynthesis of mannosylerythritol lipids consisted of 4-O-beta-D-Mannopyranosyl-(2R,3S)-Erythritol as the sugar moiety in a Basidiomycetous yeast Pseudozyma tsukubaensis. PLoS ONE 2016, 11, e0157858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Hatada, Y.; Horiuchi, J. Draft genome sequence of the Basidiomycetous yeast-like fungus Pseudozyma hubeiensis SY62, which produces an abundant amount of the biosurfactant mannosylerythritol lipids. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Saika, A.; Koike, H.; Fukuoka, T.; Morita, T. Tailor-made mannosylerythritol lipids: Current state and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 6877–6884. [Google Scholar] [CrossRef]
- Kitamoto, D.; Yanagishita, H.; Endo, A.; Nakaiwa, M.; Nakane, T.; Akiya, T. Remarkable antiagglomeration effect of a yeast biosurfactant, diacylmannosylerythritol, on ice-water slurry for cold thermal storage. Biotechnol. Prog. 2001, 17, 362–365. [Google Scholar] [CrossRef]
- Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Mannosylerythritol lipids: Production and applications. J. Oleo Sci. 2015, 64, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Kitamoto, D. Surface active properties and antimicrobial activities of mannosylerythritol lipids as biosurfactants produced by Candida antarctica. Sci. World J. 1993, 29, 91–96. [Google Scholar] [CrossRef]
- Fukuoka, T.; Morita, T.; Konishi, M.; Imura, T.; Sakai, H.; Kitamoto, D. Structural characterization and surface-active properties of a new glycolipid biosurfactant, mono-acylated mannosylerythritol lipid, produced from glucose by Pseudozyma antarctica. Appl. Microbiol. Biotechnol. 2007, 76, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Recke, V.K.; Beyrle, C.; Gerlitzki, M.; Hausmann, R.; Syldatk, C.; Wray, V.; Tokuda, H.; Suzuki, N.; Lang, S. Lipase-catalyzed acylation of microbial mannosylerythritol lipids (biosurfactants) and their characterization. Carbohydr. Res. 2013, 373, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Okuhira, K.; Koike, S.; Ito, S.; Kobayashi, Y. The bio-surfactant mannosylerythritol lipid acts as a selective antibacterial agent to modulate rumen fermentation. Anim. Sci. J. 2020, 91, e13464. [Google Scholar] [CrossRef]
- Nashida, J.; Nishi, N.; Takahashi, Y.; Hayashi, C.; Igarashi, M.; Takahashi, D.; Toshima, K. Systematic and stereoselective total synthesis of mannosylerythritol lipids and evaluation of their antibacterial activity. J. Org. Chem. 2018, 83, 7281–7289. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Niu, Y.; Zhao, W.; Chen, Q. Antibacterial activity of mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Shu, Q.; Wei, T.; Lu, H.; Niu, Y.; Chen, Q. Mannosylerythritol lipids: Dual inhibitory modes against Staphylococcus aureus through membrane-mediated apoptosis and biofilm disruption. Appl. Microbiol. Biotechnol. 2020, 104, 5053–5064. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shu, Q.; Chen, Q.; Pang, X.; Wu, Y.; Zhou, W.; Wu, Y.; Niu, J.; Zhang, X. Antibacterial efficacy and mechanism of mannosylerythritol lipids-A on Listeria monocytogenes. Molecules 2020, 25, 4857. [Google Scholar] [CrossRef]
- Yoshida, S.; Koitabashi, M.; Nakamura, J.; Fukuoka, T.; Sakai, H.; Abe, M.; Kitamoto, D.; Kitamoto, H. Effects of biosurfactants, mannosylerythritol lipids, on the hydrophobicity of solid surfaces and infection behaviours of plant pathogenic fungi. J. Appl. Microbiol. 2015, 119, 215–224. [Google Scholar] [CrossRef]
- Merghni, A.; Dallel, I.; Noumi, E.; Kadmi, Y.; Hentati, H.; Tobji, S.; Ben Amor, A.; Mastouri, M. Antioxidant and antiproliferative potential of biosurfactants isolated from Lactobacillus casei and their anti-biofilm effect in oral Staphylococcus aureus strains. Microb. Pathog. 2017, 104, 84–89. [Google Scholar] [CrossRef]
- Haskins, R.H.; Thorn, J.A. Biochemistry of the ustilaginales: VII. antibiotic activity of ustilagic acid. Can. J. Botany 1951, 29, 585–592. [Google Scholar] [CrossRef]
- Spoeckner, S.; Wray, V.; Nimtz, M.; Lang, S. Glycolipids of the smut fungus Ustilago maydis from cultivation on renewable resources. Appl. Microbiol. Biotechnol. 1999, 51, 33–39. [Google Scholar] [CrossRef]
- Kulakovskaya, E.V.; Kulakovskaya, T.V.; Golubev, V.I.; Shashkov, A.S.; Grachev, A.A.; Nifantiev, N.E. Fungicidal activity of cellobiose lipids from culture broth of yeast Cryptococcus humicola and Pseudozyma fusiformata. Russ. J. Bioorganic Chem. 2007, 33, 156–160. [Google Scholar] [CrossRef]
- Golubev, W.I.; Kulakovskaya, T.V.; Shashkov, A.S.; Kulakovskaya, E.V.; Golubev, N.V. Antifungal cellobiose lipid secreted by the epiphytic yeast Pseudozyma graminicola. Microbiology 2008, 77, 171–175. [Google Scholar] [CrossRef]
- Mimee, B.; Labbe, C.; Belanger, R.R. Catabolism of flocculosin, an antimicrobial metabolite produced by Pseudozyma flocculosa. Glycobiology 2009, 19, 995–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oraby, A.; Werner, N.; Sungur, Z.; Zibek, S. Factors affecting the synthesis of cellobiose lipids by Sporisorium scitamineum. Front. Bioeng. Biotechnol. 2020, 8, 555647. [Google Scholar] [CrossRef]
- Ludmila, V.; Trilisenko, E.V.K.; Kulakovskaya, T.V.; Yu Ivanov, A.; Penkov, N.V.; Vagabov, V.M.; Kulaev, I.S. The antifungal effect of cellobiose lipid on the cells of Saccharomyces cerevisiae depends on carbon source. Springerplus 2012, 1, 1–6. [Google Scholar]
- Puchkov, E.O.; Zähringer, U.; Lindner, B.; Kulakovskaya, T.V.; Seydel, U.; Wiese, A. Mycocidal, membrane-active complex of Cryptococcus humicola is a new type of cellobiose lipid with detergent features. Biochim. Biophys. Acta 2002, 1558, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Kulakovskaya, E.V.; Ivanov, A.Y.; Kulakovskaya, T.V.; Vagabov, V.M.; Kulaev, I.S. Effects of cellobiose lipid B on Saccharomyces cerevisiae cells: K+ leakage and inhibition of polyphosphate accumulation. Microbiology 2008, 77, 288–292. [Google Scholar] [CrossRef]
- Mimee, B.; Labbe, C.; Pelletier, R.; Belanger, R.R. Antifungal activity of flocculosin, a novel glycolipid isolated from Pseudozyma flocculosa. Antimicrob. Agents Chemother. 2005, 49, 1597–1599. [Google Scholar] [CrossRef] [Green Version]
- Mimee, B.; Pelletier, R.; Belanger, R.R. In vitro antibacterial activity and antifungal mode of action of flocculosin, a membrane-active cellobiose lipid. J. Appl. Microbiol. 2009, 107, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Chen, Y.; Wang, Y.; Jiang, H.; Wang, X. Advances and challenges in metallic nanomaterial synthesis and antibacterial applications. J. Mater. Chem. B 2020, 8, 4764–4777. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Ehsani, A.; Jafari, S.M. Lipid-based nano delivery of antimicrobials to control food-borne bacteria. Adv. Colloid Interface Sci. 2019, 270, 263–277. [Google Scholar] [CrossRef]
- Kasture, M.B.; Patel, P.; Prabhune, A.A.; Ramana, C.V.; Kulkarni, A.A.; Prasad, B.L.V. Synthesis of silver nanoparticles by sophorolipids: Effect of temperature and sophorolipid structure on the size of particles. J. Chem. Sci. 2008, 120, 515–520. [Google Scholar] [CrossRef]
- Xin, Q.; Shah, H.; Nawaz, A.; Xie, W.; Akram, M.Z.; Batool, A.; Tian, L.; Jan, S.U.; Boddula, R.; Guo, B.; et al. Antibacterial carbon-based nanomaterials. Adv. Mater. 2019, 31, e1804838. [Google Scholar] [CrossRef]
- Basak, G.; Das, D.; Das, N. Dual role of acidic diacetate sophorolipid as biostabilizer for ZnO nanoparticle synthesis and biofunctionalizing agent against Salmonella enterica and Candida albicans. J. Microbiol. Biotechnol. 2014, 24, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Khalid, H.F.; Tehseen, B.; Sarwar, Y.; Hussain, S.Z.; Khan, W.S.; Raza, Z.A.; Bajwa, S.Z.; Kanaras, A.G.; Hussain, I.; Rehman, A. Biosurfactant coated silver and iron oxide nanoparticles with enhanced anti-biofilm and anti-adhesive properties. J. Hazard Mater. 2019, 364, 441–448. [Google Scholar] [CrossRef]
- Das, M.; Patowary, K.; Vidya, R.; Malipeddi, H. Microemulsion synthesis of silver nanoparticles using biosurfactant extracted from Pseudomonas aeruginosa MKVIT3 strain and comparison of their antimicrobial and cytotoxic activities. IET Nanobiotechnol. 2016, 10, 411–418. [Google Scholar] [CrossRef]
- Joanna, C.; Marcin, L.; Ewa, K.; Grazyna, P. A nonspecific synergistic effect of biogenic silver nanoparticles and biosurfactant towards environmental bacteria and fungi. Ecotoxicology 2018, 27, 352–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidyarani, N.; Srivastav, A.K.; Gupta, S.K.; Kumar, U. Synthesis and physicochemical characterization of rhamnolipid-stabilized carvacrol-loaded zein nanoparticles for antimicrobial application supported by molecular docking. J. Nanoparticle Res. 2020, 22, 1–13. [Google Scholar] [CrossRef]
- Marangon, C.A.; Martins, V.C.A.; Ling, M.H.; Melo, C.C.; Plepis, A.M.G.; Meyer, R.L.; Nitschke, M. Combination of rhamnolipid and chitosan in nanoparticles boosts their antimicrobial efficacy. ACS Appl. Mater. Interfaces 2020, 12, 5488–5499. [Google Scholar] [CrossRef]
- Shikha, S.; Chaudhuri, S.R.; Bhattacharyya, M.S. Facile one pot greener synthesis of sophorolipid capped gold nanoparticles and its antimicrobial activity having special efficacy against gram negative Vibrio cholerae. Sci. Rep. 2020, 10, 1463. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shu, Q.; Niu, Y.; Jiao, Y.; Chen, Q. Preparation, characterization, and antibacterial effects of chitosan nanoparticles embedded with essential oils synthesized in an Ionic liquid containing system. J. Agric. Food Chem. 2018, 66, 7006–7014. [Google Scholar] [CrossRef]
- Bakur, A.; Niu, Y.; Kuang, H.; Chen, Q. Synthesis of gold nanoparticles derived from mannosylerythritol lipid and evaluation of their bioactivities. AMB Express 2019, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.; Prabhune, A.A. Photophysical studies on curcumin-sophorolipid nanostructures: Applications in quorum quenching and imaging. R. Soc. Open Sci. 2018, 5, 170865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakur, A.; Elshaarani, T.; Niu, Y.; Chen, Q. Comparative study of antidiabetic, bactericidal, and antitumor activities of MEL@AgNPs, MEL@ZnONPs, and Ag–ZnO/MEL/GA nanocomposites prepared by using MEL and gum arabic. RSC Adv. 2019, 9, 9745–9754. [Google Scholar] [CrossRef] [Green Version]
- Filipczak, N.; Pan, J.; Yalamarty, S.S.K.; Torchilin, V.P. Recent advancements in liposome technology. Adv. Drug Deliv. Rev. 2020, 156, 4–22. [Google Scholar] [CrossRef]
- Barenholz, Y. Doxil(R)--the first FDA-approved nano-drug: Lessons learned. J. Controll. Release Off. J. Controlled Release Soc. 2012, 160, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Manca, M.L.; Sinico, C.; Maccioni, A.M.; Diez, O.; Fadda, A.M.; Manconi, M. Composition influence on pulmonary delivery of rifampicin liposomes. Pharmaceutics 2012, 4, 590–606. [Google Scholar] [CrossRef]
- Manca, M.L.; Valenti, D.; Sales, O.D.; Nacher, A.; Fadda, A.M.; Manconi, M. Fabrication of polyelectrolyte multilayered vesicles as inhalable dry powder for lung administration of rifampicin. Int. J. Pharm. 2014, 472, 102–109. [Google Scholar] [CrossRef]
- Melis, V.; Manca, M.L.; Bullita, E.; Tamburini, E.; Castangia, I.; Cardia, M.C.; Valenti, D.; Fadda, A.M.; Peris, J.E.; Manconi, M.; et al. Inhalable polymer-glycerosomes as safe and effective carriers for rifampicin delivery to the lungs. Coll. Surf. B Biointerfaces 2016, 143, 301–308. [Google Scholar] [CrossRef]
- Fan, L.; Xie, P.; Wang, Y.; Liu, X.; Li, Y.; Zhou, J. Influences of mannosylerythritol lipid-A on the self-assembling structure formation and functional properties of heat-induced β-lactoglobulin aggregates. Food Hydrocoll. 2019, 96, 310–321. [Google Scholar] [CrossRef]
- Wu, J.; Niu, Y.; Jiao, Y.; Chen, Q. Fungal chitosan from Agaricus bisporus (Lange) Sing. Chaidam increased the stability and antioxidant activity of liposomes modified with biosurfactants and loading betulinic acid. Int. J. Biol. Macromol. 2019, 123, 291–299. [Google Scholar] [CrossRef]
- Fan, L.; Chen, Q.; Mairiyangu, Y.; Wang, Y.; Liu, X. Stable vesicle self-assembled from phospholipid and mannosylerythritol lipid and its application in encapsulating anthocyanins. Food Chem. 2020, 344, 128649. [Google Scholar] [CrossRef]
- Haque, F.; Sajid, M.; Cameotra, S.S.; Battacharyya, M.S. Anti-biofilm activity of a sophorolipid-amphotericin B niosomal formulation against Candida albicans. Biofouling 2017, 33, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Park, C.; Jang, H.J.; Kim, B.; Bae, H.W.; Chung, I.Y.; Kim, E.S.; Cho, Y.H. Antibacterial strategies inspired by the oxidative stress and response networks. J. Microbiol. 2019, 57, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Surekha, K.; Satputea, A.G.B.; Ibrahim, M.; Banatb, J.N.; Sangshettic, R.H.P.; Gaded, A.W.N. Multiple roles of biosurfactants in biofilms. Curr. Pharm. Des. 2016, 22, 1429–1448. [Google Scholar]
- Juma, A.; Lemoine, P.; Simpson, A.B.J.; Murray, J.; O’Hagan, B.M.G.; Naughton, P.J.; Dooley, J.G.; Banat, I.M. Microscopic investigation of the combined use of antibiotics and biosurfactants on methicillin resistant Staphylococcus aureus. Front. Microbiol. 2020, 11, 1477. [Google Scholar] [CrossRef]
- Joshi-Navare, K.; Prabhune, A. A biosurfactant-sophorolipid acts in synergy with antibiotics to enhance their efficiency. Biomed. Res. Int. 2013, 2013, 512495. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Producing Microorganism | Target Microorganism | Microbial Type | Inhibitory Effects | Reference |
|---|---|---|---|---|
| Pseudomonas aeruginosa PA1 | Listeria monocytogenes | Gram-positive | Anti-biofilm effects | [30] |
| Pseudomonas fluorescens | Gram-negative | |||
| Pseudomonas aeruginosa OBP1 | Staphylococcus aureus | Gram-positive | Antibacterial effects | [27] |
| Klebsiella pneumoniae | Gram-negative | |||
| P. aeruginosa DS10-129 | Staphylococcus epidermidis | Gram-positive | Anti-biofilm effects by disrupting the initial adhesion | [39] |
| Staphylococcus aureus | ||||
| Streptococcus salivarius | ||||
| Candida tropicalis | Fungi | |||
| Pseudomonas sp. PS-17 | Bacillus subtilis | Gram-positive | Antibacterial effects | [40,41] |
| — | Bacillus cereus | Gram-positive | Antibacterial effect depending on pH | [28] |
| Escherichia coli | Gram-negative | |||
| Salmonella enterica | Gram-negative | |||
| Pseudomonas aeruginosa MN1 | Streptococcus mutans | Gram-positive | Anti-biofilm effects | [31] |
| — | Helicobacter pylori | Gram-negative | Anti-biofilm effects | [42] |
| Pseudomonas aeruginosa strain B5 | Cercospora kikuchii | Fungi | Inhibiting spore germination and hyphal growth | [34] |
| Phytophthora capsici | ||||
| — | Yarrowia lipolytica | Fungi | Anti-biofilm effects | [38] |
| Pseudomonas aeruginosa DSVP20 | Candida albicans | Fungi | Anti-biofilm effects | [37] |
| Pseudomonas aeruginosa DS9 | Colletotrichum falcatum | Fungi | Inhibiting spore germination and mycelial growth | [35] |
| Pseudomonas aeruginosa SS14 | Trichophyton rubrum | Fungi | Inhibiting spore germination and hyphal proliferation | [36] |
| Producing Microorganism | Target Microorganism | Microbial Type | Inhibitory Effects | Reference |
|---|---|---|---|---|
| Candida glabrata CBS138 | Bacillus subtilis | Gram-positive | Antibacterial effects | [53] |
| Escherichia coli | Gram-negative | |||
| Candida tropicalis RA1 | Staphylococcus aureus | Gram-positive | Antibacterial effects | [51] |
| Listeria monocytogenes | ||||
| Escherichia coli | Gram-negative | |||
| Candida bombicola | Enterococcus faecium | Gram-positive | Antibacterial effects | [52] |
| Staphylococcus aureus | ||||
| Streptococcus mutans | ||||
| Proteus mirabilis, | Gram-negative | |||
| Escherichia coli | ||||
| Salmonella enterica subsp. enterica | ||||
| Candida bombicola ATCC 22214 | Lactobacillus acidophilus | Gram-positive | Antibacterial effects | [54] |
| Lactobacillus fermentum | ||||
| Streptococcus mutans | ||||
| Streptococcus salivarius | ||||
| Streptococcus sobrinus | ||||
| Starmerella (Candida) bombicola | Clostridium perfringens | Gram-positive | Antibacterial effects | [55] |
| Campylobacter jejuni | Gram-negative | |||
| Candida bombicola | Escherichia coli | Gram-negative | Antibacterial effects | [61] |
| Candida bombicola ATCC 22214 | Staphylococcus aureus | Gram-positive | Anti-biofilm effects | [56] |
| Pseudomonas aeruginosa | Gram-negative | |||
| Candida albicans | Fungi | |||
| Starmerella (Candida) bombicola MTCC1910 | Candida albicans | Fungi | Antifungal and anti-biofilm effects | [58,59] |
| Rhodotorula babjevae YS3 | Colletotrichum gloeosporioides | Fungi | Antifungal effects | [57] |
| Fusarium verticilliodes | ||||
| Fusarium oxysporum f. sp. pisi | ||||
| Corynespora cassiicola | ||||
| Trichophyton rubrum | ||||
| Wickerhamiella domercqiaeY2A | Phytophthora infestans | Fungi | Inhibiting spore germination and mycelial growth | [60] |
| Fusarium sp. | ||||
| Fusarium concentricum | ||||
| Fusarium oxysporum | ||||
| Pythium ultimum | ||||
| Pyricularia oryzae | ||||
| Rhizoctorzia solani | ||||
| Alternaria kikuchiana | ||||
| Gaeumannomyces graminis var. tritici | ||||
| Phytophthora infestans | ||||
| Rhodotorula babjevae YS3 | Trichophyton mentagrophytes | Fungi | Antifungal and anti-biofilm effects | [62] |
| Producing Microorganism | Target Microorganism | Microbial Type | Inhibitory Effects | Reference |
|---|---|---|---|---|
| Pseudozyma aphidis T-34 | Micrococcus luteus | Gram-positive | Antibacterial effects | [72] |
| Pseudozyma aphidis CBS 517.83 | Bacillus megaterium | Gram-positive | Antibacterial effects | [73] |
| Bacillus subtilis | ||||
| Staphylococcus aureus | ||||
| Candida magnoliae | Fungi | |||
| Pseudozyma aphidis | Bacillus cereus | Gram-positive | Antibacterial effects | [76] |
| Pseudozyma aphidis DSM 70725 | Staphylococcus aureus | Gram-positive | Antibacterial and anti-biofilm effects | [77] |
| Pseudozyma aphidis DSM 70,725 | Listeria monocytogenes | Gram-positive | Antibacterial effects | [78] |
| Micrococcus luteus | Gram-positive | Antibacterial effects | [75] | |
| Staphylococcus aureus | ||||
| Enterococci faecalis | ||||
| Enterococci faecium | ||||
| Pseudozyma aphidis NBRC 10182 | Streptococcus bovis, | Gram-positive | Antibacterial effects | [74] |
| Lactobacillus ruminis | ||||
| Eubacterium ruminantium | ||||
| Butyrivibirio fibrisolvens | ||||
| Ruminococcus albus | ||||
| Ruminococcus flavefaciens | ||||
| Lactobacillus casei | Staphylococcus aureus | Gram-positive | Anti-biofilm effects | [80] |
| Pseudozyma yeast | Blumeria graminis f. sp. tritici | Fungi | Inhibition of conidial germination. | [79] |
| Colletotrichum dematium | ||||
| Glomerella cingulata | ||||
| Magnaporthe grisea |
| Name | Glycolipid Type | Antimicrobial Effects | Reference |
|---|---|---|---|
| Rhamnolipid-stabilized carvacrol-loaded zein nanoparticles | Rhamnolipid | Antibacterial and antifungal activity against Pseudomonas syringae and Fusarium oxysporum. | [100] |
| Silver nanoparticles | Rhamnolipid | Antibacterial and antifungal activity against Staphylococcus aureus, Bacillus subtilis, Escherichia coli and Klebsiella pneumoniae, Aspergillus niger, and Aspergillus flavus. | [98] |
| AgNPs | Rhamnolipid | Antibacterial activity against gram-positive bacteria and antifungal activity against phytopathogens. | [99] |
| Silver and iron oxide nanoparticles | Rhamnolipid | Antibacterial and anti-adhesive properties against biofilms formed by Pseudomonas aeruginosa and Staphylococcus aureus. | [97] |
| Chitosan/rhamnolipid nanoparticles | Rhamnolipid | Antibacterial and anti-biofilm activity against Staphylococcus strains. | [101] |
| ZnO Nanoparticle | Sophorolipid | Antibacterial activity against Salmonella enterica and Candida albicans. | [96] |
| Curcumin-sophorolipid nanostructures | Sophorolipid | Antibacterial activity anti-biofilm activity against Pseudomonas aeruginosa. | [105] |
| Gold Nanoparticles | Sophorolipid | Antibacterial and anti-biofilm effects on Staphylococcus aureus and Vibrio cholerae | [102] |
| Chitosan Nanoparticles | MEL-A | Antibacterial activity against Staphylococcus aureus | [103] |
| MEL@AgNPs, MEL@ZnONPs and Ag-ZnO/MEL/GA | MEL-A | Antibacterial activity against Escherichia coli, Salmonella enterica, Bacillus cereus and Staphylococcus aureus. | [106] |
| Gold nanoparticles (AuNPs) | MEL-A | Antibacterial activity against gram-positive and gram-negative bacteria. | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, Q.; Lou, H.; Wei, T.; Liu, X.; Chen, Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics 2021, 13, 227. https://doi.org/10.3390/pharmaceutics13020227
Shu Q, Lou H, Wei T, Liu X, Chen Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics. 2021; 13(2):227. https://doi.org/10.3390/pharmaceutics13020227
Chicago/Turabian StyleShu, Qin, Hanghang Lou, Tianyu Wei, Xiayu Liu, and Qihe Chen. 2021. "Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review" Pharmaceutics 13, no. 2: 227. https://doi.org/10.3390/pharmaceutics13020227