Surface Active Agents and Their Health-Promoting Properties: Molecules of Multifunctional Significance

 , , , and
, , , and
Abstract
:1. Introduction
2. Structure of Synthetic and BioSurfactants
3. Anti-Microbial Properties
4. Anti-Oxidant Properties
5. Anti-Viral Properties
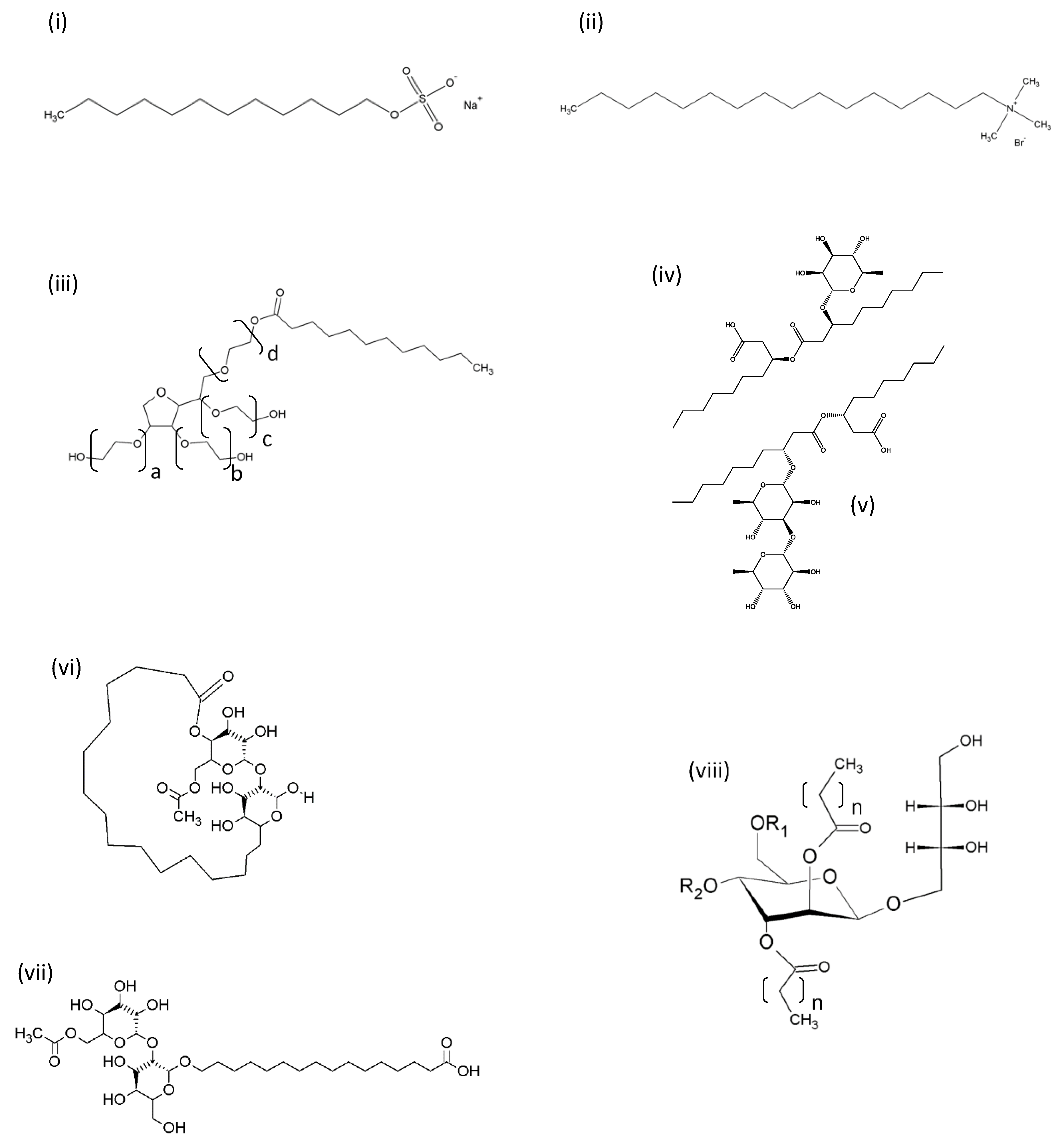
6. Anti-Inflammatory Properties
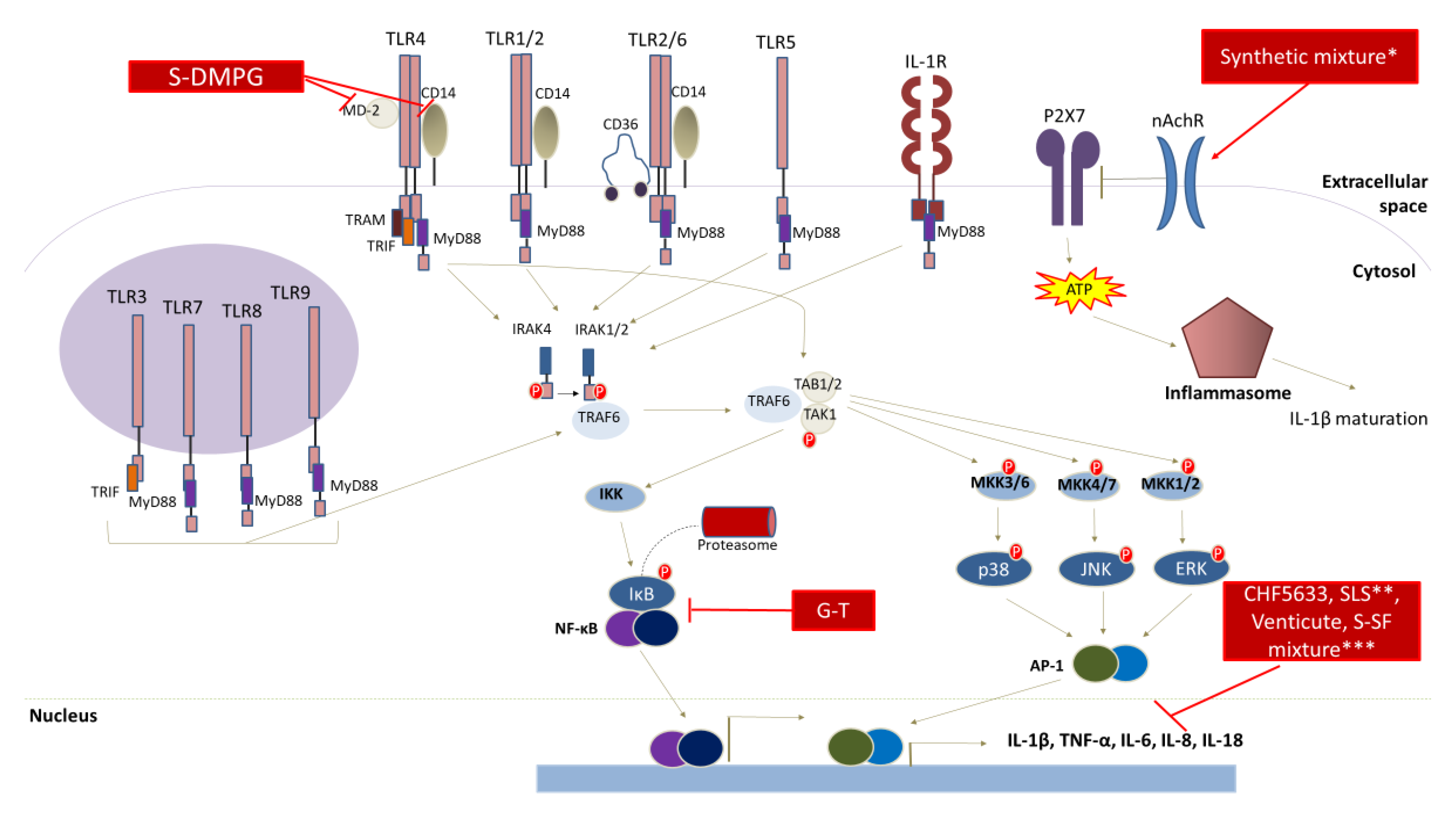
7. Anti-Cancer Properties
8. Anti-Aging Properties
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schramm, L.L.; Stasiuk, E.N.; Marangoni, D.G. Surfactants and their applications. Annu. Rep. Prog. Chem. Sect. C Phys. 2003, 99, 3–48. [Google Scholar] [CrossRef]
- Pedersen, J.N.; Lyngsø, J.; Zinn, T.; Otzen, D.E.; Pedersen, J.S. A complete picture of protein unfolding and refolding in surfactants. Chem. Sci. 2020, 11, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Otzen, D.E. Biosurfactants and surfactants interacting with membranes and proteins: Same but different? Biochim. Biophys. Acta-Biomembr. 2017, 1859, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Sałek, K.; Euston, S.R. Sustainable microbial biosurfactants and bioemulsifiers for commercial exploitation. Process Biochem. 2019, 85, 143–155. [Google Scholar] [CrossRef]
- Hasenhuettl, G.L. Synthesis and commercial preparation of food emulsifiers. In Food Emulsifiers and Their Applications, 3rd ed.; Hasenhuettl, G.L., Hartel, R., Eds.; Springer: New York, NY, USA, 2019; pp. 11–39. [Google Scholar]
- Bell, B.G.; Schellevis, F.; Stobberingh, E.; Goossens, H.; Pringle, M. A systematic review and meta-analysis of the effects of antibiotic consumption on antibiotic resistance. BMC Infect. Dis. 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Wang, F.; Chen, H.; Li, M.; Qiao, F.; Liu, Z.; Hou, Y.; Wu, C.; Fan, Y.; Liu, L.; et al. Selective Antimicrobial Activities and Action Mechanism of Micelles Self-Assembled by Cationic Oligomeric Surfactants. ACS Appl. Mater Interfaces 2016, 8, 4242–4249. [Google Scholar] [CrossRef]
- Shaban, S.M.; Abd-Elaal, A.A. Studying the silver nanoparticles influence on thermodynamic behavior and antimicrobial activities of novel amide Gemini cationic surfactants. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 76, 871–885. [Google Scholar] [CrossRef]
- Kashapov, R.R.; Razuvayeva, Y.S.; Ziganshina, A.Y.; Mukhitova, R.K.; Sapunova, A.S.; Voloshina, A.D.; Zakharova, L.Y. Self-assembling and biological properties of single-chain dicationic pyridinium-based surfactants. Colloids Surf. B Biointerfaces 2019, 175, 351–357. [Google Scholar] [CrossRef]
- Colomer, A.; Pinazo, A.; Manresa, M.A.; Vinardell, M.P.; Mitjans, M.; Infante, M.R.; Pérez, L. Cationic surfactants derived from lysine: Effects of their structure and charge type on antimicrobial and hemolytic activities. J. Med. Chem. 2011, 54, 989–1002. [Google Scholar] [CrossRef]
- Perinelli, D.R.; Petrelli, D.; Vitali, L.A.; Bonacucina, G.; Cespi, M.; Vllasaliu, D.; Giorgioni, G.; Palmieri, G.F. Quaternary Ammonium Leucine-Based Surfactants: The Effect of a Benzyl Group on Physicochemical Properties and Antimicrobial Activity. Pharmaceutics 2019, 11, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.F.; Zepeda-Velazquez, L.; Brook, M.A. Tunable, antibacterial activity of silicone polyether surfactants. Colloids Surf. B Biointerfaces 2015, 132, 216–224. [Google Scholar] [CrossRef]
- Pinazo, A.; Petrizelli, V.; Bustelo, M.; Pons, R.; Vinardell, M.P.; Mitjans, M.; Manresa, A.; Perez, L. New cationic vescicles prepared with double chain surfactants from arginine: Role of the hydrophobic group on the antimicrobial activity and cytotoxicity. Colloids Surf. B Biointerfaces 2016, 141, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, R.; Shibata, M.; Koeda, S.; Miyagawa, A.; Yamamura, H.; Mizuno, T. Development of New Antimicrobial Agents from Cationic PG-surfactants containing oligo-Lys peptides. Bioconjug. Chem. 2018, 29, 4072–4082. [Google Scholar] [CrossRef]
- Díaz De Rienzo, M.A.; Stevenson, P.; Marchant, R.; Banat, I.M. Antibacterial properties of biosurfactants against selected Gram-positive and-negative bacteria. FEMS Microbiol. Lett. 2016, 363, fnv224. [Google Scholar] [CrossRef] [Green Version]
- Yalçin, E.; Ergene, A. Screening the antimicrobial activity of biosurfactants produced by microorganisms isolated from refinery wastewaters. J. Appl. Biol. Sci. 2009, 3, 163–168. [Google Scholar]
- De Freitas Ferreira, J.; Vieira, E.A.; Nitschke, M. The antibacterial activity of rhamnolipid biosurfactant is pH dependent. Food Res. Int. 2019, 116, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.; Nitschke, M. Antimicrobial activity of rhamnolipids against Listeria monocytogenes and their synergistic interaction with nisin. Food Control 2013, 29, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Elshikh, M.; Moya-Ramírez, I.; Moens, H.; Roelants, S.L.K.W.; Soetaert, W.; Marchant, R.; Banat, I.M. Rhamnolipids and lactonic sophorolipids: Natural antimicrobial surfactants for oral hygiene. J. Appl. Microbiol. 2017, 123, 1111–1123. [Google Scholar] [CrossRef]
- Silveira, V.A.I.; Freitas, C.A.U.Q.; Celligoi, M.A.P.C. Antimicrobial applications of sophorolipid from Candida bombicola: A promising alternative to conventional drugs. J. Appl. Biol. Biotechnol. 2018, 6, 87–90. [Google Scholar]
- De Rienzo, M.A.D.; Banat, I.M.; Dolman, B.; Winterburn, J.; Martin, P.J. Sophorolipid biosurfactants: Possible uses as antibacterial and antibiofilm agent. New Biotechnol. 2015, 32, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Ceresa, C.; Fracchia, L.; Williams, M.; Banat, I.M.; De Rienzo, M.D. The effect of sophorolipids against microbial biofilms on medical-grade silicone. J. Biotechnol. 2020, 309, 34–43. [Google Scholar] [CrossRef]
- Valotteau, C.; Banat, I.M.; Mitchell, C.A.; Lydon, H.; Marchant, R.; Babonneau, F.; Pradier, C.M.; Baccile, N.; Humblot, V. Antibacterial properties of sophorolipid modified gold surfaces against Gram positive and Gram negative pathogens. Colloid Surf. B 2017, 157, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valotteau, C.; Baccile, N.; Humblot, V.; Roelants, S.; Soetaert, W.; Stevens, C.V.; Dufrêne, Y.F. Nanoscale antiadhesion properties of sophorolipid-coated surfaces against pathogenic bacteria. Nanoscale Horizons 2019, 4, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Shu, Q.; Niu, Y.; Zhao, W.; Chen, Q. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Ceresa, C.; Hutton, S.; Lajarin-Cuesta, M.; Heaton, R.; Hargreaves, I.; Fracchia, L.; De Rienzo, M.A.D. Production of Mannosylerythritol Lipids (MELs) to be Used as Antimicrobial Agents Against S. aureus ATCC 6538. Curr. Microbiol. 2020, 77, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Janek, T.; Krasowska, A.; Czyżnikowska, Ż.; Łukaszewicz, M. Trehalose Lipid Biosurfactant Reduces Adhesion of Microbial Pathogens to Polystyrene and Silicone Surfaces: An Experimental and Computational Approach. Front. Microbiol. 2018, 9, 2441. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; García-Fernández, C.; Alonso-Calleja, C.; Capita, R. Susceptibility of Listeria monocytogenesplanktonic cultures and biofilms to sodiumhypochlorite and benzalkoniumchloride. Food Microbiol. 2019, 82, 533–540. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, Y.S. Effect of Sub Minimal Inhibitory Concentration Cetylpyridinium Chloride on Biofilm Formation and Hydropyobicity of Streptococci and Actinomycetes. J. Adv. Microbiol. 2018, 9, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, K.; Amaechi, B.T.; Ralph, R.H.; Lee, V.A. Antimicrobial activity of nanoemulsion on cariogenic planktonic and biofilm organisms. Arch. Oral Biol. 2012, 57, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, C.; Hattori, Y.; Yagi, S.; Murai, T.; Takai, C.; Ogawa, N.; Tanemura, M.; Fuji, M.; Kawashima, Y.; Yamamoto, H. Optimization of ionic liquid-incorporated PLGA nanoparticles for treatment of biofilm infections. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 97, 78–83. [Google Scholar] [CrossRef]
- Ruiz, A.; Pinazo, A.; Pérez, L.; Manresa, A.; Marqués, A.M. Green Catanionic Gemini Surfactant-Lichenysin Mixture: Improved Surface, Antimicrobial, and Physiological Properties. ACS Appl. Mater Interfaces 2017, 9, 22121–22131. [Google Scholar] [CrossRef] [PubMed]
- Obłąk, E.; Piecuch, A.; Dworniczek, E.; Olejniczak, T. The influence of biodegradable gemini surfactants, N,N’-bis(1-decyloxy-1-oxopronan-2-yl)-N,N,N’,N’ tetramethylpropane-1,3-diammonium dibromide and N,N’-bis(1-dodecyloxy-1-oxopronan-2-yl) N,N,N’,N’-tetramethylethane-1,2-diammonium dibromide, on fungal biofilm and adhesion. J. Oleo Sci. 2015, 64, 527–537. [Google Scholar] [PubMed] [Green Version]
- E Silva, S.S.; Carvalho, J.W.P.; Aires, C.P.; Nitschke, M. Disruption of Staphylococcus aureus biofilms using rhamnolipid biosurfactants. J. Dairy Sci. 2017, 100, 7864–7873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksic, I.; Petkovic, M.; Jovanovic, M.; Milivojevic, D.; Vasiljevic, B.; Nikodinovic-Runic, J.; Senerovic, L. Anti-biofilm properties of bacterial di-rhamnolipids and their semi-synthetic amide derivatives. Front. Microbiol. 2017, 8, 2454. [Google Scholar] [CrossRef]
- De Rienzo, M.A.D.; Martin, P.J. Effect of mono and di-rhamnolipids on biofilms pre-formed by Bacillus subtilis BBK006. Curr. Microbiol. 2016, 73, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do Valle Gomes, M.Z.; Nitschke, M. Evaluation of rhamnolipid and surfactin to reduce the adhesion and remove biofilms of individual and mixed cultures of food pathogenic bacteria. Food Control 2012, 25, 441–447. [Google Scholar] [CrossRef]
- Singh, N.; Pemmaraju, S.C.; Pruthi, P.A.; Cameotra, S.S.; Pruthi, V. Candida biofilm disrupting ability of di-rhamnolipid (RL-2) produced from Pseudomonas aeruginosa DSVP20. Appl. Biochem. Biotechnol. 2013, 169, 2374–2391. [Google Scholar] [CrossRef]
- Dusane, D.H.; Dam, S.; Nancharaiah, Y.V.; Kumar, A.R.; Venugopalan, V.P.; Zinjarde, S.S. Disruption of Yarrowia lipolytica biofilms by rhamnolipid biosurfactant. Aquat.Biosyst. 2012, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kurowska, A.; Ghate, V.; Kodoth, A.; Shah, A.; Shah, A.; Vishalakshi, B.; Prakash, B.; Lewis, S.A. Non-Propellant Foams of Green Nano-Silver and Sulfadiazine: Development and In Vivo Evaluation for Burn Wounds. Pharm. Res. 2019, 36, 122. [Google Scholar] [CrossRef]
- Percival, S.L.; Mayer, D.; Salisbury, A.-M. Efficacy of a surfactant-based wound dressing on biofilm control. Wound Repair Regen. 2017, 25, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, A.M.; Percival, S.L. Efficacy of a Surfactant-Based Wound Dressing in the Prevention of Biofilms. Adv. Skin Wound Care 2018, 31, 514–520. [Google Scholar] [CrossRef]
- Palumbo, F.P.; Harding, K.G.; Abbritti, F.; Bradbury, S.; Cech, J.D.; Ivins, N.; Klein, D.; Menzinger, G.; Meuleneire, F.; Seratoni, S.; et al. New Surfactant-based Dressing Product to Improve Wound Closure Rates of Nonhealing Wounds: A European Multicenter Study Including 1036 Patients. Wounds 2016, 28, 233–240. [Google Scholar]
- El-Nahhal, I.M.; Elmanama, A.A.; El Ashgar, N.M.; Amara, N.; Selmane, M.; Chehimi, M.M. Stabilization of nano-structured ZnO particles onto the surface of cotton fibers using different surfactants and their antimicrobial activity. Ultrason. Sonochem. 2017, 38, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Benavent, C.; García-Herrero, V.; Torrado, C.; Torrado-Santiago, S. Nystatin antifungal micellar systems on endotracheal tubes: Development, characterization and in vitro evaluation. Pharmazie 2019, 74, 34–38. [Google Scholar] [PubMed]
- Majchrzycka, K.; Okrasa, M.; Szulc, J.; Brycki, B.; Gutarowska, B. Time-Dependent Antimicrobial Activity of Filtering Nonwovens with Gemini Surfactant-Based Biocides. Molecules 2017, 22, 1620. [Google Scholar] [CrossRef] [PubMed]
- Diaz, D.; Church, J.; Young, M.; Kim, K.T.; Park, J.; Hwang, Y.B.; Santra, S.; Lee, W.H. Silica-quaternary ammonium “Fixed-Quat” nanofilm coated fiberglass mesh for water disinfection and harmful algal blooms control. J. Environ. Sci. (China) 2019, 82, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Chimote, G.; Banerjee, R. Evaluation of antitubercular drug-loaded surfactants as inhalable drug-delivery systems for pulmonary tuberculosis. J. Biomed. Mater. Res. A 2009, 89, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Sung, C.T.; Aljuffali, I.A.; Chen, C.H.; Hu, K.Y.; Fang, J.Y. Intravenous anti-MRSA phosphatiosomes mediate enhanced affinity to pulmonary surfactants for effective treatment of infectious pneumonia. Nanomedicine 2018, 14, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, S.; Gu, Y.; Tang, X.; Wang, T.; Wu, J.; Liu, J. Development of sulconazole-loaded nanoemulsions for enhancement of transdermal permeation and antifungal activity. Int. J. Nanomed. 2019, 28, 3955–3966. [Google Scholar] [CrossRef] [Green Version]
- Shah, J.; Nair, A.B.; Jacob, S.; Patel, R.K.; Shah, H.; Shehata, T.M.; Morsy, M.A. Nanoemulsion Based Vehicle for Effective Ocular Delivery of Moxifloxacin Using Experimental Design and Pharmacokinetic Study in Rabbits. Pharmaceutics 2019, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Eid, H.M.; Elkomy, M.H.; El Menshawe, S.F.; Salem, H.F. Development, Optimization, and In Vitro/In Vivo Characterization of Enhanced Lipid Nanoparticles for Ocular Delivery of Ofloxacin: The Influence of Pegylation and Chitosan Coating. AAPS PharmSciTech 2019, 20, 183. [Google Scholar] [CrossRef] [PubMed]
- Ziech, D.; Franco, R.; Georgakilas, A.G.; Georgakila, S.; Malamou-Mitsi, V.; Schoneveld, O.; Pappa, A.; Panayiotidis, M.I. The role of reactive oxygen species and oxidative stress in environmental carcinogenesis and biomarker development. Chem. Biol. Interact. 2010, 188, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Hanafi, R.; Anestopoulos, I.; Voulgaridou, G.P.; Franco, R.; Georgakilas, A.G.; Ziech, D.; Malamou-Mitsi, V.; Pappa, A.; Panayiotidis, M.I. Oxidative stress based-biomarkers in oral carcinogenesis: How far have we gone? Curr. Mol. Med. 2012, 12, 698–703. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Yepes, J.; Burns, M.; Anandhan, A.; Khalimonchuk, O.; del Razo, L.M.; Quintanilla-Vega, B.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Oxidative stress, redox signaling, and autophagy: Cell death versus survival. Antioxid. Redox Signal. 2014, 21, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive oxygen species (ROS)-induced genetic and epigenetic alterations in human carcinogenesis. Mutat. Res. 2011, 711, 167–173. [Google Scholar] [CrossRef]
- Ziech, D.; Anestopoulos, I.; Hanafi, R.; Voulgaridou, G.-P.; Franco, R.; Georgakilas, A.G.; Pappa, A.; Panayiotidis, M.I. Pleiotrophic effects of natural products in ROS-induced carcinogenesis: The role of plant-derived natural products in oral cancer chemoprevention. Cancer Lett. 2012, 327, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals and antioxidants-quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Voulgaridou, G.-P.; Anestopoulos, I.; Franco, R.; Panayiotidis, M.I.; Pappa, A. DNA damage induced by endogenous aldehydes: Current state of knowledge. Mutat. Res. 2011, 711, 13–27. [Google Scholar] [CrossRef]
- Garcia-Garcia, A.; Rodriguez-Rocha, H.; Madayiputhiya, N.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Biomarkers of protein oxidation in human disease. Curr. Mol. Med. 2012, 12, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Alamed, J.; Chaiyasit, W.; McClements, D.J.; Decker, E.A. Relationships between free radical scavenging and antioxidant activity in foods. J. Agric. Food Chem. 2009, 57, 2969–2976. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2006, 23, 26–78. [Google Scholar] [CrossRef] [PubMed]
- Balboa, E.M.; Conde, E.; Moure, A.; Falque, E.; Dominguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Ngo, D.H.; Kim, S.K. Antioxidant effects of chitin, chitosan, and their derivatives. Adv. Food Nutr. Res. 2014, 73, 15–31. [Google Scholar]
- Luther, M.; Parry, J.; Moore, J.; Meng, J.; Zhang, Y.; Cheng, Z.; Yu, L.L. Inhibitory effect of Chardonnay and black raspberry seed extracts on lipid oxidation in fish oil and their radical scavenging and antimicrobial properties. Food Chem. 2007, 104, 1065–1073. [Google Scholar] [CrossRef]
- McClements, D.J.; Decker, E.A. Lipid oxidation in oil-in-water emulsions: Impact of molecular environment on chemical reactions in heterogeneous food systems. J. Food Sci. 2006, 65, 1270–1282. [Google Scholar] [CrossRef]
- McClements, D.J.; Rao, J. Food-Grade Nanoemulsions: Formulation, Fabrication, Properties, Performance, Biological Fate, and Potential Toxicity. Crit. Rev. Food Sci. Nutr. 2011, 51, 285–330. [Google Scholar] [CrossRef]
- Yi, B.; Kim, M.J.; Lee, J. Effects of emulsifier charges on the oxidative stability in oil-in-water emulsions under riboflavin photosensitization. Food Sci. Biotechnol. 2016, 25, 1003–1009. [Google Scholar] [CrossRef]
- Schwarz, K.; Huang, S.W.; German, J.B.; Tiersch, B.; Hartmann, J.; Frankel, E.N. Activities of antioxidants are affected by colloidal properties of oil-in-water and water-in-oil emulsions and bulk oils. J. Agric. Food Chem. 2000, 48, 4874–4882. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasit, W.; Elias, R.J.; McClements, D.J.; Decker, E.A. Role of physical structures in bulk oils on lipid oxidation. Crit. Rev. Food Sci. Nutr. 2007, 47, 299–317. [Google Scholar] [CrossRef]
- Lee, J.H.; Decker, E.A. Effects of metal chelator, sodium azide, and superoxide dismutase (SOD) on the oxidative stability in riboflavin photosensitized O/W emulsion systems. J. Agric. FoodChem. 2011, 59, 6271–6276. [Google Scholar] [CrossRef]
- McClements, D.J.; Decker, E. Interfacial Antioxidants: A Review of Natural and Synthetic Emulsifiers and Coemulsifiers That Can Inhibit Lipid Oxidation. J. Agric. Food Chem. 2018, 66, 20–35. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.E.; Wang, W.D.; Chen, H.W.; Li, C. Autoxidation of unsaturated lipids in food emulsion. Crit. Rev. Food Sci. 2011, 51, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, M.P.; Chaiyasit, W.; Brannan, R.G.; McClements, D.J.; Decker, E.A. Ability of surfactant headgroup size to alter lipid and antioxidant oxidation in oil-in-water emulsions. J. Agric. Food Chem. 2000, 48, 2057–2061. [Google Scholar] [CrossRef]
- Chaiyasit, W.; Silvestre, M.P.; McClements, D.J.; Decker, E.A. Ability of surfactant hydrophobic tail group size to alter lipid oxidation in oil-in-water emulsions. J. Agric. Food Chem. 2000, 48, 3077–3080. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Decker, E.A.; Lee, J.H. Effects of chlorophyll photosensitization on the oxidative stability in O/W emulsion. Food Chem. 2012, 33, 1449–1455. [Google Scholar] [CrossRef]
- Lee, J.H.; Min, D.B. Changes of headspace volatiles in milk with riboflavin photosensitization. J. Food Sci. 2009, 74, 563–568. [Google Scholar] [CrossRef]
- Lee, J.H.; Min, D.B. Analysis of volatile compounds from chlorophyll photosensitized linoleic acid by headspace Solid-phase microextraction (HS-SPME). Food Sci. Biotechnol. 2010, 19, 611–616. [Google Scholar] [CrossRef]
- Mancuso, J.R.; McClements, D.J.; Decker, E.A. The effects of surfactant type, pH, and chelators on the oxidation of salmon oil-in-water emulsions. J. Agric. Food Chem. 1999, 47, 4112–4116. [Google Scholar] [CrossRef] [PubMed]
- Uluata, S.; McClements, D.J.; Decker, E.A. Physical Stability, Autoxidation, and Photosensitized Oxidation of ω-3 Oils in Nanoemulsions Prepared with Natural and Synthetic Surfactants. J. Agric. Food Chem. 2015, 63, 9333–9340. [Google Scholar] [CrossRef] [PubMed]
- Berton, C.; Genot, C.; Guilbert, D.; Ropers, M.H. Effect of lateral heterogeneity in mixed surfactant-stabilized interfaces on the oxidation of unsaturated lipids in oil-in-water emulsions. J. Colloid Interface Sci. 2012, 377, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Weiszhar, Z.; Czucz, J.; Revesz, C.; Rosivall, L.; Szebeni, J.; Rozsnyay, Z. Complement activation bypolyethoxylatedpharmaceuticalsurfactants:Cremophor-EL, Tween-80 and Tween-20. Eur. J. Pharm. Sci. 2012, 45, 492–498. [Google Scholar] [CrossRef]
- Perez-Roses, R.; Risco, E.; Vila, R.; Penalver, P.; Canigueral, S. Antioxidant activity of Tween-20 and Tween-80 evaluated through different in-vitro tests. J. Pharm. Pharmacol. 2015, 67, 666–672. [Google Scholar] [CrossRef]
- Sadoqi, M.; Lau-Cam, C.A.; Wu, S.H. Investigation of the micellar properties of the tocopheryl polyethylene glycolsuccinate surfactants TPGS 400 and TPGS 1000 by steady state fluorometry. J. Colloid Interface Sci. 2009, 333, 585–589. [Google Scholar] [CrossRef]
- Shah, A.R.; Banerjee, R. Effect of D-a-tocopheryl polyethylene glycol 1000 succinate (TPGS) on surfactant monolayers. Colloid Surf. B Biointerfaces 2011, 85, 116–124. [Google Scholar] [CrossRef]
- Guo, Y.; Luo, J.; Tan, S.; Otieno, B.O.; Zhang, Z. The applications of Vitamin E TPGS in drug delivery. Eur. J. Pharm. Sci. 2013, 49, 175–186. [Google Scholar] [CrossRef]
- Constantinides, P.P.; Han, J.H.; Davis, S.S. Advances in the use of tocols as drug delivery vehicles. Pharm. Res. 2006, 23, 243–255. [Google Scholar] [CrossRef]
- Varma, M.V.; Panchagnula, R. Enhanced oral paclitaxel absorption with Vitamin E-TPGS: Effect on solubility and permeability in vitro, in situ and in vivo. Eur. J. Pharm. Sci. 2005, 25, 445–453. [Google Scholar] [CrossRef]
- Dintaman, J.M.; Silverman, J.A. Inhibition of P-glycoprotein by D-a-tor ocopheryl polyethylene glycol 1000 succinate (TPGS). Pharm. Res. 1999, 16, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Liu, Y.T.; Feng, S.S. Formulation of Docetaxel byfolic acid-conjugated D-a-tocopheryl polyethylene glycol succinate 2000 (VitaminETPGS(2k)) micelles for targeted and synergistic chemotherapy. Biomaterials 2011, 32, 4058–4066. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.; Trendelenburg, C.F.; Diez-Fernandez, C.; Prieto, P.; Houy, S.; Trommer, W.E.; Cordier, A. Cyclosporine A-induced oxidative stress in rat hepatocytes. J. Pharmacol. Exp. Ther. 1997, 280, 1328–1334. [Google Scholar]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Technol. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Marzocchi, S.; Anankanbil, S.; Caboni, M.F.; Guo, Z. Enzymatic alkylsuccinylation of tyrosol: Synthesis, characterization and property evaluation as a dual-functional antioxidant. Food Chem. 2018, 246, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.C.; Vestner, J.; Barril, C.; Maury, C.; Prenzler, P.D.; Scollary, G.R. The influence of stereochemistry of antioxidants and flavanols on oxidation processes in a model wine system: Ascorbic acid, erythorbic acid, (+)-catechin and (-)-epicatechin. J. Agric. Food Chem. 2009, 58, 1004–1011. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Kao, M.C.; Fang, J.Y.; Zouboulis, C.C.; Zhang, L.; Gallo, R.L.; Huang, C.M. Antimicrobial property of lauric acid against Propionibacterium acnes: Its therapeutic potential for inflammatory acne vulgaris. J. Investig. Dermatol. 2009, 129, 2480–2488. [Google Scholar] [CrossRef] [Green Version]
- Park, K.M.; Lee, M.J.; Jo, S.K.; Choi, S.J.; Lee, J.; Chang, P.S. Erythorbyl laurate as a potential food additive with multi-functionalities: Interfacial characteristics and antioxidant activity. Food Chem. 2017, 215, 101–107. [Google Scholar] [CrossRef]
- Ifuku, S. Chitin and chitosan nanofibers: Preparation and chemical modifications. Molecules 2014, 19, 18367–18380. [Google Scholar] [CrossRef]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; De los Llanos Gandia, M.; Caballero, A.H. Cosmetics and Cosmeceutical Applications of Chitin, Chitosan and Their Derivatives. Polymers 2018, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Luan, F.; Wei, L.; Zhang, J.; Tan, W.; Chem, Y.; Dong, F.; Li, Q.; Guo, Z. Preparation and Characterization of Quaternized Chitosan Derivatives and Assessment of Their Antioxidant Activity. Molecules 2018, 23, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, O.S.; Lucas, R.; Cornelles, F.; Gonzalez, M.J.; Parra, J.S.; Medina, I.; Morales, J.C. Synthesis and characterization of phenolic antioxidants with surfactant properties: Glucosyl- and glucuronosyl alkyl gallates. Tetrahedron 2011, 67, 7268–7279. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, W.; Li, X.; Lu, X.; Li, N.; Gao, X.; Song, J. Preparation and in vitro antioxidant activity of kappa-carrageenan oligosaccharides and their oversulfated, acetylated, and phosphorylated derivatives. Carbohydr. Res. 2005, 340, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.W.; Li, J.; Wang, W.; Guan, H.S. Protective effects of kappa-ca3000+CP against ultraviolet-induced damage in HaCaT and MEF cells. J. Photochem. Photobiol. B 2010, 101, 22–30. [Google Scholar] [CrossRef]
- Haque, E.; Kayalvizhi, K.; Hassan, S. Biocompatibility, Antioxidant and Anti-Infective Effect of Biosurfactant Produced by Marinobacter litoralis MB15. Int. J. Pharm. Investig. 2020, 10, 172–177. [Google Scholar] [CrossRef]
- Abdollahi, S.; Tofighi, Z.; Babaee, T.; Shamsi, M.; Rahimzadeh, G.; Rezvanifar, H.; Saeidi, E.; Mohajeri Amiri, M.; Saffari Ashtiani, Y.; Samadi, N. Evaluation of Anti-oxidant and Anti-biofilm Activities of Biogenic Surfactants Derived from Bacillus amyloliquefaciens and Pseudomonas aeruginosa. Iran. J. Pharm. Res. 2020, in press. [Google Scholar]
- Liu, F.; Zhu, Z.; Ma, C.; Luo, X.; Bai, L.; Decker, E.A.; Gao, Y.; McClements, D.J. Fabrication of concentrated fish oil emulsions using dual-channel microfluidization: Impact of droplet concentration on physical properties and lipid oxidation. J. Agric. Food Chem. 2016, 64, 9532–9541. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.K.; Asif, M.; Nazari, M.V.; Baharetha, H.M.; Mahmood, S.; Yatim, A.R.M.; Majid, A.S.A.; Majid, A.M.S.A. Antiangiogenic activity of sophorolipids extracted from refined bleached deodorized palm olein. Indian J. Pharmacol. 2019, 51, 45. [Google Scholar]
- Takahashi, M.; Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Glycolipid biosurfactants, mannosylerythritol lipids, show antioxidant and protective effects against H2O2-induced oxidative stress in cultured human skin fibroblasts. J. Oleo Sci. 2012, 61, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.P.; Sasse, F.; Bronstrup, M.; Diez, J.; Meyerhans, A. Antiviral Drug Discovery: Broad-Spectrum Drugs From Nature. Nat. Prod. Rep. 2015, 32, 29–48. [Google Scholar] [CrossRef]
- Kreiss, J.; Ngugi, E.; Holmes, K.; Ndinya-Achola, J.; Waiyaki, P.; Roberts, P.L.; Ruminjo, I.; Sajabi, R.; Kimata, J.; Fleming, T.R.; et al. Efficacy of nonoxynol 9 contraceptive sponge use in preventing heterosexual acquisition of HIV in Nairobi prostitutes. JAMA 1992, 268, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Roddy, R.E.; Cordero, M.; Ryan, K.A.; Figueroa, J. A randomized controlled trial comparing nonoxynol-9 lubricated condoms with silicone lubricated condoms for prophylaxis. Sex Transm. Infect. 1998, 74, 116–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leydet, A.; El Hachemi, H.; Boyer, B.; Lamaty, G.; Roque, J.P.; Schols, D.; Snoeck, R.; Andrei, G.; Ikeda, S.; Neyts, J.; et al. Polyanion inhibitors of human immunodeficiency virus and other viruses. Part 2. Polymerized anionic surfactants derived from amino acids and dipeptides. J. Med. Chem. 1996, 39, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Leydet, A.; Barragan, V.; Boyer, B.; Montéro, J.L.; Roque, J.P.; Witvrouw, M.; Este, J.; Snoeck, R.; Andrei, G.; De Clercq, E. Polyanion inhibitors of human immunodeficiency virus and other viruses. 5. Telomerized anionic surfactants derived from amino acids. J. Med. Chem. 1997, 40, 342–349. [Google Scholar] [CrossRef]
- Howett, M.K.; Neely, E.B.; Christensen, N.D.; Wigdahl, B.; Krebs, F.C.; Malamud, D.; Patrick, S.D.; Pickel, M.D.; Welsh, P.A.; Reed, C.A.; et al. A broad-spectrum microbicide with virucidal activity against sexually transmitted viruses. Antimicrob. Agents Chemother. 1999, 43, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Piret, J.; Désormeaux, A.; Bergeron, M.G. Sodium lauryl sulfate, a microbicide effective against enveloped and nonenveloped viruses. Curr. Drug Targets 2002, 3, 17–30. [Google Scholar] [CrossRef]
- Bailey, A.; Longson, M. Virucidal activity of chlorhexidine on strains of Herpesvirus hominis, poliovirus, and adenovirus. J. Clin. Pathol. 1972, 25, 76–78. [Google Scholar] [CrossRef] [Green Version]
- Jenabian, M.A.; Auvert, B.; Saïdi, H.; Lissouba, P.; Matta, M.; Bélec, L. Potent in vitro inactivation of both free and cell-associated CCR5- and CXCR4-tropic HIV-1 by common commercial soap bars from South Africa. J. Acquir. Immune Defic. Syndr. 2010, 54, 340–342. [Google Scholar] [CrossRef]
- Alkhatib, M.H.; Aly, M.M.; Rahbeni, R.A.; Balamash, K.S. Antimicrobial Activity of Biocompatible Microemulsions Against Aspergillus niger and Herpes Simplex Virus Type 2. Jundishapur J. Microbiol. 2016, 9, e37437. [Google Scholar] [CrossRef] [Green Version]
- Chepurnov, A.A.; Bakulina, L.F.; Dadaeva, A.A.; Ustinova, E.N.; Chepurnova, T.S.; Baker, J.R., Jr. Inactivation of Ebola virus with a surfactant nanoemulsion. Acta Trop. 2003, 87, 315–320. [Google Scholar] [CrossRef]
- Jefferson, T.; Jones, M.A.; Doshi, P.; Del Mar, C.B.; Hama, R.; Thompson, M.J.; Spencer, E.A.; Onakpoya, I.; Mahtani, K.R.; Nunan, D.; et al. Neuraminidase inhibitors for preventing and treating influenza in healthy adults and children. Cochrane Database Syst. Rev. 2014, 4, CD008965. [Google Scholar] [CrossRef] [Green Version]
- Greatorex, J.S.; Digard, P.; Curran, M.D.; Moynihan, R.; Wensley, H.; Wreghitt, T.; Varsani, H.; Garcia, F.; Enstone, J.; Nguyen-Van-Tam, J.S. Survival of influenza A(H1N1) on Materials Found in Households: Implications for Infection Control. PLoS ONE 2011, 6, e27932. [Google Scholar] [CrossRef] [PubMed]
- Botequim, D.; Maia, J.; Lino, M.M.; Lopes, L.M.; Simoes, P.N.; IIharco, L.M.; Ferreira, L. Nanoparticles and surfaces presenting antifungal, antibacterial and antiviral properties. Langmuir 2012, 28, 7646–7656. [Google Scholar] [CrossRef] [PubMed]
- Donovan, B.W.; Reuter, J.D.; Cao, Z.; Myc, A.; Johnson, K.J.; Baker, J.R., Jr. Prevention of murine influenza A virus pneumonitis by surfactant nano-emulsions. Antivir. Chem. Chemother. 2000, 11, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimolai, N. Environmental and Decontamination Issues for Human Coronaviruses and Their Potential Surrogates. J. Med.Virol. 2020, 1–13. [Google Scholar] [CrossRef]
- Predmore, A.; Li, J. Enhanced removal of a human norovirus surrogate from fresh vegetables and fruits by a combination of surfactants and sanitizers. Appl. Environ. Microbiol. 2011, 77, 4829–4838. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Zuber, S.; Cantergiani, F.; Butot, S.; Li, D.; Stroheker, T.; Devlieghere, F.; Lima, A.; Piantini, U.; Uyttendaele, M. Inactivation of viruses and bacteria on strawberries using a levulinic acid plus sodium dodecyl sulfate based sanitizer, taking sensorial and chemical food safety aspects into account. Int. J. Food Microbiol. 2017, 257, 176–182. [Google Scholar] [CrossRef]
- Mertens, B.S.; Velev, O.D. Characterization and control of surfactant-mediated Norovirus interactions. Soft Matter 2015, 11, 8621–8631. [Google Scholar] [CrossRef] [Green Version]
- Steinmann, J.; Paulmann, D.; Becker, B.; Bischoff, B.; Steinmann, E.; Steinmann, J. Comparison of virucidal activity of alcohol-based hand sanitizers versus antimicrobial hand soaps in vitro and in vivo. J. Hosp. Infect. 2012, 82, 277–280. [Google Scholar] [CrossRef]
- Lages, S.L.; Ramakrishnan, M.A.; Goyal, S.M. In-vivo efficacy of hand sanitisers against feline calicivirus: A surrogate for norovirus. J. Hosp. Infect. 2008, 68, 159–163. [Google Scholar] [CrossRef]
- Liu, P.; Yuen, Y.; Hsiao, H.M.; Jaykus, L.A.; Moe, C. Effectiveness of liquid soap and hand sanitizer against Norwalk virus on contaminated hands. Appl. Environ. Microbiol. 2009, 76, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Tuladhar, E.; Hazeleger, W.C.; Koopmans, M.; Zwietering, M.H.; Duizer, E.; Beumer, R.R. Reducing viral contamination from finger pads: Handwashing is more effective than alcohol-based hand disinfectants. J. Hosp. Infect. 2015, 90, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Remichkova, M.; Galabova, D.; Roeva, I.; Karpenko, E.; Shulga, A.; Galabov, A.S. Anti-herpesvirus activities of Pseudomonas sp. S-17 rhamnolipid and its complex with alginate. Z. Naturforsch. C 2008, 63, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Doncel, G.F.; Seyoum, T.; Eaton, K.M.; Zalenskaya, I.; Hagver, R.; Azim, A.; Gross, R. Sophorolipids, microbial glycolipids with anti-human immunodeficiency virus and sperm-immobilizing activities. Antimicrob. Agents Chemother. 2005, 49, 4093–4100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, R.A.; Shah, V. Anti-Herpes Virus Properties of various Forms of Sophorolipids. Patent WO2007130738 A1, 15 November 2007. [Google Scholar]
- Gross, R.A.; Shah, V.; Doncel, G. Virucidal Properties of Various Forms of Sophorolipids. Patent US8648055:B2, 11 February 2014. [Google Scholar]
- Fernando, I.P.S.; Nah, J.W.; Jeon, Y.J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- Mason, R.J. Thoughts on the alveolar phase of COVID-19. AJP Lung Cell. Mol. Phys. 2020, 319, L115–L120. [Google Scholar] [CrossRef]
- Hite, R.D.; Morris, P.E. Acute respiratory distress syndrome: Pharmacological treatment options in development. Drugs 2001, 61, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, F.; Pugliese, F.; Ranieri, V.M. The Role of Rescue Therapies in the Treatment of Severe ARDS. Respir. Care 2018, 63, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Glaser, K.; Fehrholz, M.; Papsdorf, M.; Curstedt, T.; Kunzmann, S.; Speer, C.P. The new generation synthetic reconstituted surfactant CHF5633 suppresses LPS-induced cytokine responses in human neonatal monocytes. Cytokine 2016, 86, 119–123. [Google Scholar] [CrossRef]
- Glaser, K.; Fehrholz, M.; Curstedt, T.; Kunzmann, S.; Speer, C.P. Effects of the New Generation Synthetic Reconstituted Surfactant CHF5633 on Pro- and Anti-Inflammatory Cytokine Expression in Native and LPS-Stimulated Adult CD14+ Monocytes. PLoS ONE 2016, 11, e0146898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrholz, M.; Glaser, K.; Seidenspinner, S.; Ottensmeier, B.; Curstedt, T.; Speer, C.P.; Kunzmann, S. Impact of the New Generation Reconstituted Surfactant CHF5633 on Human CD4+ Lymphocytes. PLoS ONE 2016, 11, e0153578. [Google Scholar] [CrossRef] [PubMed]
- Kuronuma, K.; Mitsuzawa, H.; Takeda, K.; Nishitani, C.; Chan, E.D.; Kuroki, Y.; Nakamura, M.; Voelker, D.R. Anionic pulmonary surfactant phospholipids inhibit inflammatory responses from alveolar macrophages and U937 cells by binding the lipopolysaccharide-interacting proteins CD14 and MD-2. J. Biol. Chem. 2009, 284, 25488–25500. [Google Scholar] [CrossRef] [Green Version]
- Otsubo, E.; Irimajiri, K.; Takei, T.; Nomura, M. Characterization of synthetic lung surfactant activity against proinflammatory cytokines in human monocytes. Biol. Pharm. Bull. 2002, 25, 312–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemhöner, A.; Rüdiger, M.; Gortner, L. Inflammatory cytokine mRNA in monocytes is modified by a recombinant (SP-C)-based surfactant and porcine surfactant. Methods Find Exp. Clin. Pharmacol. 2009, 31, 317–323. [Google Scholar] [CrossRef]
- Backhaus, S.; Zakrzewicz, A.; Richter, K.; Damm, J.; Wilker, S.; Fuchs-Moll, G.; Küllmar, M.; Hecker, A.; Manzini, I.; Ruppert, C.; et al. Surfactant inhibits ATP-induced release of interleukin-1β via nicotinic acetylcholine receptors. J. Lipid. Res. 2017, 58, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.; Chang, K.J.; Park, S.Y.; Um, J.H.; Han, E.J.; Lee, W.; Kim, E.A.; Oh, J.Y.; Kim, S.H.; Jeon, Y.J.; et al. Anti-inflammatory Effects of Galactose-Taurine Sodium Salt in LPS-Activated RAW 264.7 Cells. Adv. Exp. Med. Biol. 2017, 975, 943–953. [Google Scholar]
- Mittal, N.; Sanyal, S.N. In vivo effect of surfactant on inflammatory cytokines during endotoxin-induced lung injury in rodents. J. Immunotoxicol. 2011, 8, 274–283. [Google Scholar] [CrossRef]
- Maeng, Y.; Kim, K.T.; Zhou, X.; Jin, L.; Kim, K.S.; Kim, Y.H.; Lee, S.; Park, J.H.; Chen, X.; Kong, M.; et al. A novel microbial technique for producing high-quality sophorolipids from horse oil suitable for cosmetic applications. Microb. Biotechnol. 2018, 11, 917–929. [Google Scholar] [CrossRef]
- Hagler, M.; Smith-Norowitz, T.A.; Chice, S.; Wallner, S.R.; Viterbo, D.; Mueller, C.M.; Gross, R.; Nowakowski, M.; Schulze, R.; Zenilman, M.E.; et al. Sophorolipids decrease IgE production in U266 cells by downregulation of BSAP (Pax5), TLR-2, STAT3 and IL-6. J. Allergy Clin. Immun. 2007, 119, S263. [Google Scholar] [CrossRef]
- Morita, Y.; Tadokoro, S.; Sasai, M.; Kitamoto, D.; Hirashima, N. Biosurfactant mannosyl-erythritol lipid inhibits secretion of inflammatory mediators from RBL-2H3 cells. BBA Gen. Subj. 2011, 1810, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour, S.H.; Dehghani, M. Review of cancer from perspective of molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.G.; Juárez-Portilla, C.; Olivares-Bañuelos, T.; Zepeda, R.C. Anticancer activity of seaweeds. Drug Discov. Today 2018, 23, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Abdelnour, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Gebriel, M.G. Microalgae in modern cancer therapy: Current knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, R.; Suzuki, C.; Ohno, M.; Ohasi, T.; Futagami, R.; Ishikawa, K.; Komae, M.; Nishino, T.; Konishi, Y.; Lee, E. Cationic Surfactants Induce Apoptosis in Normal and Cancer Cells. Ann. N. Y. Acad. Sci. 2007, 1095, 1–6. [Google Scholar] [CrossRef]
- Yip, K.W.; Mao, X.; Au, P.Y.B.; Hedley, D.W.; Chow, S.; Dalili, S.; Mocanu, J.D.; Bastianutto, C.; Schimmer, A.; Liu, F.F. Benzethonium Chloride: A Novel Anticancer Agent Identified by Using a Cell-Based Small-Molecule Screen. Clin. Cancer Res. 2006, 12, 5557–5569. [Google Scholar] [CrossRef] [Green Version]
- Rayess, H.M.; Xi, Y.; Garshott, D.M.; Brownell, A.L.; Yoo, G.H.; Callaghan, M.U.; Fribley, A.M. Benzethonium chloride activates ER stress and reduces proliferation in HNSCC. Oral Oncol. 2018, 76, 27–33. [Google Scholar] [CrossRef]
- Zerin, T.; Song, H.; Gil, H.; Hong, S. Surfactant 4-nonylphenyl-polyethylene glycol stimulates reactive oxygen species generation and apoptosis in human neuroblastoma cells. J. Environ. Sci. 2017, 53, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Busto, J.V.; Sot, J.; Goñi, F.M.; Mollinedo, F.; Alonso, A. Surface-active properties of the anti-tumor ether lipid 1-O-octadecyl-2-O-methyl-rac-glycero-3-phosphocholine (edelfosine). Biochim. Biophys. Acta 2007, 1768, 1855–1860. [Google Scholar] [CrossRef] [PubMed]
- Gajate, C.; Santos-Beneit, A.M.; Macho, A.; Lazaro, M.d.; Hernandez-De Rojas, A.; Modolell, M.; Munoz, E.; Mollinedo, F. Involvement of mitochondria and caspase-3 in ET-18-OCH3-induced apoptosis of human leukemic cells. Int. J. Cancer 2000, 86, 208–218. [Google Scholar] [CrossRef]
- Gajate, C.; Mollinedo, F. Edelfosine and perifosine induce selective apoptosis in multiple myeloma by recruitment of death receptors and downstream signaling molecules into lipid rafts. Blood 2007, 109, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Gajate, C.; del Canto-Jañez, E.; Acuña, A.U.; Amat-Guerri, F.; Geijo, E.; Santos-Beneit, A.M.; Veldman, R.J.; Mollinedo, F. Intracellular Triggering of Fas Aggregation and Recruitment of Apoptotic Molecules into Fas-enriched Rafts in Selective Tumor Cell Apoptosis. J. Exp. Med. 2004, 200, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Mollinedo, F.; de la Iglesia-Vicente, J.; Gajate, C.; Estella-Hermoso de Mendoza, A.; Villa-Pulgarin, J.A.; Campanero, M.A.; Blanco-Prieto, M.J. Lipid raft-targeted therapy in multiple myeloma. Oncogene 2010, 29, 3748–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.C.; Parajuli, K.R.; Han, S.I. The alkyllysophospholipidedelfosine enhances TRAIL-mediated apoptosis in gastric cancer cells through death receptor 5 and the mitochondrial pathway. Tumor Biol. 2016, 37, 6205–6216. [Google Scholar] [CrossRef]
- Melo-Lima, S.; Lopes, M.C.; Mollinedo, F. ERK1/2 acts as a switch between necrotic and apoptotic cell death in ether phospholipid edelfosine-treated glioblastoma cells. Pharmacol. Res. 2015, 95–96, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Na, H.K.; Surh, Y.J. The antitumor ether lipid edelfosine (ET-18-O-CH3) induces apoptosis in H-ras transformed human breast epithelial cells: By blocking ERK1/2 and p38 mitogen- activated protein kinases as potential targets. Asia Pac. J. Clin. Nutr. 2008, 17, 204–207. [Google Scholar] [PubMed]
- Udayakumar, T.S.; Stoyanova, R.; Shareef, M.M.; Mu, Z.; Philip, S.; Burnstein, K.L.; Pollack, A. Edelfosine Promotes Apoptosis in Androgen-Deprived Prostate Tumors by Increasing ATF3 and Inhibiting Androgen Receptor Activity. Mol. Cancer Ther. 2016, 15, 1353–1363. [Google Scholar] [CrossRef] [Green Version]
- Estella-Hermoso de Mendoza, A.; Préat, V.; Mollinedo, F.; Blanco-Prieto, M.J. In vitro and in vivo efficacy of edelfosine-loaded lipid nanoparticles against glioma. J. Control. Release 2011, 156, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Steinestel, K.; Eder, S.; Ehinger, K.; Schneider, J.; Genze, F.; Winkler, E.; Wardelmann, E.; Schrader, A.J.; Steinestel, J. The small conductance calcium-activated potassium channel 3 (SK3) is a molecular target for Edelfosine to reduce the invasive potential of urothelial carcinoma cells. Tumor Biol. 2016, 37, 6275–6283. [Google Scholar] [CrossRef]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. Chemical structure and in vitro antitumor activity of rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol. 2013, 170, 676–689. [Google Scholar] [CrossRef]
- Rahimi, K.; Lotfabad, T.B.; Jabeen, F.; Ganji, S.M. Cytotoxic effects of mono-and di-rhamnolipids from Pseudomonas aeruginosa MR01 on MCF-7 human breast cancer cells. Colloid Surf. B 2019, 181, 943–952. [Google Scholar] [CrossRef]
- Thanomsub, B.; Pumeechockchai, W.; Limtrakul, A.; Arunrattiyakorn, P.; Petchleelaha, W.; Nitoda, T.; Kanzaki, H. Chemical structures and biological activities of rhamnolipids produced by Pseudomonas aeruginosa B189 isolated from milk factory waste. Bioresour. Technol. 2015, 97, 2457–2461. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, X.; Zhang, H.; Qu, Y. Production, structure elucidation and anticancer properties of sophorolipid from Wickerhamiella domercqiae. Enzyme Microb. Technol. 2006, 39, 501–506. [Google Scholar] [CrossRef]
- Ribeiro, I.A.; Faustino, C.M.; Guerreiro, P.S.; Frade, R.F.; Bronze, M.R.; Castro, M.F.; Ribeiro, M.H. Development of novel sophorolipids with improved cytotoxic activity toward MDA-MB-231 breast cancer cells. J. Mol. Recognit. 2015, 28, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Song, X.; Ma, X.; Li, H.; Qu, Y. Bioactivities of sophorolipid with different structures against human esophageal cancer cells. J. Surg. Res. 2012, 173, 286–291. [Google Scholar] [CrossRef]
- Nawale, L.; Dubey, P.; Chaudhari, B.; Sarkar, D.; Prabhune, A. Anti-proliferative effect of novel primary cetyl alcohol derived sophorolipids against human cervical cancer cells HeLa. PLoS ONE 2017, 12, e0174241. [Google Scholar] [CrossRef]
- Li, H.; Guo, W.; Ma, X.J.; Li, J.S.; Song, X. In vitro and in vivo anticancer activity of sophorolipids to human cervical cancer. Appl. Biochem. Biotechnol. 2017, 181, 1372–1387. [Google Scholar] [CrossRef]
- Fu, S.L.; Wallner, S.R.; Bowne, W.B.; Hagler, M.D.; Zenilman, M.E.; Gross, R.; Bluth, M.H. Sophorolipids and their derivatives are lethal against human pancreatic cancer cells. J. Surg. Res. 2008, 148, 77–82. [Google Scholar] [CrossRef]
- Joshi-Navare, K.; Shiras, A.; Prabhune, A. Differentiation-inducing ability of sophorolipids of oleic and linoleic acids using a glioma cell line. Biotechnol. J. 2011, 6, 509–512. [Google Scholar] [CrossRef]
- Coelho, A.L.S.; Feuser, P.E.; Carciofi, B.A.M.; de Andrade, C.J.; de Oliveira, D. Mannosylerythritol lipids: Antimicrobial and biomedical properties. Appl. Microbiol. Biot. 2020, 104, 2297–2318. [Google Scholar] [CrossRef]
- Isoda, H.; Kitamoto, D.; Shinmoto, H.; Matsumura, M.; Nakahara, T. Microbial extracellular glycolipid induction of differentiation and inhibition of the protein kinase C activity of human promyelocytic leukemia cell line HL-60. Biosci. Biotechnol. Biochem. 1997, 61, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Isoda, H.; Nakahara, T. Mannosylerythritol lipid induces granulocytic differentiation and inhibits the tyrosine phosphorylation of human myelogenous leukemia cell line K562. Cytotechnology 1997, 25, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M. Amphiphilic glycoconjugates as potential anticancer chemotherapeutics. Eur. J. Med. Chem. 2018, 143, 1208–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wakamatsu, Y.; Shibahara, M.; Nomura, N.; Geltinger, C.; Nakahara, T.; Murata, T.; Yokoyama, K.K. Mannosylerythritol lipid is a potent inducer of apoptosis and differentiation of mouse melanoma cells in culture. Cancer Res. 1999, 59, 482–486. [Google Scholar] [PubMed]
- Zhao, X.; Murata, T.; Ohno, S.; Day, N.; Song, J.; Nomura, N.; Nakahara, T.; Yokoyama, K.K. Protein kinase Cα plays a critical role in mannosylerythritol lipid-induced differentiation of melanoma B16 Cells. J. Biol. Chem. 2001, 276, 39903–39910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakur, A.; Niu, Y.; Kuang, H.; Chen, Q. Synthesis of gold nanoparticles derived from mannosylerythritol lipid and evaluation of their bioactivities. AMB Express 2019, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Bakur, A.; Elshaarani, T.; Niu, Y.; Chen, Q. Comparative study of antidiabetic, bactericidal, and antitumor activities of MEL@AgNPs, MEL@ZnONPs, and Ag–ZnO/MEL/GA nanocomposites prepared by using MEL and gum arabic. RSC Adv. 2019, 9, 9745–9754. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Samoski, E.A.; Acar, M. The Systems Biology of Single-Cell Aging. iScience 2018, 7, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campisi, J. Aging, cellular senescence, and cancer. Annu. Rev. Physiol. 2013, 75, 685–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Tollefsbol, T. Telomeres and telomerase: Basic science implications for aging. J. Am. Geriatr. Soc. 2001, 49, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Depinho, R.A. Linking functional decline of telomeres, mitochondria and stem cells during ageing. Nature 2010, 464, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riera, C.E.; Merkwirth, C.; De Magalhaes Filho, C.D.; Dillin, A. Signaling networks determining life span. Annu. Rev. Biochem. 2016, 85, 35–64. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.W.; Chen, Y.C.; Hsieh, W.L.; Chiou, S.H.; Kao, C.L. Ageing and neurodegenerative diseases. Ageing Res. Rev. 2010, 9, S36–S46. [Google Scholar] [CrossRef]
- Kyriakakis, E.; Princz, A.; Tavernarakis, N. Stress responses during ageing: Molecular pathways regulating protein homeostasis. Methods Mol. Biol. 2015, 1292, 215–234. [Google Scholar]
- De Almeida, A.J.P.O.; Ribeiro, T.P.; de Medeiros, I.A. Aging: Molecular pathways and implications on the cardiovascular system. Oxid. Med. Cell Longev. 2017, 2017, 7941563. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Functional and physiological characteristics of the aging skin. Aging Clin. Exp. Res. 2008, 20, 195–200. [Google Scholar] [CrossRef]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Markantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Dermatoendocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Mailbach, H.I. Characteristics of the skin aging. Adv. Wound Care (New Rochelle) 2013, 2, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Weihermann, A.C.; Lorencini, M.; Brohem, C.A.; de Carvalho, C.M. Elastin structure and its involvement in skin photoageing. Int. J. Cosmet. Sci. 2017, 39, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G. Molecular mechanisms of skin ageing. Mech. Ageing Dev. 2002, 123, 801–810. [Google Scholar] [CrossRef]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Chaikul, P.; Khat-Udomkiri, N.; Iangthnarat, K.; Manosroi, J.; Manosroi, A. Characteristics and in vitro anti-skin aging activity of gallic acid loaded in cationic CTABniosome. Eur. J. Pharm. Sci. 2019, 131, 39–49. [Google Scholar] [CrossRef]
- Ortonne, J.P. Photoprotective properties of skin melanin. Br. J. Dermatol. 2002, 61, 7–10. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Pittayapruek, P.; Meephansan, I.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.C.; Young, D.A.; Waters, J.G.; Rowan, A.D.; Chantry, A.; Edwards, D.R.; Clark, I.M. The comparative role of activator protein 1 and Smad factors in the regulation of Timp-1 and MMP-1 gene expression by transforming growth factor-beta 1. J. Biol. Chem. 2003, 278, 10304–10313. [Google Scholar] [CrossRef] [Green Version]
- Sardy, M. Role of matrix metalloproteinases in skin ageing. Connect. Tissue Res. 2009, 50, 132–138. [Google Scholar] [CrossRef]
- Wen, K.C.; Fan, P.C.; Tsai, S.Y.; Shih, I.C.; Chiang, H.M. Ixora parviflora protects against UVB-induced photoaging by inhibiting the expression of MMPs, MAP kinases and COX-2 and by promoting type I procollagen synthesis. Evid. Based Complement Altern. Med. 2012, 2012, 417346. [Google Scholar] [CrossRef] [Green Version]
- Poon, F.; Kang, S.; Chien, A.I. Mechanisms and treatments of photoaging. Photodermatol. Photoimmunol. Photomed. 2015, 31, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Corinaldsesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine microbial-derived molecules and their potential use in cosmeceutical and cosmetic products. Mar. Drugs 2017, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Rajapakse, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydr. Polym. 2005, 62, 357–368. [Google Scholar] [CrossRef]
- Kim, J.A.; Ahn, B.N.; Kong, C.S.; Park, S.H.; Park, B.J.; Kim, S.K. Antiphotoaging effect of chitoligosaccharides on human dermal fibroblasts. Photodermatol. Photoimmunol. Photomed. 2012, 28, 299–306. [Google Scholar] [CrossRef]
- Van Deursen, J.M. The role of senescent cells in aging. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Ito, I.; Osaki, T.; Ifuku, S.; Saimoto, H.; Takamori, Y.; Kurozumi, S.; Inagawa, T.; Azuma, K.; Tsuka, T.; Okamoto, Y.; et al. Evaluation of the effects of chitin nanofibrils on skin function using skin models. Carbohydr. Polym. 2014, 101, 464–470. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, S.Z.; Li, J.C.; Li, S.D.; Liao, M.N.; Li, C.P.; Zheng, P.J.; Guo, M.H.; Tan, W.X.; Zheng, Z.H.; Hu, Z. Anti-aging effect of chitosan oligosaccharide on d-galactose-induced subacute aging in mice. Mar. Drugs 2018, 16, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piljac, T.; Piljac, G. Use of Rhamnolipids in Wound Healing, Treating Burn Shock, Atherosclerosis, Organ Transplants, Depression, Schizophrenia and Cosmetics. Patent WO1999043334 A1, 2 September 1999. [Google Scholar]
- Kitamoto, D.; Morita, T.; Fukuoka, T.; Konishi, M.A.; Imura, T. Self-assembling properties of glycolipid biosurfactants and their potential applications. Curr. Opin. Colloid 2009, 14, 315–328. [Google Scholar] [CrossRef]
- Yamamoto, S.; Morita, T.; Fukuoka, T.; Imura, T.; Yanagidani, S.; Sogabe, A.; Kitamoto, D.; Kitagawa, M. The moisturizing effects of glycolipid biosurfactants, mannosylerythritol lipids, on human skin. J. Oleo Sci. 2012, 61, 407–412. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Polymer | Pathogen | MIC | MBIC | References |
|---|---|---|---|---|
| Hexameric with amide moieties (PAHB) | E. coli | 0.93 μΜ | NA | [8] |
| Alkylbenzyldimethylammonium chlorides (BAC) analogues | S. aureus | 2.6–10.6 μΜ | NA | [12] |
| E. faecalis | 3.5–10.6 μΜ | NA | ||
| E. coli | 27.9–123.7 μΜ | NA | ||
| P.aeruginosa | 223.1–247.5 μΜ | NA | ||
| C. albicans | 13.9–123.7 μΜ | NA | ||
| Clorexhidine | S. anginosus | 0.97–3.91 μg/mL | 15.63–31.25 μg/mL | [30] |
| S. constellatus | 0.97–3.91 μg/mL | 7.81–31.25 μg/mL | ||
| S. gordonii | 1.95–7.81 μg/mL | 7.81–31.25 μg/mL | ||
| S. mitis | 1.95–15.63 μg/mL | 3.91–31.25 μg/mL | ||
| S. mutans | 0.12–0.97 μg/mL | 15.63–31.25 μg/mL | ||
| S. oralis | 0.97–7.81 μg/mL | 3.91–15.63 μg/mL | ||
| S. salivarius | 0.48–3.91 μg/mL | 1.95–15.63 μg/mL | ||
| S. sanguinis | 1.95–3.91 μg/mL | 7.81–31.25 μg/mL | ||
| Cetylpyridium chloride | S. anginosus | 0.12–1.95 μg/mL | 3.91–15.63 μg/mL | |
| S. constellatus | 0.06–0.48 μg/mL | 3.91–31.25 μg/mL | ||
| S. gordonii | 0.12–0.97 μg/mL | 3.91–31.25 μg/mL | ||
| S. mitis | 0.12–0.97 μg/mL | 3.91–31.25 μg/mL | ||
| S. mutans | 0.06–0.48 μg/mL | 3.91–31.25 μg/mL | ||
| S. oralis | 0.12–1.95 μg/mL | 3.91–31.25 μg/mL | ||
| S. salivarius | 0.06–0.48 μg/mL | 3.91–15.63 μg/mL | ||
| S. sanguinis | 0.12–0.48 μg/mL | 3.91–31.25 μg/mL | ||
| Peptide gemini (PG)surfactants | E. coli | 8–512 μg/mL | NA | [15] |
| S. typhimurium | 8–512 μg/mL | NA | ||
| S. aureus | 4–512μg/mL | NA | ||
| B. subtilis | 4–512 μg/mL | NA | ||
| Na-lauroyl-arginine methyle ester-derived cationic double-chain surfactants (LANHCx) | M. luteus ATCC 9341 | 17–243 μΜ | NA | [14] |
| B. subtilis ATCC9341 | 19–486 μΜ | NA | ||
| S. aureus ATCC29213 | 17–486 μΜ | NA | ||
| S. epiermidis ATCC12228 | 17–510 μΜ | NA | ||
| P. aeruginosa ATCC27853 | 68–78 μΜ | NA | ||
| E. coli ATCC25922 | 38–269 μΜ | NA | ||
| K. pneumonea ATCC 13883 | 78 μΜ | NA | ||
| C. albicans ATCC10231 | 19–243 μΜ | NA | ||
| C. albicans | 35–436 μΜ | NA | ||
| P. aeruginosa | 218–459 μΜ | NA | ||
| K. pneumonieae | 218–459 μΜ | NA | ||
| E. coli | 166–331 μΜ | NA | ||
| Gemini alanine-derived ammonium salts (bromides and chlorides) | C. cerevisiae | 40–1200 μΜ | NA | [34] |
| C.albicans | 80–1200 μΜ | NA | ||
| Rhodotorulamucilaginosa | 10–1200 μΜ | NA | ||
| Rhamnolipid (RL) | P. aeruginosa PAO1 | NA | NA | [16] |
| E. coli NCTC10418 | NA | NA | ||
| B. subtilis NCTC10400 | 0.5% (w/v) | NA | ||
| S. aureus ATCC9144 | 0.5% (w/v) | NA | ||
| E. coli | 12–16 mg/mL | NA | [17] | |
| P. aeruginosa | 8–12 mg/mL | NA | ||
| S. aureus | 16–24 mg/mL | NA | ||
| B. cereus | 12–16 mg/mL | NA | ||
| K. pneomoniae | 16–24 mg/mL | NA | ||
| C. albicans | 8–12 mg/mL | NA | ||
| C. krusei | 12–16 mg/mL | NA | ||
| L. monocytogenes | 78.1– >2500 µg/mL | NA | [19] | |
| B. cereus | pH 5 no growth pH 6 no growth pH 7 19.5 µg/mL pH 8 78.1 µg/mL pH 9 156.2 µg/mL | NA | [18] | |
| L. moncytogenes | pH 5 no growth pH 6 19.5 µg/mL pH 7 156.2 µg/mL pH 8 > 2500 µg/mL pH 9 >2500 µg/mL | NA | ||
| S. aureus | pH 5 9.8 µg/mL pH 6 39.1 µg/mL pH 7 > 2500 µg/mL pH 8 > 2500 µg/mL pH 9 > 2500 µg/mL | NA | ||
| E. coli | pH 5 > 2500 µ g/mL pH 6 > 2500 µg/mL pH 7 > 2500 µg/mL pH 8 > 2500 µg/mL pH 9 > 2500 µg/mL | NA | ||
| S. enterica | pH 5 > 2500 µg/mL pH 6 > 2500 µg/mL pH 7 > 2500 µg/mL pH 8 > 2500 µg/mL pH 9 > 2500 µg/mL | NA | ||
| S. mutans | Mixed RL 390 mg/mL MonoRL 780 mg/mL diRL 390 mg/mL | NA | [20] | |
| S. oralis | Mixed RL 97.5 mg/mL monoRL 780 mg/mL diRL 780 mg/mL | NA | ||
| S. sanguinis | Mixed RL 97.5 mg/mL monoRL 780 mg/mL diRL 390 mg/mL | NA | ||
| A. naeslundii | Mixed RL 195 mg/mL monoRL 1560 mg/mL diRL 780 mg/mL | NA | ||
| N. mucosa | Mixed RL 390 mg/mL monoRL 780 mg/mL diRL 390 mg/mL | NA | ||
| MELs | B. Cereus | 1.25 mg/mL | NA | [26] |
| SAAs | Assay | Mechanism of Action | References |
|---|---|---|---|
| SDS (anionic) Tween-20 (non-ionic) CTAB (cationic) surfactants, O/W emulsions | Lipid hydroperoxides | ↓ lipid oxidation | [71] |
| SDS (anionic) Tween-80 (non-ionic) surfactants, O/W emulsions | Propanal/hydroperoxides | ↓ lipid oxidation | [83] |
| Tween-20 (non-ionic) Tween-20 (non-ionic/Span-20 (co-surfactant) mixture, O/W emulsions | oxygen uptake, conjugated dienes (CD) and formation of volatile compounds | ↓ lipid oxidation | [84] |
| Tween-20/Tween-80 (non-ionic) surfactants | DCFH-DA / Flow Cytometry | ↓ ROS productionin PMA/H2O2-treated human neutrophils | [86] |
| D-α-Tocopheryl polyethylene glycol 1000 succinate (TPGS/Vitamin E TPGS) | TBARS | ↓ ROS production | [94] |
| Alkyl succinylatedtyrosol synthetic amphiphilic lipids | DPPH radical scavenging, TBARS | ↓ formation of DPPH radicals ↓ lipid oxidation | [96] |
| Erythorbyl laurate (6-O-lauroyl-erythorbic acid), | TBARS | ↓ lipid oxidation | [99] |
| single N-quaternized (QCS) and double N-diquaternized (DQCS) chitosan | DPPH, hydroxyl, superoxide radicals, reducing power | ↓ formation of DPPH, hydroxyl and superoxide radicals ↑ reducing power | [102] |
| Glucosyl- and glucuronosyl alkyl gallates derivatives | DPPH, FRAP | ↓ formation of DPPH, ↑ rates of reduction of Fe3+ to Fe2+ | [103] |
| κ-carrageenan oligosaccharides, oversulfated (SD) low acetylated (LAD) high acetylated (HAD) phosphorylated (PD) κ-carrageenans | DPPH, superoxide anion, hydroxyl radical scavenging activity | ↓ formation of DPPH, superoxide and hydroxyl radicals | [104] |
| κ-ca3000 + CP complex (κ-carrageenan 3kD+ collagen peptides) | DCFH-DA/FlowCytometry | ↓ ROS production Protection of HaCaT abd MEF cells from UV-induced cell death | [105] |
| Rhamnolipids | DPPH | ↓ formation of DPPH | [106] |
| DPPH, FRAP | ↓ formation of DPPH, ↑ rates of reduction of Fe3+ to Fe2+ | [107] | |
| TBARS, Lipid hydroperoxides | ↓ lipid oxidation | [108] | |
| Sophorolipids | DPPH, FRAP | ↓ formation of DPPH, ↑ rates of reduction of Fe3+ to Fe2+ | [109] |
| MELs | DPPH, superoxide anion scavenging activity | ↓ formation of DPPHand superoxide radicals | [110] |
| SAA | Mechanism of Action | Viral Target | Experimental Model | References |
|---|---|---|---|---|
| SDS/SLS (anionic surfactant) | Membrane disruption and capsid denaturation | HIV 1 and 2, HPV | various cell lines | [116] |
| Membrane disruption and capsid denaturation | HIV 1 and 2, HPV | various cell lines | [117] | |
| HSV 2 | in vivo (hairless mice) | |||
| SDS (anionic surfactant),levulinic acid and sodium hypochlorite | Undefined | MNV, HAV | in vitro | [129] |
| CTAB (cationic surfactant), SDS (anionic surfactant) | Particle disassociation and entrapment in micelles | MNV | in vivo | [130] |
| Tween-20 (non-ionic surfactant), Triton X-100 (non-ionic surfactant) NP-40 (non-ionic surfactant) | Capsid denaturation | MNV | RAW 264.7 murine macrophages | [128] |
| Chlorhexidine (cationic surfactant) | Membrane disruption | Herpes virus | hep-2 and RK13 cells | [118] |
| African soap bars diluted in water | Undefined | Cell-free HIV-1, HIV-1 infected lymphocytes | human PBMCs | [119] |
| Common soaps | Undefined | MNV-1, HuNoV GI.4 and GII.4 | in vivo | [134] |
| Micro-emulsion of Tween-80 (non-ionic surfactant and emulsifier), Span-20 (co-surfactant), ethanol, oil, isopropyl myristate (IPM), and distilled water | Undefined | HSV-2 | vero cells | [120] |
| Nano-emulsion ATB | Undefined | Cell culture medium and surfaces | vero cells | [121] |
| Nano-emulsion of soybean oil, tributyl phosphate AND Triton X-100 (non-ionic surfactant) | Limited infectivity in animals | Murine influenza A virus | in vivo (CD-1 mice) | [125] |
| Nanoparticles coated with DDAB (cationic surfactant) | Structural damage and inactivation | H1N1 influenza strain | MDCK cells | [124] |
| Rhamnolipid | Not defined | HSV 1 and 2 | MDBK cells | [135] |
| Sophorolipid | Membrane disruption | HIV | cell-free HIV inactivation model | [136] |
| SAA | Experimental Model | Mechanism(s) of Action | References |
|---|---|---|---|
| Chito-oligosaccharides 3-5kDa (COS) | UV-A/B-irradiated human dermal fibroblasts (HDF)()UV-A/B-irradiated hairless BALB/c mice D-gal induced subacute aging mice (Kunming mice) | ↓ MMP-1/8/13 ↓ MMP-2/9 ↓AP-1, c-Jun, c-Fos ↑ TIMP-1/2 ↑ procollagen I-III-IV Improvement of macroscopic and histopathological features ↑ total collagen and collagen I ↑CAT, SOD, GSH-Px↓TNF-a, IL-1β, IL-6 Improvement of kidney and liver function ↓ALT, AST, ALP, UA, CREA ↑ CAT, GSH-Px, SOD ↑IgG/IgM | [215,216,217] |
| κ-ca3000 + CP | UV-A/B-irradiated HaCaT and MEF cells | ↓ UV-induced apoptotic death Inhibition of MAPK pathway and ROS production ↓ MMP-1 | [105] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anestopoulos, I.; Kiousi, D.E.; Klavaris, A.; Galanis, A.; Salek, K.; Euston, S.R.; Pappa, A.; Panayiotidis, M.I. Surface Active Agents and Their Health-Promoting Properties: Molecules of Multifunctional Significance. Pharmaceutics 2020, 12, 688. https://doi.org/10.3390/pharmaceutics12070688
Anestopoulos I, Kiousi DE, Klavaris A, Galanis A, Salek K, Euston SR, Pappa A, Panayiotidis MI. Surface Active Agents and Their Health-Promoting Properties: Molecules of Multifunctional Significance. Pharmaceutics. 2020; 12(7):688. https://doi.org/10.3390/pharmaceutics12070688
Chicago/Turabian StyleAnestopoulos, Ioannis, Despoina Eugenia Kiousi, Ariel Klavaris, Alex Galanis, Karina Salek, Stephen R. Euston, Aglaia Pappa, and Mihalis I. Panayiotidis. 2020. "Surface Active Agents and Their Health-Promoting Properties: Molecules of Multifunctional Significance" Pharmaceutics 12, no. 7: 688. https://doi.org/10.3390/pharmaceutics12070688