
Hyaluronic Acid-Glycine-Cholesterol Conjugate-Based Nanoemulsion as a Potent Vaccine Adjuvant for T Cell-Mediated Immunity

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
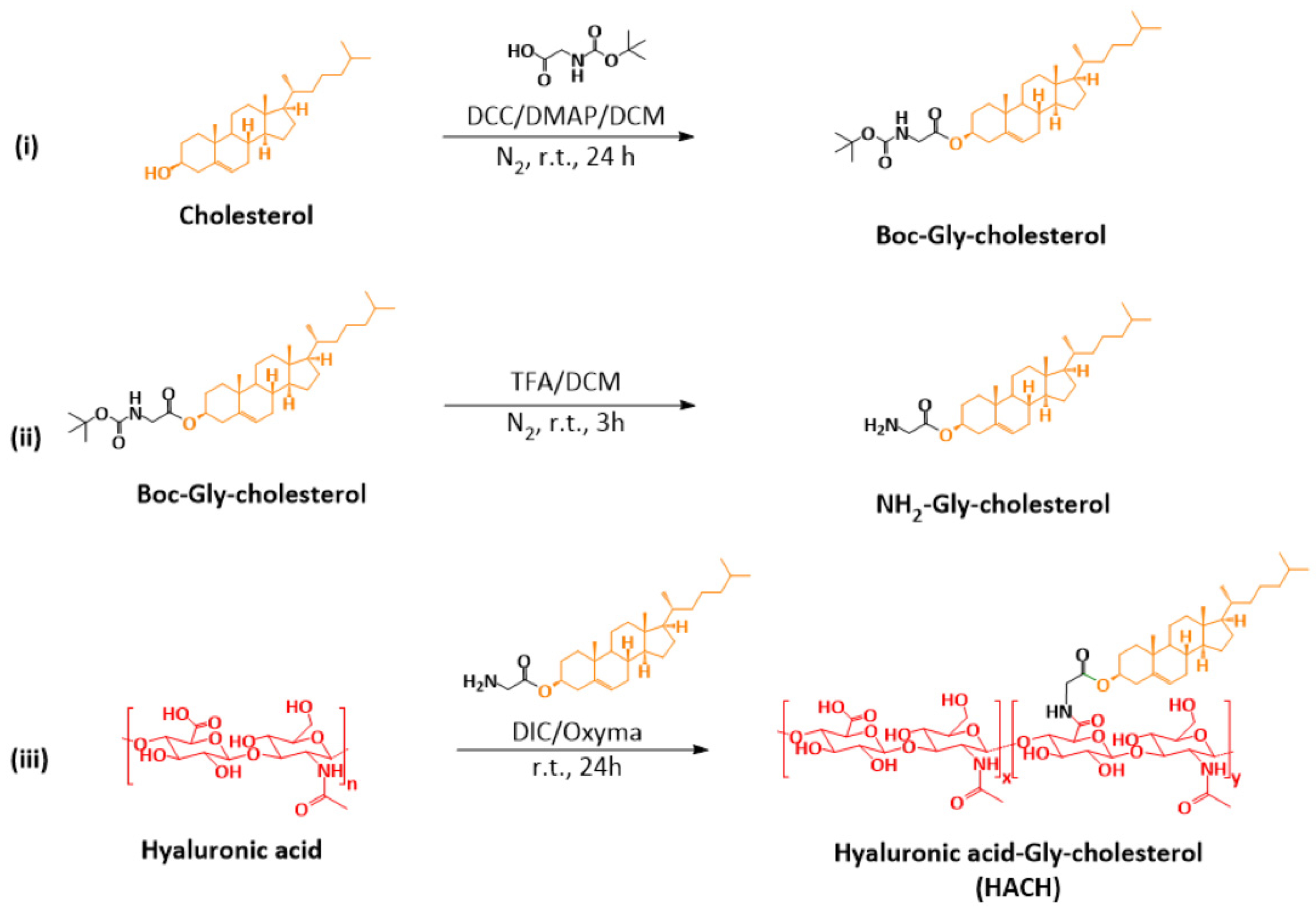
2.2. Synthesis and Characterization of Hyaluronic Acid-Gly-Cholesterol (HACH) Conjugates
2.2.1. Synthesis of Boc-Gly-Cholesterol
2.2.2. Deprotection of Boc-Gly-Cholesterol
2.2.3. Conjugation of HA and NH2-Gly-Cholesterol
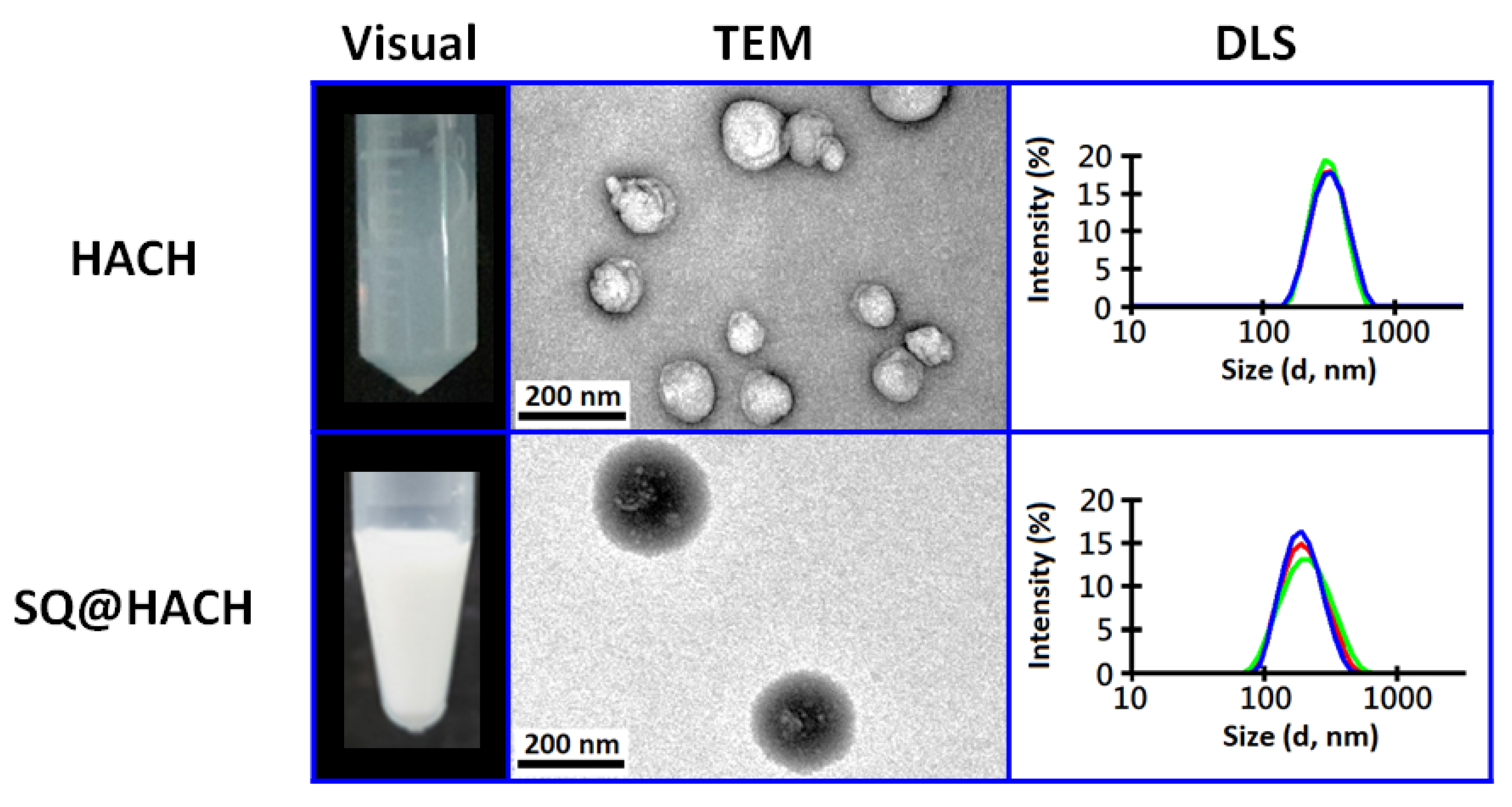
2.3. Preparation and Characterization of HACH-Stabilized Squalene Emulsion (SQ@HACH)
2.4. Animals and Ethics Statement
2.5. Immunization In Vivo and Tissue Sample Preparation
2.5.1. Immunization Schedule
2.5.2. Preparation of Muscular Tissues at the Injection Sites and Splenocyte Samples
2.5.3. Preparation of the Sera Samples
2.6. MRNA Expression of T Cells
2.7. Cytotoxic T Lymphocyte-Mediated Killing Activity
2.8. IgG and IgG Subtype Titers
2.9. Statistical Analysis
3. Results and Discussion
3.1. Synthesis and Characterization of HACH
3.2. Self-Assembling Properties of HACH without or with Squalene
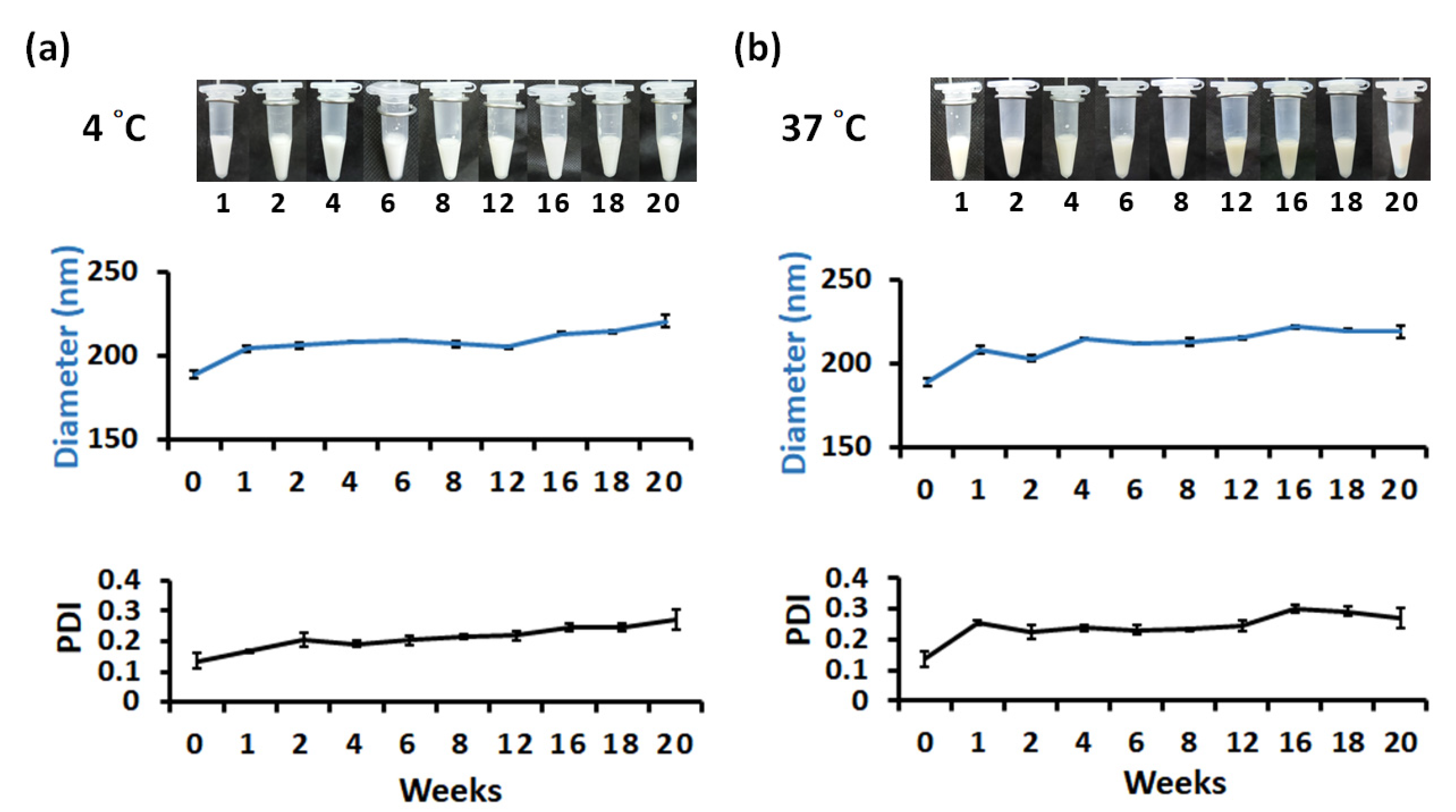
3.3. Long-Term Storage of SQ@HACH
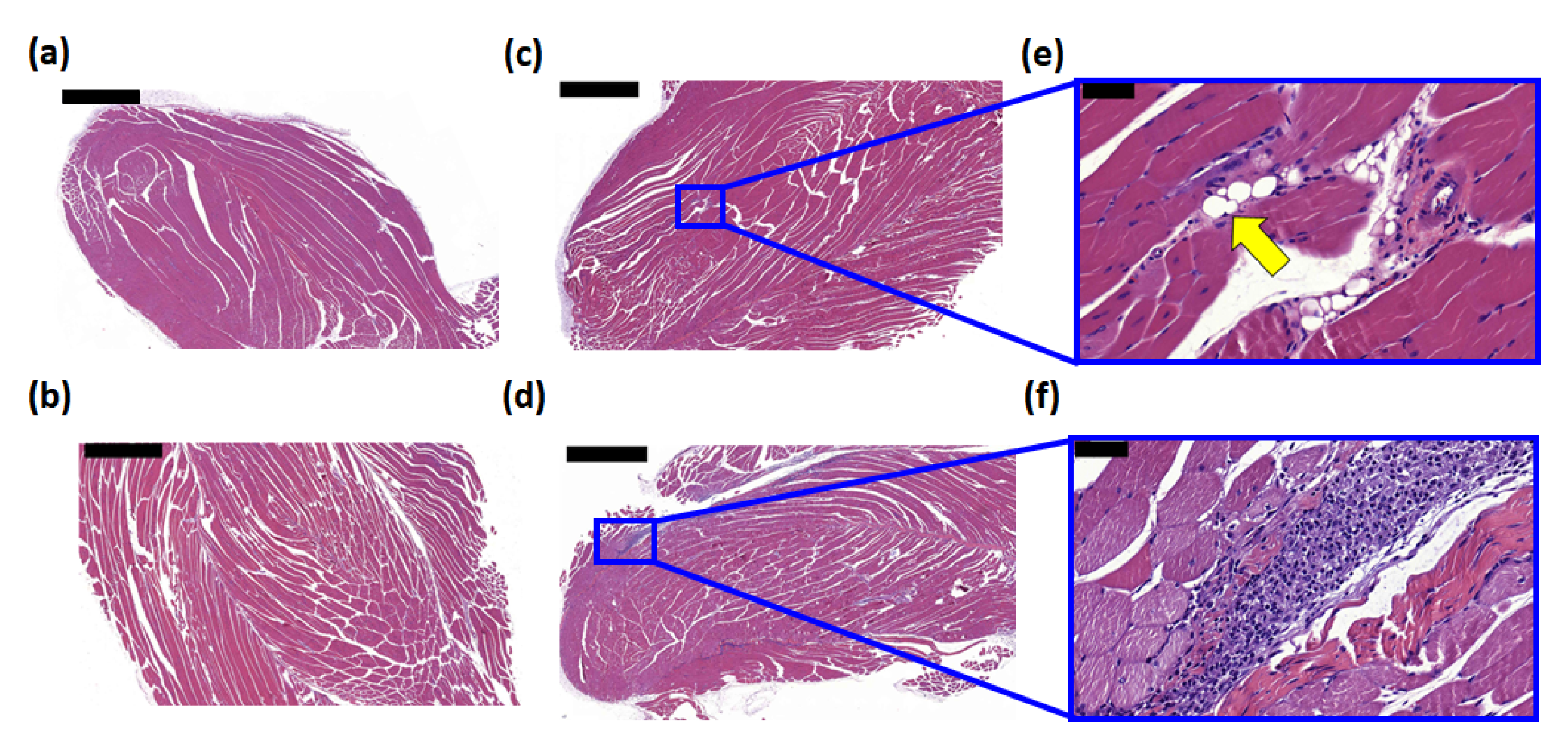
3.4. Histological Examination of the Injection Site
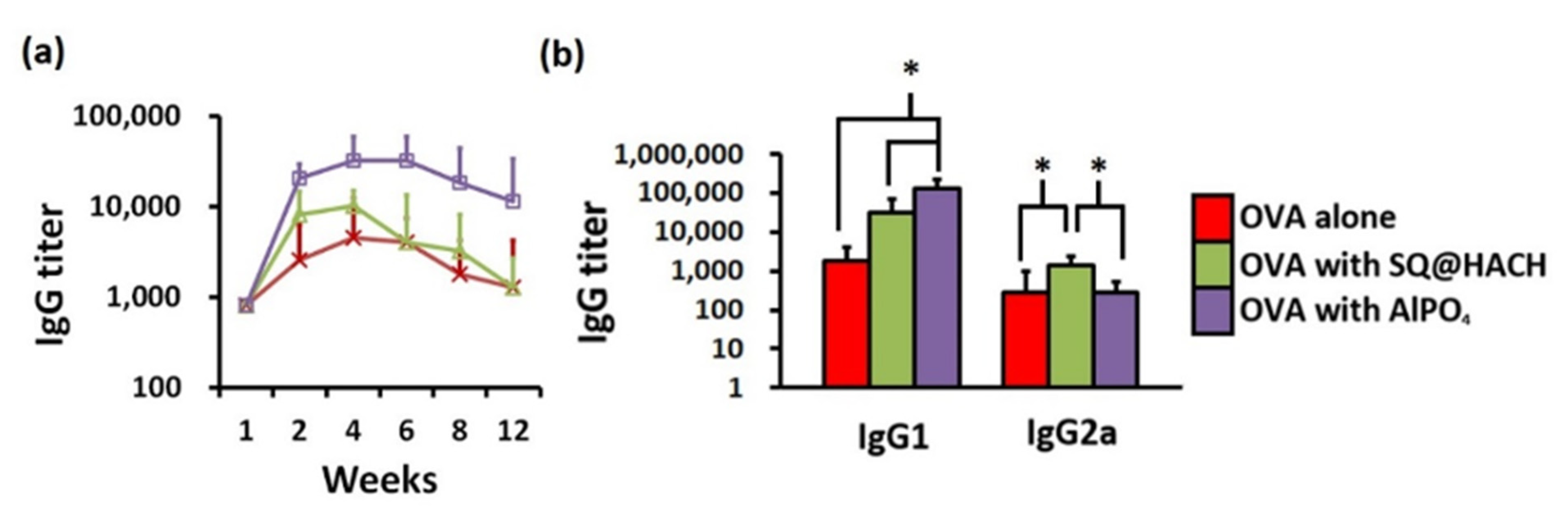
3.5. Serum IgG Antibodies
3.6. Modulation of the Adjuvant-Mediated T Cell Activation
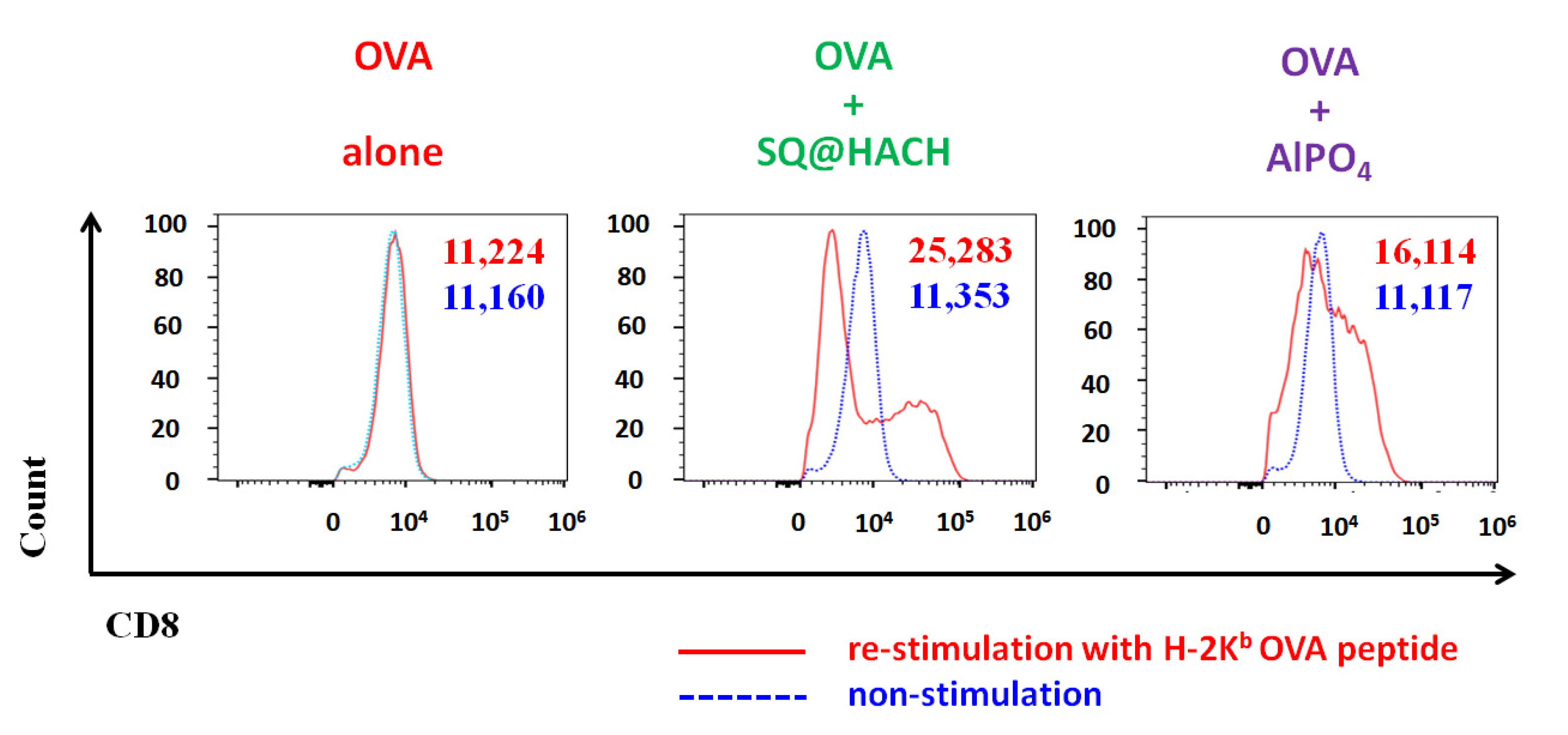
3.7. Evaluation of the OVA-Specific Splenic Cytotoxic T Lymphocyte Response
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mascola, J.R.; Fauci, A.S. Novel vaccine technologies for the 21st century. Nat. Rev. Immunol. 2020, 20, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, M.; Afkhami, S.; Smaill, F.; Miller, M.S.; Lichty, B.D.; Xing, Z. Immunological considerations for COVID-19 vaccine strategies. Nat. Rev. Immunol. 2020, 20, 615–632. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Krishnan, L.; Dicaire, C.J.; Patel, G.B.; Sprott, G.D. Archaeosome vaccine adjuvants induce strong humoral, cell-mediated, and memory responses: Comparison to conventional liposomes and alum. Infect. Immun. 2000, 68, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Tandrup Schmidt, S.; Foged, C.; Smith Korsholm, K.; Rades, T.; Christensen, D. Liposome-based adjuvants for subunit vaccines: Formulation strategies for subunit antigens and immunostimulators. Pharmaceutics 2016, 8, 7. [Google Scholar] [CrossRef]
- Petrovsky, N. Comparative safety of vaccine adjuvants: A summary of current evidence and future needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef]
- Su, J.R.; Moro, P.L.; Ng, C.S.; Lewis, P.W.; Said, M.A.; Cano, M.V. Anaphylaxis after vaccination reported to the Vaccine Adverse Event Reporting System, 1990–2016. J. Allergy Clin. Immunol. 2019, 143, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Sellaturay, P.; Nasser, S.; Ewan, P. Polyethylene glycol–induced systemic allergic reactions (anaphylaxis). J. Allergy Clin. Imunol. Pract. 2021, 9, 670–675. [Google Scholar] [CrossRef]
- Badiu, I.; Geuna, M.; Heffler, E.; Rolla, G. Hypersensitivity reaction to human papillomavirus vaccine due to polysorbate 80. Case Rep. 2012, 2012, bcr0220125797-b. [Google Scholar] [CrossRef] [Green Version]
- Garvey, L.H.; Nasser, S. Anaphylaxis to the first COVID-19 vaccine: Is polyethylene glycol (PEG) the culprit? Br. J. Anaesth. 2021, 126, e106–e108. [Google Scholar] [CrossRef]
- Ho, N.I.; Raaijmakers, T.K.; Adema, G.J. Adjuvants enhancing cross-presentation by dendritic cells: The key to more effective vaccines? Front. Immunol. 2018, 9, 2874. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Control. Release 2021, 333, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Castells, M.C.; Phillips, E.J. Maintaining safety with SARS-CoV-2 vaccines. N. Engl. J. Med. 2021, 384, 643–649. [Google Scholar] [CrossRef]
- Stone, C.A.; Liu, Y.; Relling, M.V.; Krantz, M.S.; Pratt, A.L.; Abreo, A.; Hemler, J.A.; Phillips, E.J. Immediate hypersensitivity to polyethylene glycols and polysorbates: More common than we have recognized. J. Allergy Clin. Imunol. Pract. 2019, 7, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Povsic, T.J.; Lawrence, M.G.; Lincoff, A.M.; Mehran, R.; Rusconi, C.P.; Zelenkofske, S.L.; Huang, Z.; Sailstad, J.; Armstrong, P.W.; Steg, P.G. Pre-existing anti-PEG antibodies are associated with severe immediate allergic reactions to pegnivacogin, a PEGylated aptamer. J. Allergy Clin. Immunol. 2016, 138, 1712–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Choi, H.; Choi, E.S.; Park, M.-H.; Ryu, J.-H. Hyaluronic acid-coated nanomedicine for targeted cancer therapy. Pharmaceutics 2019, 11, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schanté, C.E.; Zuber, G.; Herlin, C.; Vandamme, T.F. Chemical modifications of hyaluronic acid for the synthesis of derivatives for a broad range of biomedical applications. Carbohydr. Polym. 2011, 85, 469–489. [Google Scholar] [CrossRef]
- Bowman, S.; Awad, M.E.; Hamrick, M.W.; Hunter, M.; Fulzele, S. Recent advances in hyaluronic acid based therapy for osteoarthritis. Clin. Transl. Med. 2018, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.S.; Kwon, W.J.; Cho, E.B.; Park, E.J.; Kim, K.H.; Kim, K.J. A case of cellulitis-like foreign body reaction after hyaluronic acid dermal filler injection. Dermatol. Sin. 2018, 36, 46–49. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, J.S.; Yun, J.H.; Kang, H.K.; Baek, J.O.; Roh, J.Y.; Lee, J.R. Foreign body reaction to injectable hyaluronic acid: Late granuloma formation. Ann. Dermatol. 2015, 27, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Beleznay, K.; Carruthers, J.D.; Carruthers, A.; Mummert, M.E.; Humphrey, S. Delayed-onset nodules secondary to a smooth cohesive 20 mg/mL hyaluronic acid filler: Cause and management. Dermatol. Surg. 2015, 41, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.; Ryde, E.; Zetterström, E. Non-immunogenicity of a purified sodium hyaluronate preparation in man. Int. Arch. Allergy Immunol. 1979, 59, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.G.; Strobos, J.; Adkinson, N.F., Jr. Immunogenicity studies of cosmetically administered nonanimal-stabilized hyaluronic acid particles. Dermatol. Surg. 2007, 33, S176–S185. [Google Scholar]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic acid (hyaluronan): A review. Vet. Med. 2008, 53, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angiogenesis in vascular disease: Uncovering RHAMM and CD44 receptor signaling pathways. Matrix Biol. 2007, 26, 58–68. [Google Scholar] [CrossRef]
- Sheehan, K.M.; DeLott, L.B.; West, R.A.; Bonnema, J.D.; DeHeer, D.H. Hyaluronic acid of high molecular weight inhibits proliferation and induces cell death in U937 macrophage cells. Life Sci. 2004, 75, 3087–3102. [Google Scholar] [CrossRef] [PubMed]
- Bodo, M.; Pezzetti, F.; Baroni, T.; Carinci, F.; Arena, N.; Nicoletti, I.; Becchetti, E. Hyaluronic acid modulates growth, morphology and cytoskeleton in embryonic chick skin fibroblasts. Int. J. Dev. Biol. 2002, 37, 349–352. [Google Scholar]
- Harvima, I.T.; Heikura, H.; Hyttinen, M.; Naukkarinen, A. Hyaluronic acid inhibits the adherence and growth of monolayer keratinocytes but does not affect the growth of keratinocyte epithelium. Arch. Dermatol. Res. 2006, 298, 207–219. [Google Scholar] [CrossRef]
- Rayahin, J.E.; Buhrman, J.S.; Zhang, Y.; Koh, T.J.; Gemeinhart, R.A. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater. Sci. Eng. 2015, 1, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Tolg, C.; Hamilton, S.R.; Nakrieko, K.-A.; Kooshesh, F.; Walton, P.; McCarthy, J.B.; Bissell, M.J.; Turley, E.A. Rhamm−/− fibroblasts are defective in CD44-mediated ERK1, 2 motogenic signaling, leading to defective skin wound repair. J. Cell Biol. 2006, 175, 1017–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, Y.; Nagarkatti, P.S.; Nagarkatti, M. Role of CD44 and hyaluronic acid (HA) in activation of alloreactive and antigen-specific T cells by bone marrow-derived dendritic cells. J. Immunother. 2004, 27, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.-h.; Shin, E.-C.; Noh, Y.-W.; Lim, Y.T. Evaluation of hyaluronic acid-based combination adjuvant containing monophosphoryl lipid A and aluminum salt for hepatitis B vaccine. Vaccine 2015, 33, 4762–4769. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cao, F.; Liu, X.; Wang, H.; Zhang, C.; Sun, H.; Wang, C.; Leng, X.; Song, C.; Kong, D.; et al. Hyaluronic acid-modified cationic Lipid–PLGA hybrid nanoparticles as a nanovaccine induce robust humoral and cellular immune responses. ACS Appl. Mater. Interfaces 2016, 8, 11969–11979. [Google Scholar] [CrossRef]
- Briuglia, M.-L.; Rotella, C.; McFarlane, A.; Lamprou, D.A. Influence of cholesterol on liposome stability and on in vitro drug release. Drug Deliv. Transl. Res. 2015, 5, 231–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fieber, W.; Herrmann, A.; Ouali, L.; Velazco, M.I.; Kreutzer, G.; Klok, H.-A.; Ternat, C.; Plummer, C.J.; Månson, J.-A.E.; Sommer, H. NMR diffusion and relaxation studies of the encapsulation of fragrances by amphiphilic multiarm star block copolymers. Macromolecules 2007, 40, 5372–5378. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Huang, M.-H. Emulsifying properties and degradation characteristics of bioresorbable polymeric emulsifiers in aqueous solution and oil-in-water emulsion. Polym. Degrad. Stab. 2017, 139, 138–142. [Google Scholar] [CrossRef]
- Capek, I. Degradation of kinetically-stable o/w emulsions. Adv. Colloid Interfaces Sci. 2004, 107, 125–155. [Google Scholar] [CrossRef]
- Saint-Leger, D.; Bague, A.; Lefebvre, E.; Cohen, E.; Chivot, M. A possible role for squalene in the pathogenesis of acne. II. In vivo study of squalene oxides in skin surface and intra-comedonal lipids of acne patients. Br. J. Dermatol. 1986, 114, 543–552. [Google Scholar] [CrossRef]
- Yanase, N.; Toyota, H.; Hata, K.; Yagyu, S.; Seki, T.; Harada, M.; Kato, Y.; Mizuguchi, J. OVA-bound nanoparticles induce OVA-specific IgG1, IgG2a, and IgG2b responses with low IgE synthesis. Vaccine 2014, 32, 5918–5924. [Google Scholar] [CrossRef]
- Lee-Sayer, S.S.; Dong, Y.; Arif, A.A.; Olsson, M.; Brown, K.L.; Johnson, P. The where, when, how, and why of hyaluronan binding by immune cells. Front. Immunol. 2015, 6, 150. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.; Yan, M.; Liu, Y.; Liu, L.; Ma, G. Photothermally controlled MHC class I restricted CD8+ T-cell responses elicited by hyaluronic acid decorated gold nanoparticles as a vaccine for cancer immunotherapy. Adv. Healthc. Mater. 2018, 7, 1701439. [Google Scholar] [CrossRef]
- Nawwab Al-Deen, F.M.; Selomulya, C.; Kong, Y.Y.; Xiang, S.D.; Ma, C.; Coppel, R.L.; Plebanski, M. Design of magnetic polyplexes taken up efficiently by dendritic cell for enhanced DNA vaccine delivery. Gene Ther. 2014, 21, 212–218. [Google Scholar] [CrossRef]
- Johnson, L.A.; Banerji, S.; Lawrance, W.; Gileadi, U.; Prota, G.; Holder, K.A.; Roshorm, Y.M.; Hanke, T.; Cerundolo, V.; Gale, N.W.; et al. Dendritic cells enter lymph vessels by hyaluronan-mediated docking to the endothelial receptor LYVE-1. Nat. Immunol. 2017, 18, 762–770. [Google Scholar] [CrossRef]
- Yoo, E.; Salyer, A.C.; Brush, M.J.; Li, Y.; Trautman, K.L.; Shukla, N.M.; De Beuckelaer, A.; Lienenklaus, S.; Deswarte, K.; Lambrecht, B.N. Hyaluronic acid conjugates of TLR7/8 agonists for targeted delivery to secondary lymphoid tissue. Bioconj. Chem. 2018, 29, 2741–2754. [Google Scholar] [CrossRef]
- Ruppert, S.; Hawn, T.; Arrigoni, A.; Wight, T.; Bollyky, P. Tissue integrity signals communicated by high-molecular weight hyaluronan and the resolution of inflammation. Immunol. Res. 2014, 58, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Scheibner, K.A.; Lutz, M.A.; Boodoo, S.; Fenton, M.J.; Powell, J.D.; Horton, M.R. Hyaluronan fragments act as an endogenous danger signal by engaging TLR2. J. Immunol. 2006, 177, 1272–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Termeer, C.; Benedix, F.; Sleeman, J.; Fieber, C.; Voith, U.; Ahrens, T.; Miyake, K.; Freudenberg, M.; Galanos, C.; Simon, J.C. Oligosaccharides of Hyaluronan activate dendritic cells via toll-like receptor 4. J. Exp. Med. 2002, 195, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Ang, B.; Xu, X.; Huang, X.; Wu, Y.; Sun, Y.; Wang, W.; Li, N.; Cao, X.; Wan, T. TLR4 is essential for dendritic cell activation and anti-tumor T-cell response enhancement by DAMPs released from chemically stressed cancer cells. Cell. Mol. Immunol. 2014, 11, 150–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandran, S.S.; Verhoeven, D.; Teijaro, J.R.; Fenton, M.J.; Farber, D.L. TLR2 engagement on dendritic cells promotes high frequency effector and memory CD4 T cell responses. J. Immunol. 2009, 183, 7832–7841. [Google Scholar] [CrossRef]
- Dalla Pietà, A.; Carpanese, D.; Grigoletto, A.; Tosi, A.; Dalla Santa, S.; Pedersen, G.K.; Christensen, D.; Meléndez-Alafort, L.; Barbieri, V.; De Benedictis, P. Hyaluronan is a natural and effective immunological adjuvant for protein-based vaccines. Cell. Mol. Immunol. 2021, 18, 1197–1210. [Google Scholar] [CrossRef]
- Falo, L.D.; Kovacsovics-Bankowski, M.; Thompson, K.; Rock, K.L. Targeting antigen into the phagocytic pathway in vivo induces protective tumour immunity. Nat. Med. 1995, 1, 649–653. [Google Scholar] [CrossRef]
- Ganji, A.; Farahani, I.; Khansarinejad, B.; Ghazavi, A.; Mosayebi, G. Increased expression of CD8 marker on T-cells in COVID-19 patients. Blood Cells Mol. Dis. 2020, 83, 102437. [Google Scholar] [CrossRef]
- Menares, E.; Gálvez-Cancino, F.; Cáceres-Morgado, P.; Ghorani, E.; López, E.; Díaz, X.; Saavedra-Almarza, J.; Figueroa, D.A.; Roa, E.; Quezada, S.A.; et al. Tissue-resident memory CD8+ T cells amplify anti-tumor immunity by triggering antigen spreading through dendritic cells. Nat. Commun. 2019, 10, 4401. [Google Scholar] [CrossRef] [Green Version]
- Woodland, D.L.; Kohlmeier, J.E. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat. Rev. Immunol. 2009, 9, 153–161. [Google Scholar] [CrossRef]
- Mercier, B.C.; Cottalorda, A.; Coupet, C.-A.; Marvel, J.; Bonnefoy-Bérard, N. TLR2 engagement on CD8 T cells enables generation of functional memory cells in response to a suboptimal TCR signal. J. Immunol. 2009, 182, 1860–1867. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Arévalo, D.; Zeng, M. Nanoparticle-based vaccines: Opportunities and limitations. In Nanopharmaceuticals; Elsevier: Amsterdam, The Netherlands, 2020; pp. 135–150. [Google Scholar]
- Liu, L.; Kshirsagar, P.; Christiansen, J.; Gautam, S.K.; Aithal, A.; Gulati, M.; Kumar, S.; Solheim, J.C.; Batra, S.K.; Jain, M. Polyanhydride nanoparticles stabilize pancreatic cancer antigen MUC4β. J. Biomed. Mater. Res. Part A 2021, 109, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Gautam, S.K.; Kshirsagar, P.; Ross, K.A.; Spagnol, G.; Sorgen, P.; Wannemuehler, M.J.; Narasimhan, B.; Solheim, J.C.; Kumar, S. Amphiphilic polyanhydride-based recombinant MUC4β-nanovaccine activates dendritic cells. Genes Cancer 2019, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhang, S.H.; Won, N.; Lee, T.-J.; Jin, H.; Nam, J.; Park, J.; Chung, H.; Park, H.-S.; Sung, Y.-E.; Hahn, S.K. Hyaluronic acid-quantum dot conjugates for in vivo lymphatic vessel imaging. ACS Nano 2009, 3, 1389–1398. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C Element (Mole%) a | N Element (Mole%) a | C/N b | DS(%) c | |
|---|---|---|---|---|
| HA | 3.125 | 0.223 | 14.0 | - |
| HACH10 | 3.314 | 0.226 | 14.7 | 9.2 |
| HACH20 | 3.143 | 0.210 | 15.0 | 13.3 |
| HACH30 | 3.361 | 0.216 | 15.6 | 21.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-A.; Ho, H.-M.; Venkatesan, P.; Huang, C.-Y.; Cheng, Y.-J.; Lin, Y.-H.; Lin, H.-Y.; Chen, T.-Y.; Huang, M.-H.; Lai, P.-S. Hyaluronic Acid-Glycine-Cholesterol Conjugate-Based Nanoemulsion as a Potent Vaccine Adjuvant for T Cell-Mediated Immunity. Pharmaceutics 2021, 13, 1569. https://doi.org/10.3390/pharmaceutics13101569
Lin C-A, Ho H-M, Venkatesan P, Huang C-Y, Cheng Y-J, Lin Y-H, Lin H-Y, Chen T-Y, Huang M-H, Lai P-S. Hyaluronic Acid-Glycine-Cholesterol Conjugate-Based Nanoemulsion as a Potent Vaccine Adjuvant for T Cell-Mediated Immunity. Pharmaceutics. 2021; 13(10):1569. https://doi.org/10.3390/pharmaceutics13101569
Chicago/Turabian StyleLin, Chih-An, Hui-Min Ho, Parthiban Venkatesan, Chiung-Yi Huang, Yu-Jhen Cheng, Yu-Hsing Lin, Hua-Yang Lin, Tzu-Yang Chen, Ming-Hsi Huang, and Ping-Shan Lai. 2021. "Hyaluronic Acid-Glycine-Cholesterol Conjugate-Based Nanoemulsion as a Potent Vaccine Adjuvant for T Cell-Mediated Immunity" Pharmaceutics 13, no. 10: 1569. https://doi.org/10.3390/pharmaceutics13101569