
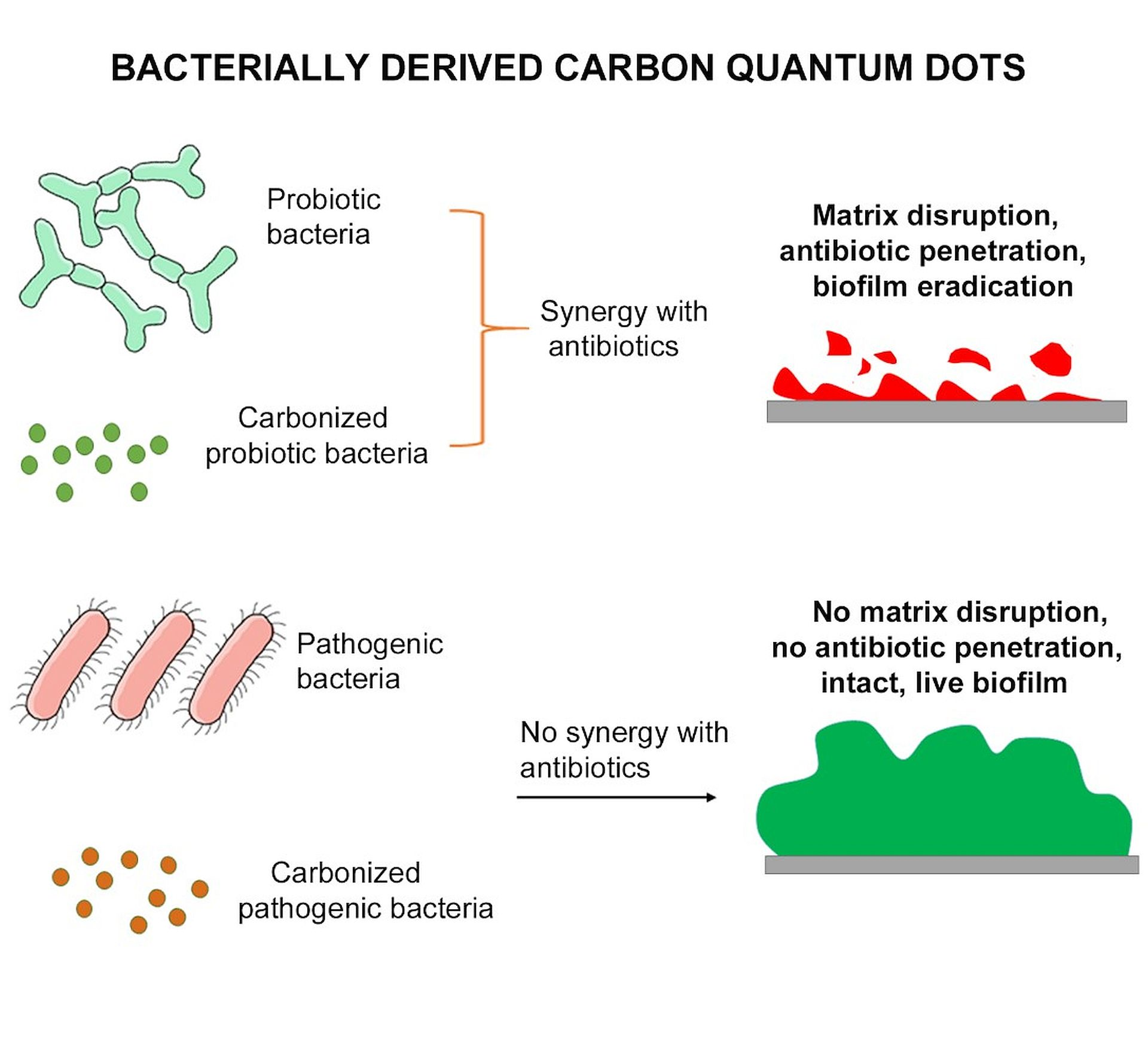
Synergy between “Probiotic” Carbon Quantum Dots and Ciprofloxacin in Eradicating Infectious Biofilms and Their Biosafety in Mice

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacteria Growth Conditions and Harvesting
2.2. Preparation and Characterization of Carbon Quantum Dots
2.3. Minimum Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC) of Carbon Quantum Dots
2.4. Biofilm Exposure to B-C- and E-C-Dots
2.5. Combined Effects of Pre-Exposure of Biofilms to Carbon Quantum Dots Followed by Antibiotics
2.6. Biosafety Assay in Mice
2.7. Statistical Analyses
3. Results and Discussion
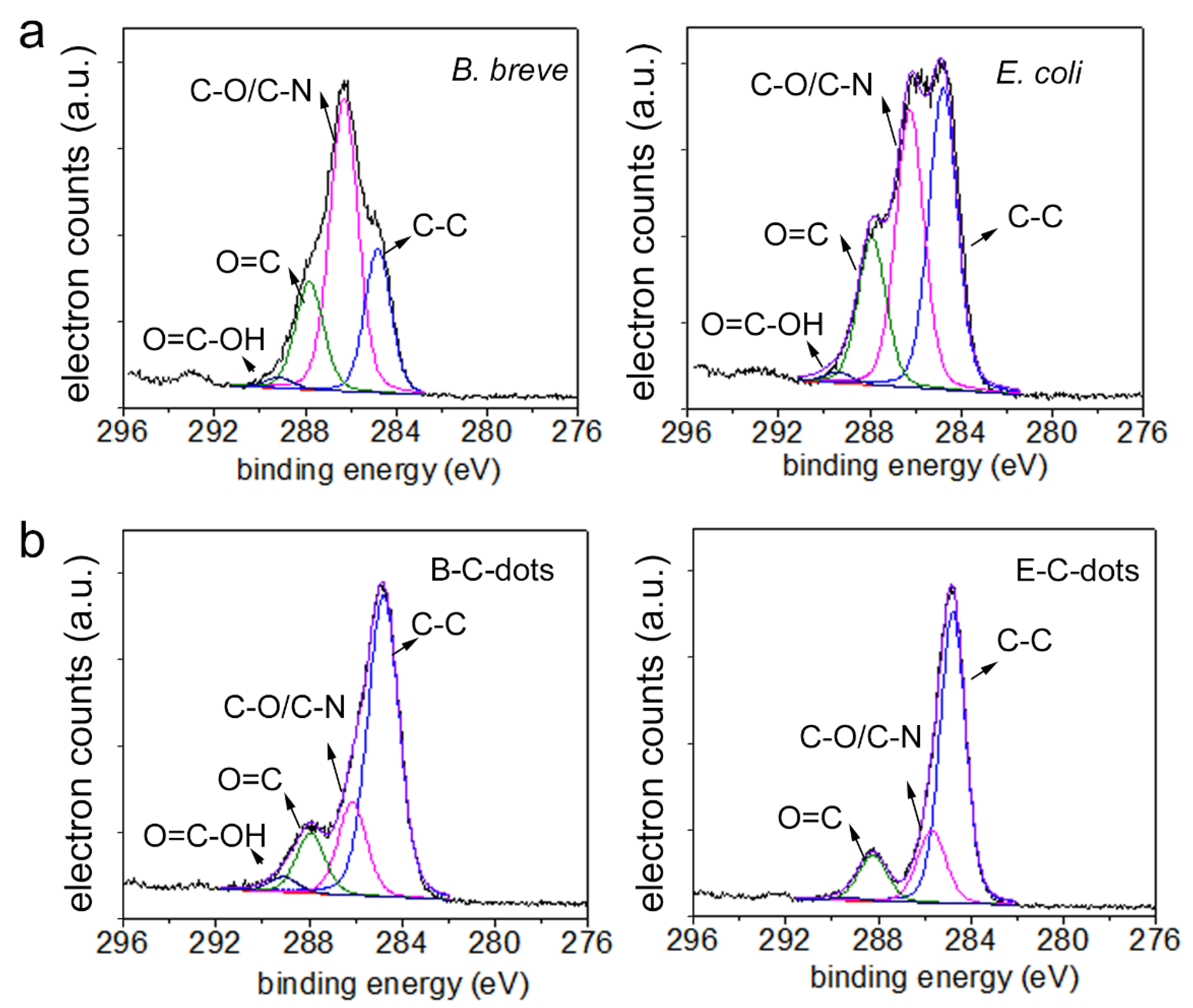
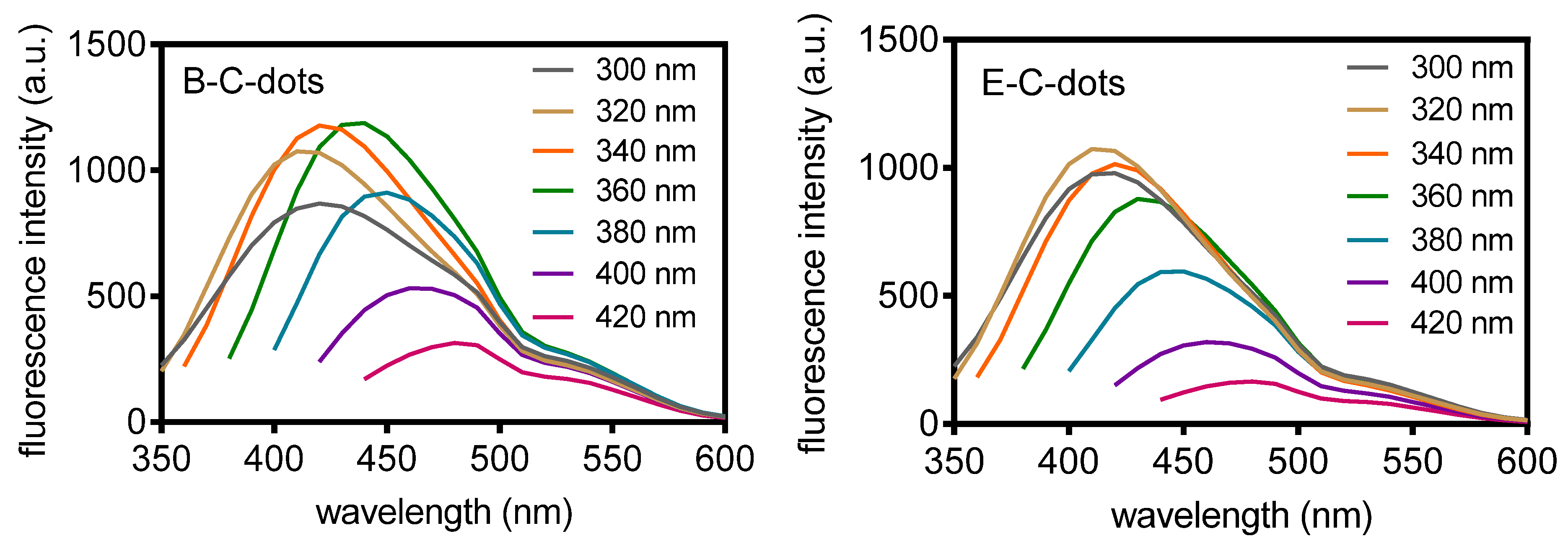
3.1. Physico-Chemical Characterization of Hydrothermally Derived C-Dots and Their Source Bacteria
3.2. Antibacterial Activity of Hydrothermally Derived Probiotic and Pathogenic C-Dots
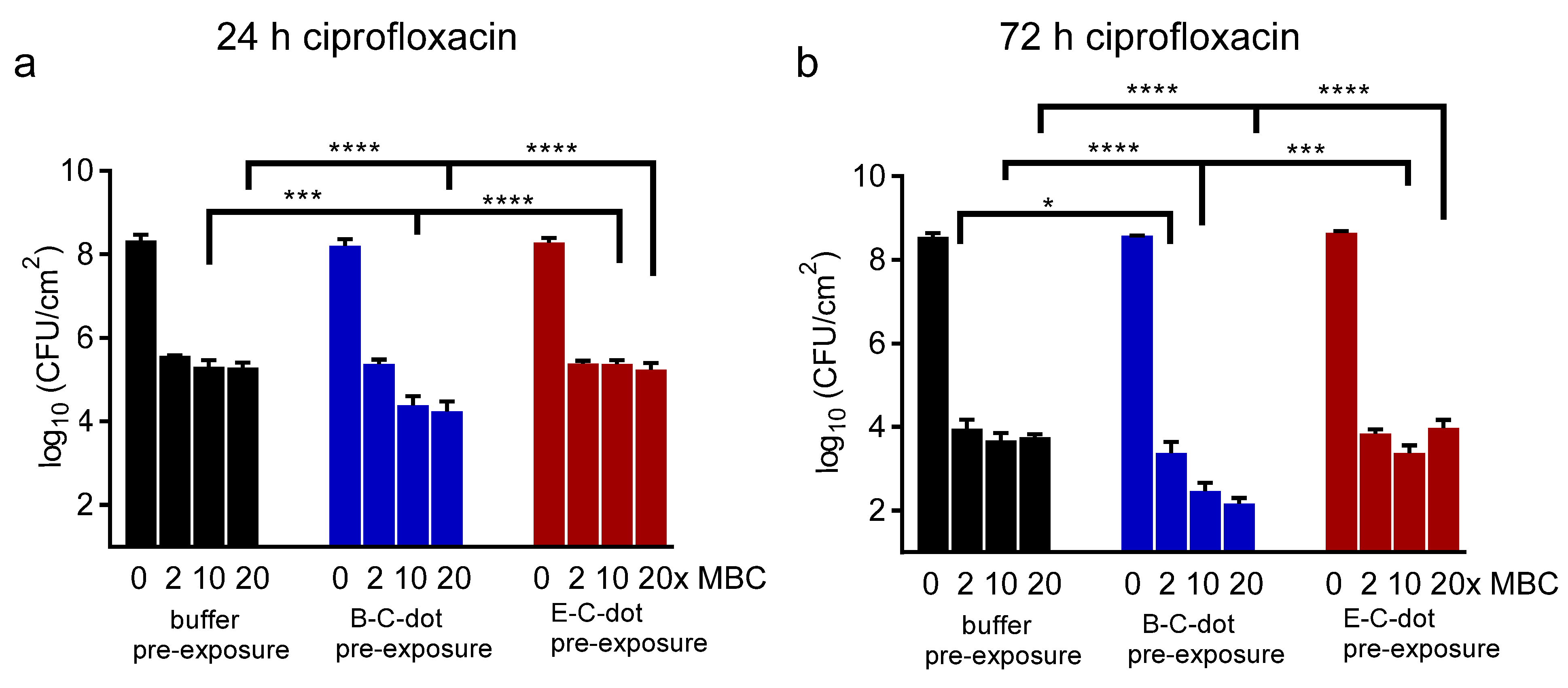
3.3. Synergy between Probiotic or Pathogenic C-Dots and Antibiotics in Eradicating E. coli Biofilm
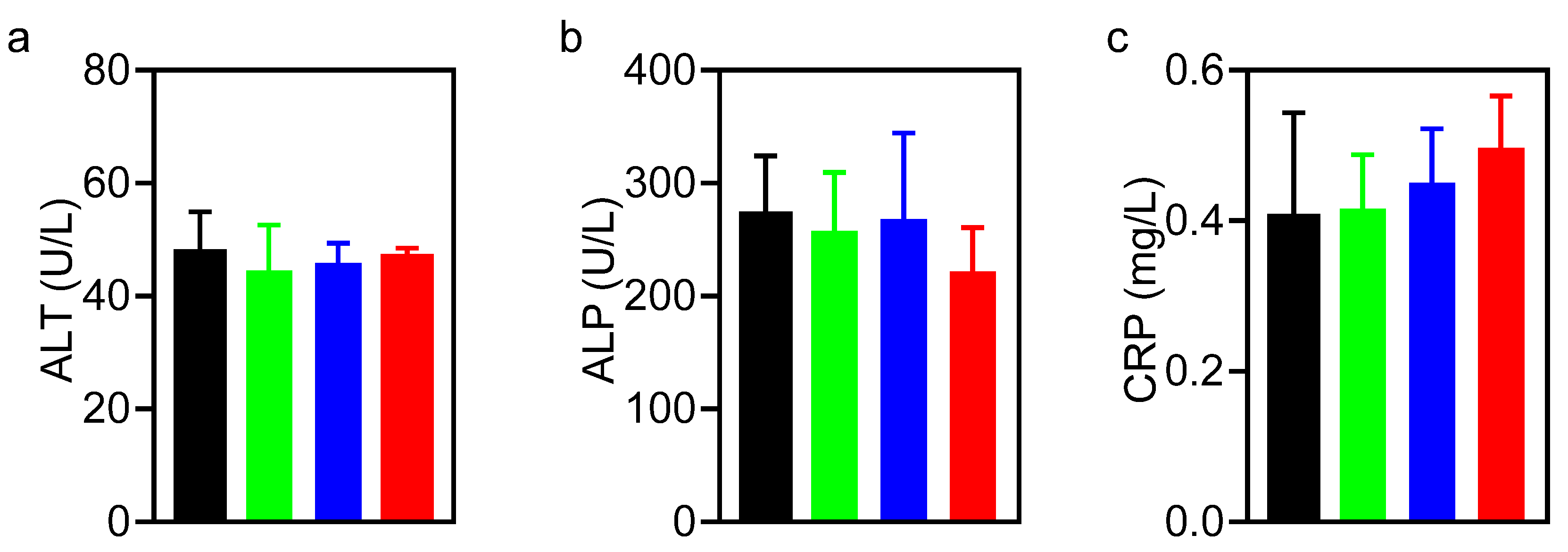
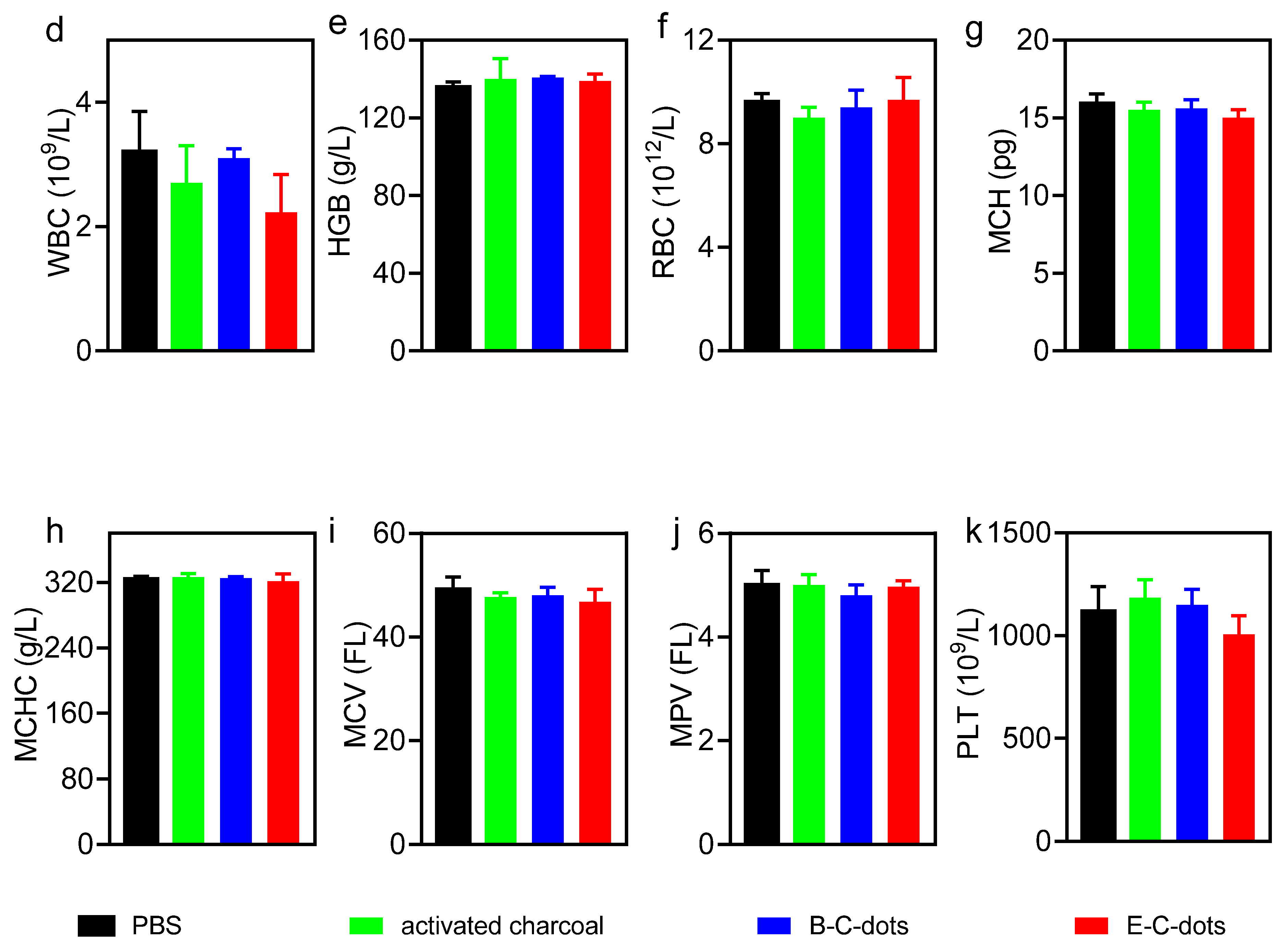
3.4. Biosafety of Hydrothermally Derived Probiotic and Pathogenic C-Dots in Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, D. Understanding Biofilm Resistance to Antibacterial Agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S. Mechanisms of Antibiotic Resistance in Bacterial Biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Leahy, S.C.; Higgins, D.G.; Fitzgerald, G.F.; Van Sinderen, D. Getting Better with Bifidobacteria. J. Appl. Microbiol. 2005, 98, 1303–1315. [Google Scholar] [CrossRef]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review Article: Bifidobacteria as Probiotic Agents—Physiological Effects and Clinical Benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In Vitro Evaluation of the Antimicrobial Activity of a Range of Probiotics against Pathogens: Evidence for the Effects of Organic Acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef]
- Yang, J.; Yang, H. Antibacterial Activity of Bifidobacterium breve Against Clostridioides difficile. Front. Cell. Infect. Microbiol. 2019, 9, 288. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.W.; Myers, L.E.S.; Ray, L.; Song, S.-C.; Nasr, T.R.; Berardinelli, A.J.; Kundu, K.; Murthy, N.; Hansen, J.M.; Neish, A.S. Lactobacillus rhamnosus Blocks Inflammatory Signaling In Vivo via Reactive Oxygen Species Generation. Free Radic. Biol. Med. 2009, 47, 1205–1211. [Google Scholar] [CrossRef] [Green Version]
- Dodoo, C.C.; Wang, J.; Basit, A.W.; Stapleton, P.; Gaisford, S. Targeted Delivery of Probiotics to Enhance Gastrointestinal Stability and Intestinal Colonisation. Int. J. Pharm. 2017, 530, 224–229. [Google Scholar] [CrossRef]
- Cook, M.T.; Tzortzis, G.; Khutoryanskiy, V.V.; Charalampopoulos, D. Layer-by-Layer Coating of Alginate Matrices with Chitosan–Alginate for the Improved Survival and Targeted Delivery of Probiotic Bacteria after Oral Administration. J. Mater. Chem. B 2013, 1, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.; Heinsen, F.-A.; Koenen, M.E.; Venema, K.; Knecht, H.; Hellmig, S.; Schreiber, S.; Ott, S.J. Effects of Probiotics and Antibiotics on the Intestinal Homeostasis in a Computer Controlled Model of the Large Intestine. BMC Microbiol. 2012, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Molaei, M.J. Carbon Quantum Dots and Their Biomedical and Therapeutic Applications: A Review. RSC Adv. 2019, 9, 6460–6481. [Google Scholar] [CrossRef]
- Zheng, W.; Zhou, Q.; Yuan, C. Nanoparticles for Oral Cancer Diagnosis and Therapy. Bioinorg. Chem. Appl. 2021, 2021, 9977131. [Google Scholar] [CrossRef]
- Jian, H.J.; Wu, R.S.; Lin, T.Y.; Li, Y.J.; Lin, H.J.; Harroun, S.G.; Lai, J.Y.; Huang, C.C. Super-Cationic Carbon Quantum Dots Synthesized from Spermidine as an Eye Drop Formulation for Topical Treatment of Bacterial Keratitis. ACS Nano 2017, 11, 6703–6716. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, S.; Cao, W.; Zhang, G.; Yang, X.; Gong, X.; Xing, X. Low-Toxicity Carbon Quantum Dots Derived from Gentamicin Sulfate to Combat Antibiotic Resistance and Eradicate Mature Biofilms. Chem. Commun. 2020, 56, 2316–2319. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, J.; Song, Y.; Zhang, M.; Wang, H.; Lu, F.; Huang, H.; Liu, Y.; Dai, X.; Gu, Z.; et al. Degradable Carbon Dots with Broad-Spectrum Antibacterial Activity. ACS Appl. Mater. Interfaces 2018, 10, 26936–26946. [Google Scholar] [CrossRef]
- Al Awak, M.M.; Wang, P.; Wang, S.; Tang, Y.; Sun, Y.P.; Yang, L. Correlation of Carbon Dots’ Light-Activated Antimicrobial Activities and Fluorescence Quantum Yield. RSC Adv. 2017, 7, 30177–30184. [Google Scholar] [CrossRef]
- Song, Y.; Lu, F.; Li, H.; Wang, H.; Zhang, M.; Liu, Y.; Kang, Z. Degradable Carbon Dots from Cigarette Smoking with Broad-Spectrum Antimicrobial Activities against Drug-Resistant Bacteria. ACS Appl. Bio Mater. 2018, 1, 1871–1879. [Google Scholar] [CrossRef]
- Wu, Y.; Van der Mei, H.C.; Busscher, H.J.; Ren, Y. Enhanced Bacterial Killing by Vancomycin in Staphylococcal Biofilms Disrupted by Novel, DMMA-Modified Carbon Dots Depends on EPS Production. Colloids Surf. B. Biointerfaces 2020, 193, 111114. [Google Scholar] [CrossRef]
- Lin, F.; Li, C.; Chen, Z. Bacteria-Derived Carbon Dots Inhibit Biofilm Formation of Escherichia coli without Affecting Cell Growth. Front. Microbiol. 2018, 9, 259. [Google Scholar] [CrossRef]
- Hua, X.W.; Bao, Y.W.; Wang, H.Y.; Chen, Z.; Wu, F.G. Bacteria-Derived Fluorescent Carbon Dots for Microbial Live/Dead Differentiation. Nanoscale 2017, 9, 2150–2161. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Ding, Y.; Hua, J.; Tang, B.; Ji, X.; Zhang, Q.; Wei, Y.; Qin, K.; Li, B. Bacteria-Derived Fluorescent Carbon Dots for Highly Selective Detection of: P -Nitrophenol and Bioimaging. Analyst 2019, 144, 5497–5503. [Google Scholar] [CrossRef]
- Wu, Y.; Wei, H.; Van der Mei, H.C.; De Vries, J.; Busscher, H.J.; Ren, Y. Inheritance of Physico-chemical Properties and ROS Generation by Carbon Quantum Dots Derived from Pyrolytically Carbonized Bacterial Sources. Mater. Today Bio 2021, 12, 100151. [Google Scholar]
- Hou, P.; Yang, T.; Liu, H.; Li, Y.F.; Huang, C.Z. An Active Structure Preservation Method for Developing Functional Graphitic Carbon Dots as an Effective Antibacterial Agent and a Sensitive pH and Al(III) Nanosensor. Nanoscale 2017, 9, 17334–17341. [Google Scholar] [CrossRef]
- Wang, H.; Song, Z.; Gu, J.; Li, S.; Wu, Y.; Han, H. Nitrogen-Doped Carbon Quantum Dots for Preventing Biofilm Formation and Eradicating Drug-Resistant Bacteria Infection. ACS Biomater. Sci. Eng. 2019, 5, 4739–4749. [Google Scholar] [CrossRef]
- Harris, J.R.; Roos, C.; Djalali, R.; Rheingans, O.; Maskos, M.; Schmidt, M. Application of the Negative Staining Technique to both Aqueous and Organic Solvent Solutions of Polymer Particles. Micron 1999, 30, 289–298. [Google Scholar] [CrossRef]
- Van der Mei, H.C.; Noordmans, J.; Busscher, H.J. Molecular Surface Characterization of Oral Streptococci by Fourier Transform Infrared Spectroscopy. BBA Gen. Subj. 1989, 991, 395–398. [Google Scholar] [CrossRef]
- Amory, D.E.; Genet, M.J.; Rouxhet, P.G. Application of XPS to the Surface Analysis of Yeast Cells. Surf. Interface Anal. 1988, 11, 478–486. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, P.; Zhai, X.; Tian, F.; Li, W.; Yang, J.; Liu, Y.; Wang, H.; Wang, W.; Liu, W. Nano-Carrier for Gene Delivery and Bioimaging Based on Carbon Dots with PEI-Passivation Enhanced Fluorescence. Biomaterials 2012, 33, 3604–3613. [Google Scholar] [CrossRef]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersboll, B.K.; Molin, S. Quantification of Biofilm Structures by the Novel Computer Program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Naumann, D. Some Ultrastructural Information on Intact, Living Bacterial Cells and Related Cell-Wall Fragments as given by FTIR. Infrared Phys. 1984, 24, 233–238. [Google Scholar] [CrossRef]
- Stanković, N.K.; Bodik, M.; Šiffalovič, P.; Kotlar, M.; Mičušik, M.; Špitalsky, Z.; Danko, M.; Milivojević, D.D.; Kleinova, A.; Kubat, P.; et al. Antibacterial and Antibiofouling Properties of Light Triggered Fluorescent Hydrophobic Carbon Quantum Dots Langmuir-Blodgett Thin Films. ACS Sustain. Chem. Eng. 2018, 6, 4154–4163. [Google Scholar] [CrossRef]
- Long, P.; Feng, Y.; Cao, C.; Li, Y.; Han, J.; Li, S.; Peng, C.; Li, Z.; Feng, W. Self-Protective Room-Temperature Phosphorescence of Fluorine and Nitrogen Codoped Carbon Dots. Adv. Funct. Mater. 2018, 28, 1800791. [Google Scholar] [CrossRef]
- Dittrich, M.; Sibler, S. Cell Surface Groups of Two Picocyanobacteria Strains Studied by Zeta Potential Investigations, Potentiometric Titration, and Infrared Spectroscopy. J. Colloid Interface Sci. 2005, 286, 487–495. [Google Scholar] [CrossRef]
- Ojeda, J.J.; Romero-González, M.E.; Bachmann, R.T.; Edyvean, R.G.J.; Banwart, S.A. Characterization of the Cell Surface and Cell Wall Chemistry of Drinking Water Bacteria by Combining XPS, FTIR Spectroscopy, Modeling, and Potentiometric Titrations. Langmuir 2008, 24, 4032–4040. [Google Scholar] [CrossRef]
- Turesky, R.J. Formation and Biochemistry of Carcinogenic Heterocyclic Aromatic Amines in Cooked Meats. Toxicol. Lett. 2007, 168, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Van der Mei, H.C.; De Vries, J.; Busscher, H.J. X-Ray Photoelectron Spectroscopy for the Study of Microbial Cell Surfaces. Surf. Sci. Rep. 2000, 39, 1–24. [Google Scholar] [CrossRef]
- Rouxhet, P.G.; Genet, M.J. XPS Analysis of Bio-Organic Systems. Surf. Interface Anal. 2011, 43, 1453–1470. [Google Scholar] [CrossRef]
- Van der Mei, H.C.; Leonard, A.J.; Weerkamp, A.H.; Rouxhet, P.G.; Busscher, H.J. Surface Properties of Streptococcus salivarius HB and Nonfibrillar Mutants: Measurement of Zeta Potential and Elemental Composition with X-Ray Photoelectron Spectroscopy. J. Bacteriol. 1988, 170, 2462–2466. [Google Scholar] [CrossRef] [Green Version]
- Barman, M.K.; Jana, B.; Bhattacharyya, S.; Patra, A. Photophysical Properties of Doped Carbon Dots (N, P, and B) and Their Influence on Electron/Hole Transfer in Carbon Dots-Nickel (II) Phthalocyanine Conjugates. J. Phys. Chem. C 2014, 118, 20034–20041. [Google Scholar] [CrossRef]
- Nandi, S.; Ritenberg, M.; Jelinek, R. Bacterial Detection with Amphiphilic Carbon Dots. Analyst 2015, 140, 4232–4237. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Bond, A.E.; Pan, N.; Coleman, M.; Tang, Y.; Sun, Y.P.; Yang, L. Synergistic Photoactivated Antimicrobial Effects of Carbon Dots Combined with Dye Photosensitizers. Int. J. Nanomed. 2018, 13, 8025–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholikov, K.; Ilhom, S.; Sajjad, M.; Smith, M.E.; Monroe, J.D.; San, O.; Er, A.O. Improved Singlet Oxygen Generation and Antimicrobial Activity of Sulphur-Doped Graphene Quantum Dots Coupled with Methylene Blue for Photodynamic Therapy Applications. Photodiagnosis Photodyn. Ther. 2018, 24, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Li, W.; Liu, H.; Zhang, L.; Wang, H.; Liu, Z.; Ren, J.; Qu, X. Construction of Nanozyme-Hydrogel for Enhanced Capture and Elimination of Bacteria. Adv. Funct. Mater. 2019, 29, 1–10. [Google Scholar] [CrossRef]
- Zhang, X.; Bishop, P.L.; Kupferle, M.J. Measurement of Polysaccharides and Proteins in Biofilm Extracellular Polymers. Water Sci. Technol. 1998, 37, 345–348. [Google Scholar] [CrossRef]
- Milho, C.; Silva, M.D.; Alves, D.; Oliveira, H.; Sousa, C.; Pastrana, L.M.; Azeredo, J.; Sillankorva, S. Escherichia coli and Salmonella enteritidis Dual-Species Biofilms: Interspecies Interactions and Antibiofilm Efficacy of Phages. Sci. Rep. 2019, 9, 18183. [Google Scholar] [CrossRef] [PubMed]
- Strelko, V.; Kuts, V.; Thrower, P. On the Mechanism of Possible Influence of Heteroatoms of Nitrogen, Boron and Phosphorus in a Carbon Matrix on the Catalytic Activity of Carbons in Electron Transfer Reactions. Carbon N.Y. 2000, 38, 1499–1503. [Google Scholar] [CrossRef]
- Strelko, V.V.; Kartel, N.T.; Dukhno, I.N.; Kuts, V.S.; Clarkson, R.B.; Odintsov, B.M. Mechanism of Reductive Oxygen Adsorption on Active Carbons with Various Surface Chemistry. Surf. Sci. 2004, 548, 281–290. [Google Scholar] [CrossRef]
- Li, Y.; Feng, C.; Li, J.; Mu, Y.; Liu, Y.; Kong, M.; Cheng, X.; Chen, X. Construction of Multilayer Alginate Hydrogel Beads for Oral Delivery of Probiotics Cells. Int. J. Biol. Macromol. 2017, 105, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Behrens, A.M.; Ginat, N.; Tzeng, S.Y.; Lu, X.; Sivan, S.; Langer, R.; Jaklenec, A. Biofilm-Inspired Encapsulation of Probiotics for the Treatment of Complex Infections. Adv. Mater. 2018, 30, 1803925. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/ C-Dot | FTIR Absorption Band Ratios * | ||||
| 1660 cm−1/CH | 1540 cm−1/CH | 1590 cm−1/CH | (1260–1245 cm−1)/CH | (1200–950 cm−1)/CH | |
| B. breve | 3.4 ± 0.3 | 0.8 ± 0.1 | - | 0.7 ± 0.1 | 3.7 ± 0.2 |
| B-C-dots | 1.1 ± 0.5 | - | 3.2 ± 0.1 | 0.2 ± 0.1 | 5.3 ± 1.3 |
| E. coli | 4.9 ± 0.1 | 1.7 ± 0.0 | - | 0.8 ± 0.1 | 2.0 ± 0.1 |
| E-C-dots | 0.8 ± 0.1 | - | 1.4 ± 0.5 | 0.1 ± 0.0 | 2.2 ± 0.4 |
| XPS Elemental Surface Composition (at%) ** | |||||
| C1s | N1s | O1s | P2p | ||
| B. breve | 57.6 | 5.5 | 32.8 | 1.1 | |
| B-C-dots | 57.7 | 4.9 | 25.7 | 2.0 | |
| E. coli | 59.7 | 7.8 | 31.1 | 1.2 | |
| E-C-dots | 66.6 | 2.8 | 22.1 | 0.3 | |
| Chemical Functionalities Involving C (at%) *** | |||||
| C-C/C=C | C-N/C-O | O=C | -COOH | ||
| B. breve | 13 | 32 | 12 | 1 | |
| B-C-dots | 37 | 12 | 8 | 1 | |
| E. coli | 20 | 27 | 11 | 1 | |
| E-C-dots | 47 | 11 | 8 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Yang, G.; van der Mei, H.C.; Shi, L.; Busscher, H.J.; Ren, Y. Synergy between “Probiotic” Carbon Quantum Dots and Ciprofloxacin in Eradicating Infectious Biofilms and Their Biosafety in Mice. Pharmaceutics 2021, 13, 1809. https://doi.org/10.3390/pharmaceutics13111809
Wu Y, Yang G, van der Mei HC, Shi L, Busscher HJ, Ren Y. Synergy between “Probiotic” Carbon Quantum Dots and Ciprofloxacin in Eradicating Infectious Biofilms and Their Biosafety in Mice. Pharmaceutics. 2021; 13(11):1809. https://doi.org/10.3390/pharmaceutics13111809
Chicago/Turabian StyleWu, Yanyan, Guang Yang, Henny C. van der Mei, Linqi Shi, Henk J. Busscher, and Yijin Ren. 2021. "Synergy between “Probiotic” Carbon Quantum Dots and Ciprofloxacin in Eradicating Infectious Biofilms and Their Biosafety in Mice" Pharmaceutics 13, no. 11: 1809. https://doi.org/10.3390/pharmaceutics13111809