
Generation of a Biomimetic Substitute of the Corneal Limbus Using Decellularized Scaffolds

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining Decellularized Xenografts from Native Limbi
- −
- Protocol P1: Double-distilled water (ddH2O) for 24 h; 0.1% sodium dodecyl sulphate (SDS) (3 incubations of 24 h each).
- −
- Protocol P2: ddH2O for 24 h; 0.1% SDS for 24 h; wash in PBS; 1.5 M of NaCl (2 incubations of 24 h each).
- −
- Protocol P3: ddH2O for 24 h; 0.1% SDS for 24 h; wash in PBS; 1% sodium deoxycholate (SDC) for 24 h; wash in PBS; 0.6% triton X-100 for 24 h; wash in PBS; 100 mg/L of DNAse and 20 mg/L of RNAse for 45 min.
- −
- Protocol P4: ddH2O for 24 h; 0.1% SDS for 24 h; wash in PBS; 1% sodium deoxycholate (SDC) for 24 h; wash in PBS; 0.6% triton X-100 for 24 h; wash in PBS; 100 mg/L of DNAse and 20 mg/L of RNAse for 45 min; wash in PBS; 0.05% Trypsin for 1 h.
2.2. Evaluation of Decellularization Efficiency in DL
2.3. Generation of Recellularized Limbal Substitutes by Tissue Engineering
2.4. Histological Analyses of DL and RL
2.5. Quantitative Analysis and Statistics
3. Results
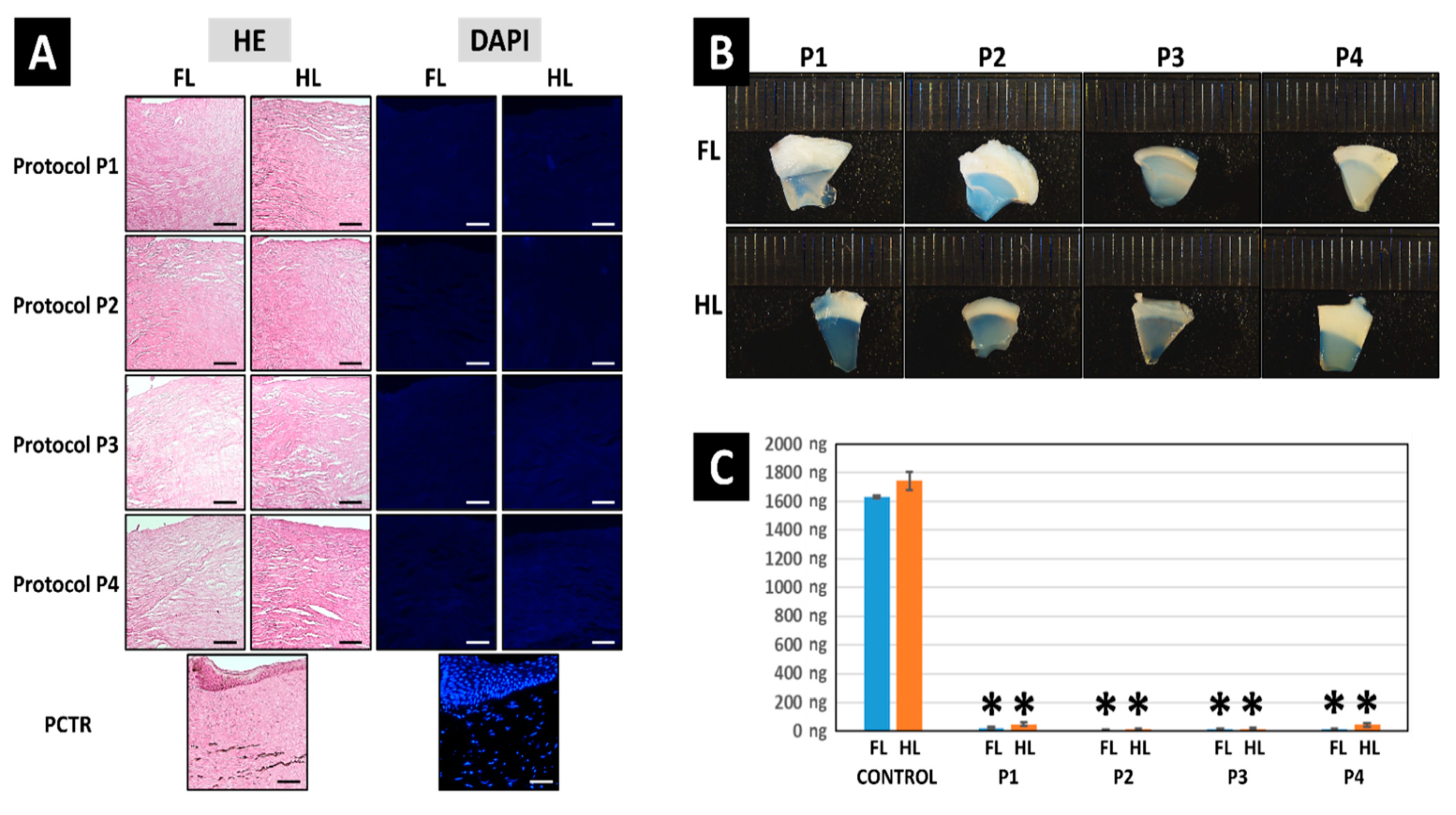
3.1. Decellularization Efficiency of the Different Protocols Applied to the Porcine Limbus
3.2. Histochemical Analysis of ECM Components Preservation in Decellularized Limbi
3.3. Histological Analysis of Recellularized Limbi
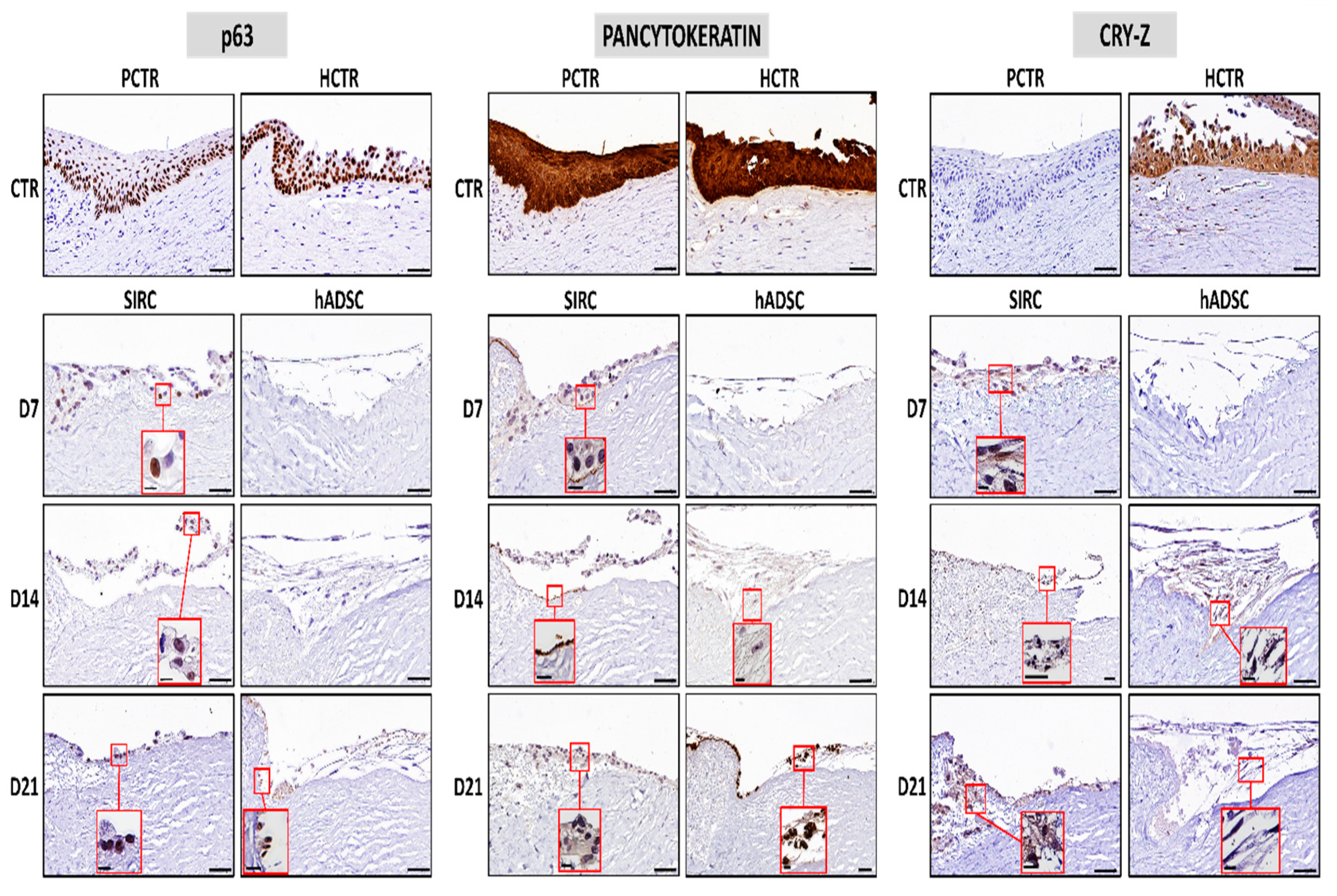
3.4. Evaluation of Limbal Cell Markers in Recellularized Limbi
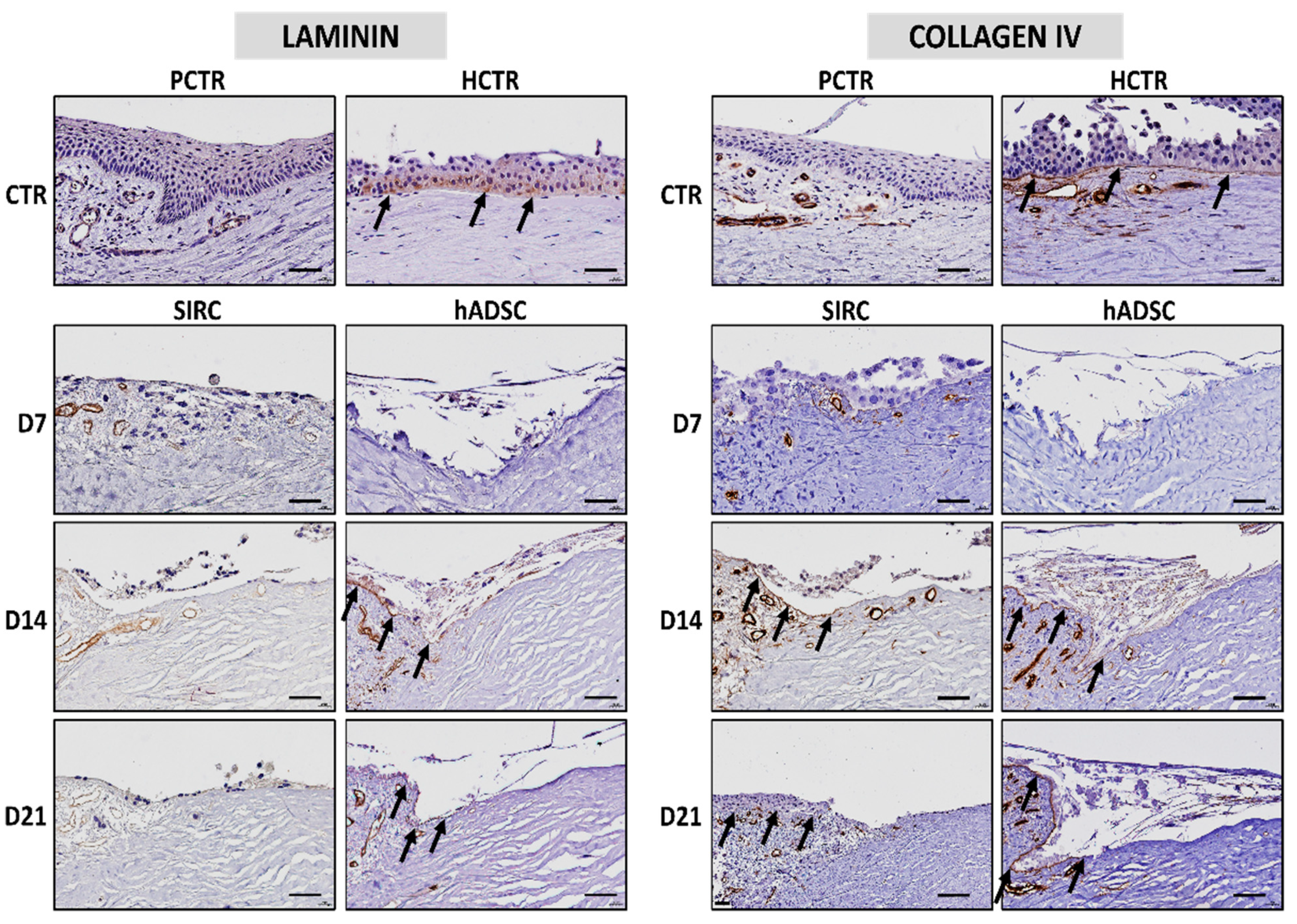
3.5. Immunohistochemical Analysis of Basement Membrane Components in Recellularized Limbi
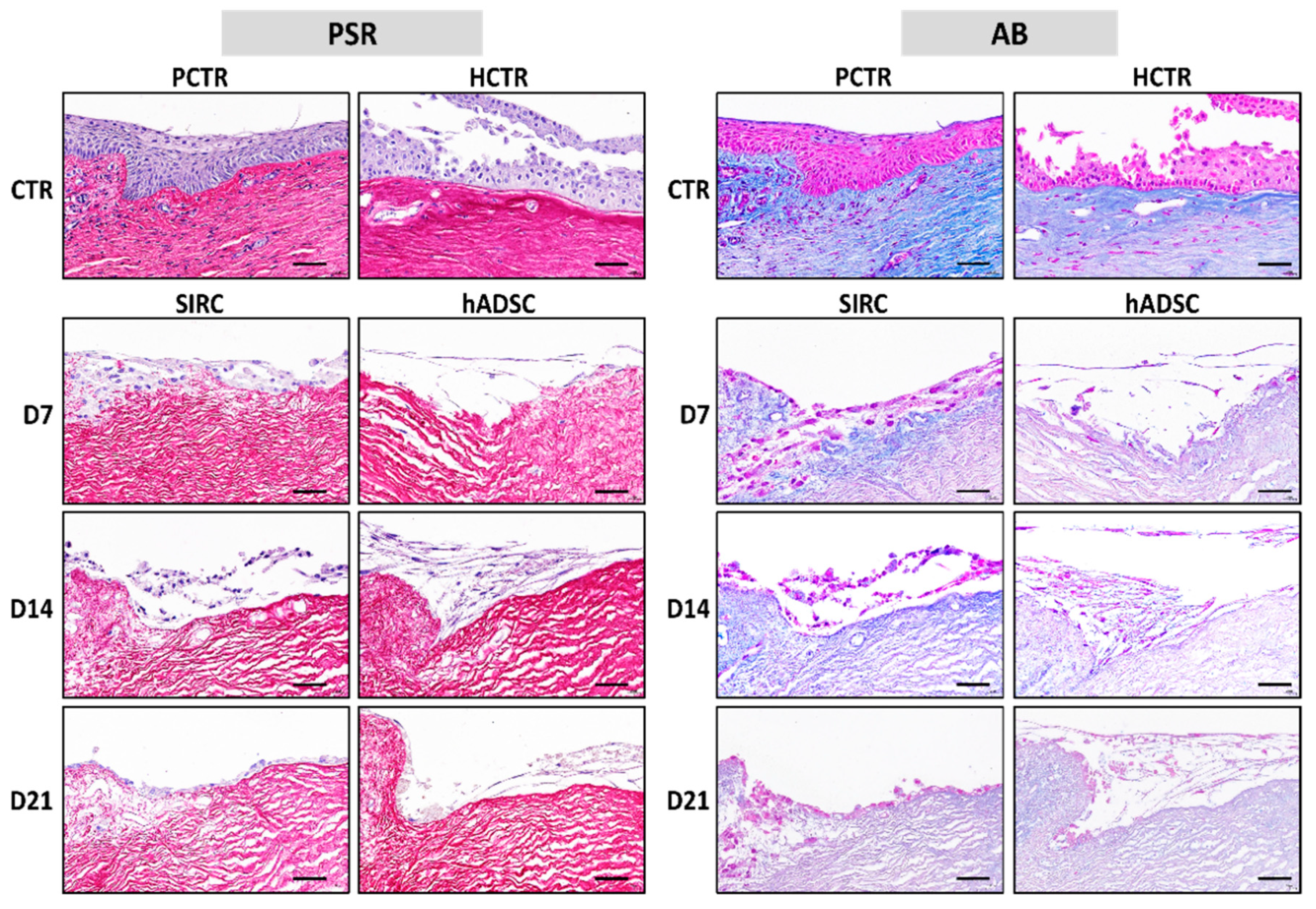
3.6. Histochemical Analysis of ECM Components in Recellularized Limbi
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jhanji, V.; Billig, I.; Yam, G.H.-F. Cell-Free Biological Approach for Corneal Stromal Wound Healing. Front. Pharmacol. 2021, 12, 671405. [Google Scholar] [CrossRef]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Bremond-Gignac, D.; Copin, H.; Benkhalifa, M. Corneal epithelial stem cells for corneal injury. Expert Opin. Biol. Ther. 2018, 18, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sangwan, V.S. Mini-Review: Regenerating the Corneal Epithelium With Simple Limbal Epithelial Transplantation. Front. Med. 2021, 8, 673330. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, F.; Glanville, J.; Arber, M.; Carr, E.; Rydevik, G.; Hogg, J.; Okonkwo, A.; Figueiredo, G.; Lako, M.; Whiter, F.; et al. A systematic review of cellular therapies for the treatment of limbal stem cell deficiency affecting one or both eyes. Ocul. Surf. 2021, 20, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.T.; Henry, J.J. Understanding cornea epithelial stem cells and stem cell deficiency: Lessons learned using vertebrate model systems. Genesis 2021, 59, e23411. [Google Scholar] [CrossRef] [PubMed]
- Miri, A.; Al-Deiri, B.; Dua, H.S. Long-term Outcomes of Autolimbal and Allolimbal Transplants. Ophthalmology 2010, 117, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal Stem-Cell Therapy and Long-Term Corneal Regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo, S.; de la Mata, A.; López-Paniagua, M.; Herreras, J.M.; Pérez, I.; Calonge, M.; Nieto-Miguel, T. Subconjunctival injection of mesenchymal stem cells for corneal failure due to limbal stem cell deficiency: State of the art. Stem Cell Res. Ther. 2021, 12, 60. [Google Scholar] [CrossRef]
- Deng, S.X.; Kruse, F.; Gomes, J.A.P.; Chan, C.C.; Daya, S.; Dana, R.; Figueiredo, F.C.; Kinoshita, S.; Rama, P.; Sangwan, V.; et al. Global Consensus on the Management of Limbal Stem Cell Deficiency. Cornea 2020, 39, 1291–1302. [Google Scholar] [CrossRef]
- Shafiq, M.A.; Milani, B.Y.; Djalilian, A.R. In Vivo Evaluation of a Decellularized Limbal Graft for Limbal Reconstruction. Int. J. Tissue Eng. 2014, 2014, e754245. [Google Scholar] [CrossRef]
- Nishida, K.; Yamato, M.; Hayashida, Y.; Watanabe, K.; Yamamoto, K.; Adachi, E.; Nagai, S.; Kikuchi, A.; Maeda, N.; Watanabe, H.; et al. Corneal Reconstruction with Tissue-Engineered Cell Sheets Composed of Autologous Oral Mucosal Epithelium. N. Engl. J. Med. 2004, 351, 1187–1196. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Herrera, J.E.; Ko, I.K.; Atala, A.; Yoo, J.J. Decellularization for whole organ bioengineering. Biomed. Mater. 2013, 8, 014106. [Google Scholar] [CrossRef]
- Ng, W.L.; Chua, C.K.; Shen, Y.-F. Print Me an Organ! Why We Are Not There Yet. Prog. Polym. Sci. 2019, 97, 101145. [Google Scholar] [CrossRef]
- Lee, J.M.; Ng, W.L.; Yeong, W.Y. Resolution and shape in bioprinting: Strategizing towards complex tissue and organ printing. Appl. Phys. Rev. 2019, 6, 011307. [Google Scholar] [CrossRef]
- Melchels, F.; Barradas, A.; van Blitterswijk, C.; de Boer, J.; Feijen, J.; Grijpma, D.W. Effects of the architecture of tissue engineering scaffolds on cell seeding and culturing. Acta Biomater. 2010, 6, 4208–4217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melchels, F.; Tonnarelli, B.; Olivares, A.L.; Martin, I.; Lacroix, D.; Feijen, J.; Wendt, D.J.; Grijpma, D.W. The influence of the scaffold design on the distribution of adhering cells after perfusion cell seeding. Biomaterials 2011, 32, 2878–2884. [Google Scholar] [CrossRef] [Green Version]
- Khosravimelal, S.; Mobaraki, M.; Eftekhari, S.; Ahearne, M.; Seifalian, A.M.; Gholipourmalekabadi, M. Hydrogels as Emerging Materials for Cornea Wound Healing. Small 2021, 17, e2006335. [Google Scholar] [CrossRef]
- Guérin, L.-P.; Le-Bel, G.; Desjardins, P.; Couture, C.; Gillard, E.; Boisselier, E.; Bazin, R.; Germain, L.; Guérin, S. The Human Tissue-Engineered Cornea (hTEC): Recent Progress. Int. J. Mol. Sci. 2021, 22, 1291. [Google Scholar] [CrossRef]
- Rico-Sánchez, L.; Garzón, I.; González-Andrades, M.; Ruíz-García, A.; Punzano, M.; Lizana-Moreno, A.; Muñoz-Ávila, J.I.; Sánchez-Quevedo, M.D.C.; Martínez-Atienza, J.; Lopez-Navas, L.; et al. Successful development and clinical translation of a novel anterior lamellar artificial cornea. J. Tissue Eng. Regen. Med. 2019, 13, 2142–2154. [Google Scholar] [CrossRef] [Green Version]
- Andrades, M.G.; Mata, R.; González-Gallardo, M.D.C.; Medialdea, S.; Arias-Santiago, S.; Martinez-Atienza, J.; Ruiz-García, A.; Pérez-Fajardo, L.; Lizana-Moreno, A.; Garzón, I.; et al. A study protocol for a multicentre randomised clinical trial evaluating the safety and feasibility of a bioengineered human allogeneic nanostructured anterior cornea in patients with advanced corneal trophic ulcers refractory to conventional treatment. BMJ Open 2017, 7, e016487. [Google Scholar] [CrossRef]
- Buznyk, O.; Pasyechnikova, N.; Islam, M.M.; Iakymenko, S.; Fagerholm, P.; Griffith, M. Bioengineered Corneas Grafted as Alternatives to Human Donor Corneas in Three High-Risk Patients. Clin. Transl. Sci. 2015, 8, 558–562. [Google Scholar] [CrossRef]
- Saldin, L.T.; Cramer, M.C.; Velankar, S.S.; White, L.J.; Badylak, S.F. Extracellular matrix hydrogels from decellularized tissues: Structure and function. Acta Biomater. 2017, 49, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.H.; Choi, H.J.; Kim, M.K. Corneal xenotransplantation: Where are we standing? Prog. Retin. Eye Res. 2021, 80, 100876. [Google Scholar] [CrossRef] [PubMed]
- Andrades, M.G.; Carriel, V.; Rivera-Izquierdo, M.; Garzón, I.; González-Andrades, E.; Medialdea, S.; Alaminos, M.; Campos, A. Effects of Detergent-Based Protocols on Decellularization of Corneas with Sclerocorneal Limbus. Evaluation of Regional Differences. Transl. Vis. Sci. Technol. 2015, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.; Du, L.; Wu, X. A rabbit anterior cornea replacement derived from acellular porcine cornea matrix, epithelial cells and keratocytes. Biomaterials 2010, 31, 7257–7265. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, H.; Dai, Z.; Cao, Y.; Jin, C. Preparation and Biomechanical Properties of an Acellular Porcine Corneal Stroma. Cornea 2017, 36, 1343–1351. [Google Scholar] [CrossRef]
- Isidan, A.; Liu, S.; Chen, A.M.; Zhang, W.; Li, P.; Smith, L.J.; Hara, H.; Cooper, D.K.C.; Ekser, B. Comparison of porcine corneal decellularization methods and importance of preserving corneal limbus through decellularization. PLoS ONE 2021, 16, e0243682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-C.; Liu, X.; Jin, Y.; Jiang, D.-L.; Wei, X.-S.; Xie, H.-T. Lamellar Keratoplasty Treatment of Fungal Corneal Ulcers with Acellular Porcine Corneal Stroma. Arab. Archaeol. Epigr. 2015, 15, 1068–1075. [Google Scholar] [CrossRef]
- Spaniol, K.; Witt, J.; Mertsch, S.; Borrelli, M.; Geerling, G.; Schrader, S. Generation and characterisation of decellularised human corneal limbus. Graefe’s Arch. Clin. Exp. Ophthalmol. 2018, 256, 547–557. [Google Scholar] [CrossRef]
- Seyed-Safi, A.G.; Daniels, J.T. The limbus: Structure and function. Exp. Eye Res. 2020, 197, 108074. [Google Scholar] [CrossRef]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit In Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef]
- Galindo, S.; Herreras, J.M.; López-Paniagua, M.; Rey, E.; De La Mata, A.; Plata-Cordero, M.; Calonge, M.; Nieto-Miguel, T. Therapeutic Effect of Human Adipose Tissue-Derived Mesenchymal Stem Cells in Experimental Corneal Failure Due to Limbal Stem Cell Niche Damage. Stem Cells 2017, 35, 2160–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Coulson-Thomas, V.J.; Ferreira, T.G.; Kao, W.W.Y. Mesenchymal stem cells for treating ocular surface diseases. BMC Ophthalmol. 2015, 15, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crapo, P.M.; Gilbert, T.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philips, C.; Cornelissen, M.; Carriel, V. Evaluation methods as quality control in the generation of decellularized peripheral nerve allografts. J. Neural Eng. 2018, 15, 021003. [Google Scholar] [CrossRef]
- Mailey, B.; Hosseini, A.; Baker, J.; Young, A.; Alfonso, Z.; Hicok, K.; Wallace, A.M.; Cohen, S.R. Adipose-Derived Stem Cells: Methods for Isolation and Applications for Clinical Use. Methods Mol. Biol. 2014, 1210, 161–181. [Google Scholar] [CrossRef]
- El Soury, M.; García-García, D.; Moretti, M.; Perroteau, I.; Raimondo, S.; Lovati, A.; Carriel, V. Comparison of Decellularization Protocols to Generate Peripheral Nerve Grafts: A Study on Rat Sciatic Nerves. Int. J. Mol. Sci. 2021, 22, 2389. [Google Scholar] [CrossRef]
- Garzón, I.; Jaimes-Parra, B.; Pascual-Geler, M.; Cózar, J.; Sánchez-Quevedo, M.; Mosquera-Pacheco, M.; Sánchez-Montesinos, I.; Fernández-Valadés, R.; Campos, F.; Alaminos, M. Biofabrication of a Tubular Model of Human Urothelial Mucosa Using Human Wharton Jelly Mesenchymal Stromal Cells. Polymers 2021, 13, 1568. [Google Scholar] [CrossRef]
- Carriel, V.; Campos, A.; Alaminos, M.; Raimondo, S.; Geuna, S. Staining Methods for Normal and Regenerative Myelin in the Nervous System. Methods Mol. Biol. 2017, 1560, 207–218. [Google Scholar] [CrossRef]
- Chato-Astrain, J.; Philips, C.; Campos, F.; Durand-Herrera, D.; García-García, O.D.; Roosens, A.; Alaminos, M.; Campos, A.; Carriel, V. Detergent-based decellularized peripheral nerve allografts: An in vivo preclinical study in the rat sciatic nerve injury model. J. Tissue Eng. Regen. Med. 2020, 14, 789–806. [Google Scholar] [CrossRef] [PubMed]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Mehta, G.; Keikhosravi, A.; Liu, Y.; Lemancik, S.; Ricke, W.; Weber, S.M.; Kao, W.J.; et al. Comparison of Picrosirius Red Staining With Second Harmonic Generation Imaging for the Quantification of Clinically Relevant Collagen Fiber Features in Histopathology Samples. J. Histochem. Cytochem. 2016, 64, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oie, Y.; Komoto, S.; Kawasaki, R. Systematic review of clinical research on regenerative medicine for the cornea. Jpn. J. Ophthalmol. 2021, 65, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Tidu, A.; Schanne-Klein, M.-C.; Borderie, V.M. Development, structure, and bioengineering of the human corneal stroma: A review of collagen-based implants. Exp. Eye Res. 2020, 200, 108256. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Schrader, S.; Daniels, J.T. The Porcine Limbal Epithelial Stem Cell Niche as a New Model for the Study of Transplanted Tissue-Engineered Human Limbal Epithelial Cells. Tissue Eng. Part A 2011, 17, 741–750. [Google Scholar] [CrossRef]
- Wong, M.L.; Griffiths, L.G. Immunogenicity in xenogeneic scaffold generation: Antigen removal vs. decellularization. Acta Biomater. 2014, 10, 1806–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, J.; Disesa, V.; Gail, D.; Massicot-Fisher, J.; National Heart, Lung, and Blood Institute Heart and Lung Xenotransplantation Working Group. Recommendations of the National Heart, Lung, and Blood Institute Heart and Lung Xenotransplantation Working Group. Circulation 2002, 106, 1043–1047. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Li, N.; Wu, Z.; Wan, P.; Liang, X.; Zhang, W.; Wang, X.; Li, C.; Xiao, J.; Zhou, Q.; et al. Using acellular porcine limbal stroma for rabbit limbal stem cell microenvironment reconstruction. Biomaterials 2011, 32, 7812–7821. [Google Scholar] [CrossRef]
- Murab, S.; Ghosh, S. Impact of osmoregulatory agents on the recovery of collagen conformation in decellularized corneas. Biomed. Mater. 2016, 11, 065005. [Google Scholar] [CrossRef]
- Wang, M.; Cui, C.; Ibrahim, M.M.; Han, B.; Li, Q.; Pacifici, M.; Lawrence, J.T.R.; Han, L.; Han, L.-H. Regulating Mechanotransduction in Three Dimensions using Sub-Cellular Scale, Crosslinkable Fibers of Controlled Diameter, Stiffness, and Alignment. Adv. Funct. Mater. 2019, 29, 1808967. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, R.; Han, C.; Huang, L. Extracellular matrix grafts: From preparation to application (Review). Int. J. Mol. Med. 2021, 47, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Marin-Tapia, H.A.; Romero-Salazar, L.; Arteaga-Arcos, J.C.; Rosales-Ibáñez, R.; Mayorga-Rojas, M. Micro-mechanical properties of corneal scaffolds from two different bio-models obtained by an efficient chemical decellularization. J. Mech. Behav. Biomed. Mater. 2021, 119, 104510. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Miguel, T.; Galindo, S.; Reinoso, R.; Corell, A.; Martino, M.; Pérez-Simón, J.A.; Calonge, M. In VitroSimulation of Corneal Epithelium Microenvironment Induces a Corneal Epithelial-like Cell Phenotype from Human Adipose Tissue Mesenchymal Stem Cells. Curr. Eye Res. 2013, 38, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Garzon, I.; Chato-Astrain, J.; Campos, F.; Fernandez-Valades, R.; Sanchez-Montesinos, I.; Campos, A.; Alaminos, M.; D’Souza, R.N.; Martin-Piedra, M.A. Expanded Differentiation Capability of Human Wharton’s Jelly Stem Cells Toward Pluripotency: A Systematic Review. Tissue Eng. Part B Rev. 2020, 26, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Timoneda, M.A.; Levato, R.; Puñet, X.; Cano, I.; Castano, O.; Engel, E. Biofunctionalization of Polymeric Surfaces. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 1745–1748. [Google Scholar]
- Olivieri, M.; Cristaldi, M.; Pezzino, S.; Rusciano, D.; Tomasello, B.; Anfuso, C.D.; Lupo, G. Phenotypic characterization of the SIRC (Statens Seruminstitut Rabbit Cornea) cell line reveals a mixed epithelial and fibroblastic nature. Exp. Eye Res. 2018, 172, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, F.; Goh, T.-W.; Setiawan, M.; Yam, G.H.-F.; Mehta, J.S. Cellular therapy of corneal epithelial defect by adipose mesenchymal stem cell-derived epithelial progenitors. Stem Cell Res. Ther. 2020, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Martin-Piedra, M.; Alfonso-Rodriguez, C.; Zapater, A.; Durand-Herrera, D.; Chato-Astrain, J.; Campos, F.; Sanchez-Quevedo, M.; Alamino, M.; Garzon, I. Effective use of mesenchymal stem cells in human skin substitutes generated by tissue engineering. Eur. Cells Mater. 2019, 37, 233–249. [Google Scholar] [CrossRef]
- del Barrio, J.L.A.; Chiesa, M.; Garagorri, N.; Garcia-Urquia, N.; Fernandez-Delgado, J.; Bataille, L.; Rodriguez, A.; Arnalich-Montiel, F.; Zarnowski, T.; de Toledo, J.P.; et al. Acellular human corneal matrix sheets seeded with human adipose-derived mesenchymal stem cells integrate functionally in an experimental animal model. Exp. Eye Res. 2015, 132, 91–100. [Google Scholar] [CrossRef]
- Garzón, I.; Miyake, J.; González-Andrades, M.; Carmona, R.; Carda, C.; Sánchez-Quevedo, M.D.C.; Campos, A.; Alaminos, M. Wharton’s Jelly Stem Cells: A Novel Cell Source for Oral Mucosa and Skin Epithelia Regeneration. Stem Cells Transl. Med. 2013, 2, 625–632. [Google Scholar] [CrossRef]
- Tsai, R.J.-F.; Tsai, R.Y.-N. Ex Vivo Expansion of Corneal Stem Cells on Amniotic Membrane and Their Outcome. Eye Contact Lens 2010, 36, 305–309. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Porras, D.; Caro-Magdaleno, M.; González-Gallardo, C.; García-García, Ó.D.; Garzón, I.; Carriel, V.; Campos, F.; Alaminos, M. Generation of a Biomimetic Substitute of the Corneal Limbus Using Decellularized Scaffolds. Pharmaceutics 2021, 13, 1718. https://doi.org/10.3390/pharmaceutics13101718
Sánchez-Porras D, Caro-Magdaleno M, González-Gallardo C, García-García ÓD, Garzón I, Carriel V, Campos F, Alaminos M. Generation of a Biomimetic Substitute of the Corneal Limbus Using Decellularized Scaffolds. Pharmaceutics. 2021; 13(10):1718. https://doi.org/10.3390/pharmaceutics13101718
Chicago/Turabian StyleSánchez-Porras, David, Manuel Caro-Magdaleno, Carmen González-Gallardo, Óscar Darío García-García, Ingrid Garzón, Víctor Carriel, Fernando Campos, and Miguel Alaminos. 2021. "Generation of a Biomimetic Substitute of the Corneal Limbus Using Decellularized Scaffolds" Pharmaceutics 13, no. 10: 1718. https://doi.org/10.3390/pharmaceutics13101718