Targeting the Tumor Microenvironment with Fluorescence-Activatable Bispecific Endoglin/Fibroblast Activation Protein Targeting Liposomes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lipids and Materials Used for Liposome Preparation
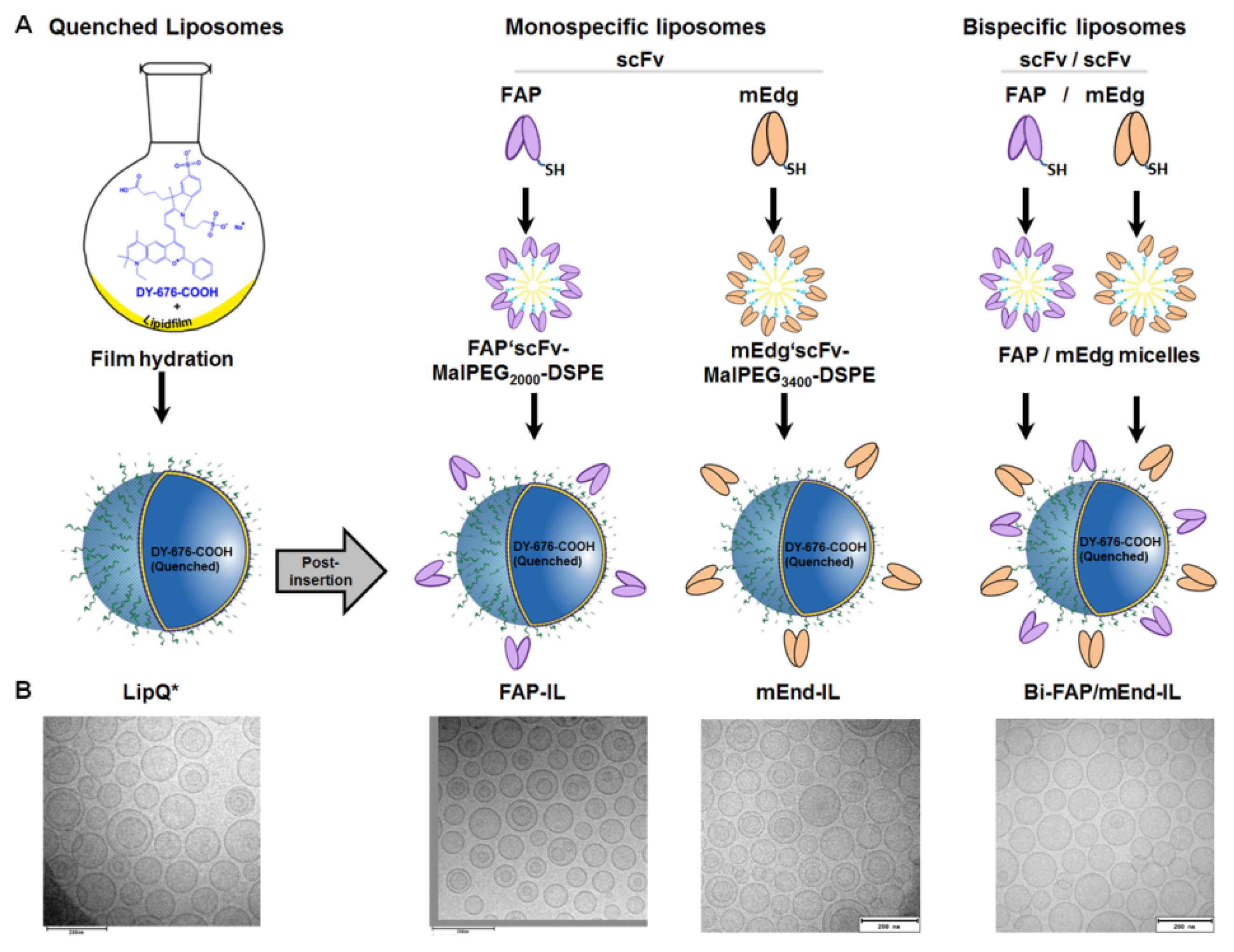
2.2. Preparation and Physicochemical Characterization of Quenched Liposomes
2.3. Preparation and Conjugation of FAP and Murine Endoglin Antibody Fragments to Preformed Quenched Liposomes
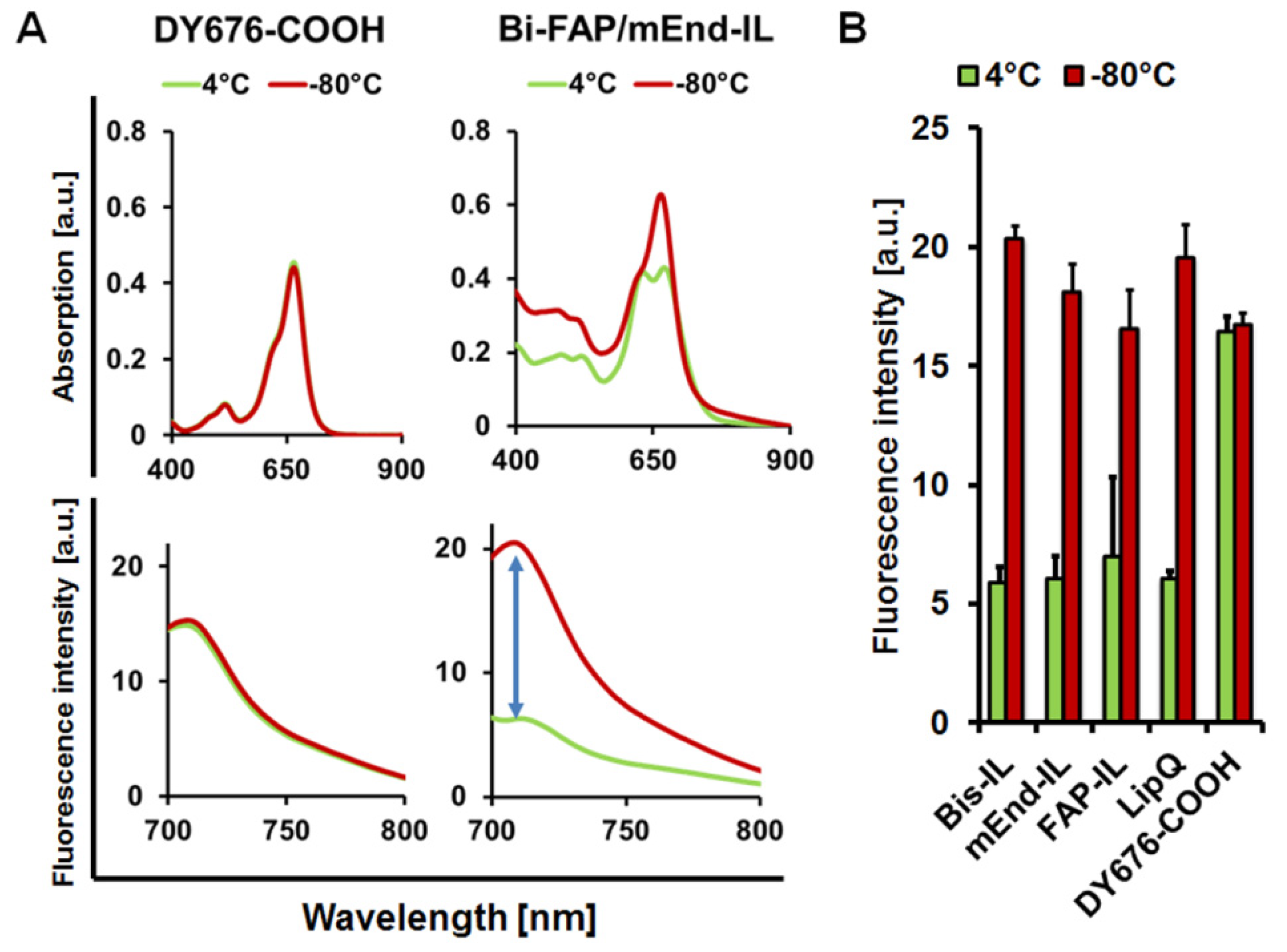
2.4. Determination of the Liposomal Fluorescence Quenching and Activation Property
2.5. Cell Lines, Media and Culture Conditions Implemented
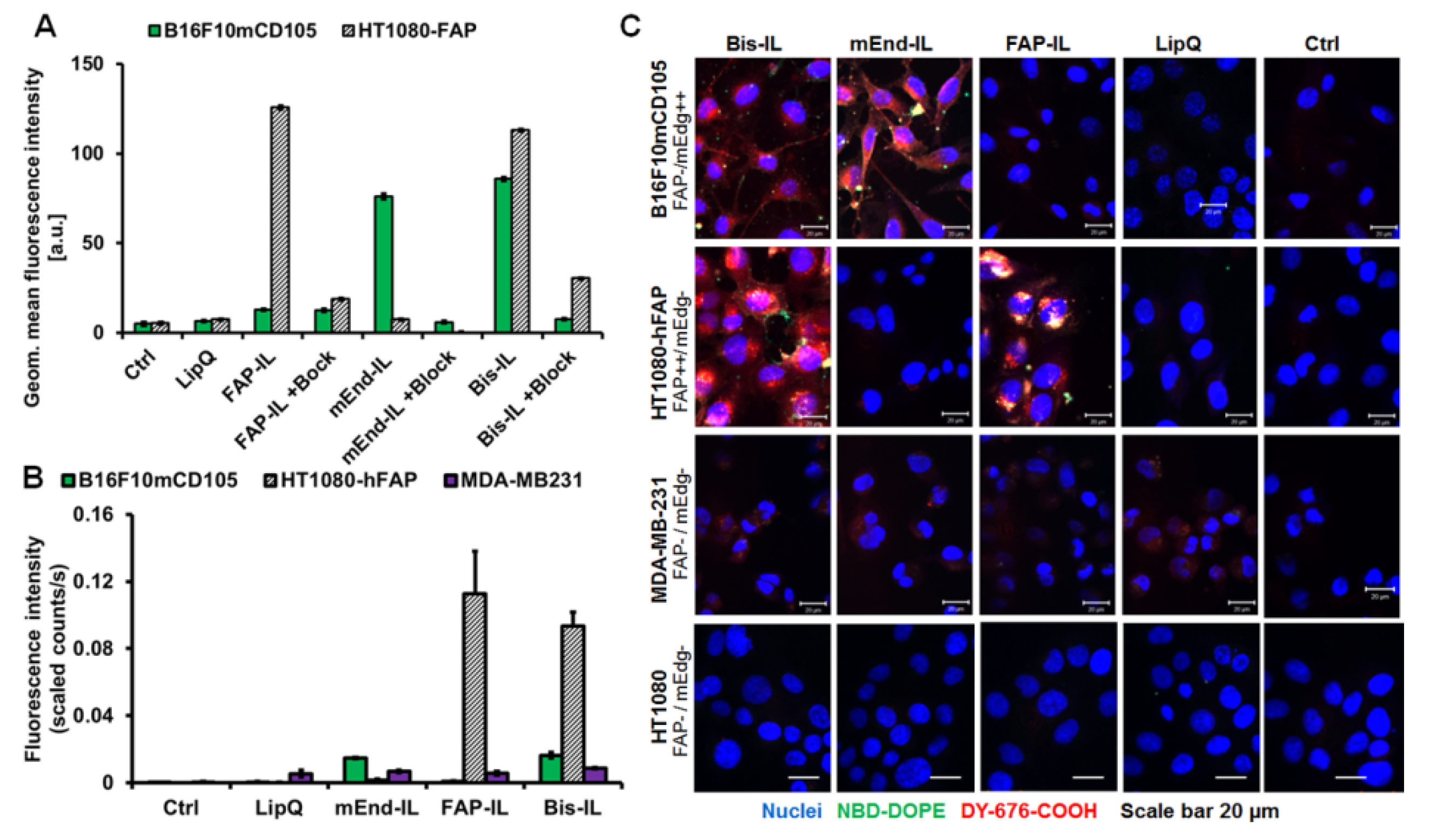
2.6. Validation of the Liposomal Binding and Uptake Selectivity in Cultured Target Cells
2.7. Animals and Implantation
2.8. Determination of Tumor Volumes
2.9. Whole Body NIRF Imaging of Mice, Excised Organs and Determination of Fluorescence Intensities
2.10. Euthanasia and Ex Vivo Determination of the Biodistribution of Liposomes
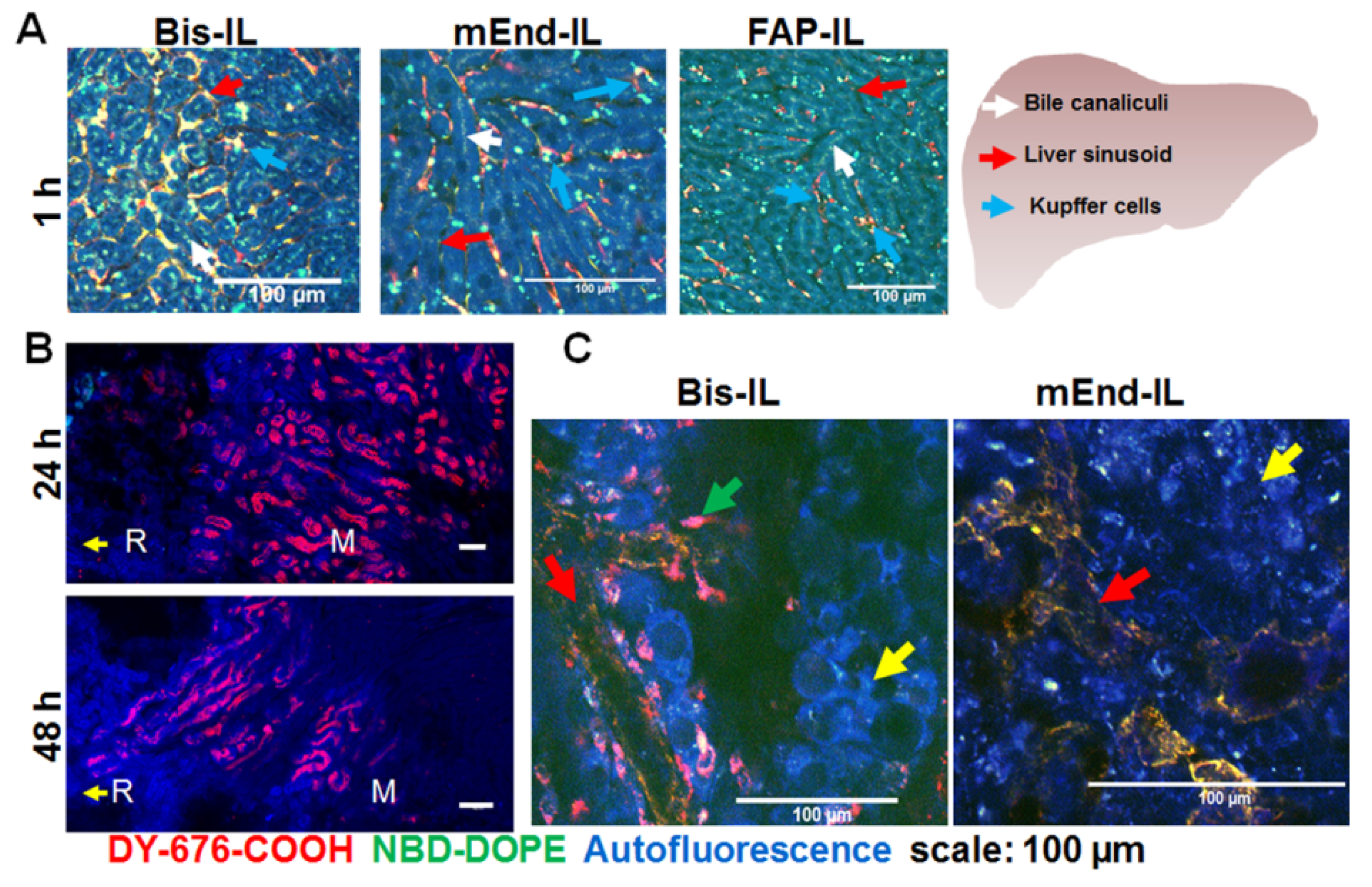
2.11. Verification of the Localization of Liposome Signals in Freshly Isolated Tissues
2.12. Statistical Evaluation of Significance
3. Results
3.1. Preparation and Physicochemical Characterization of Activatable Liposomes
3.2. The Targeted and Control Liposomes Show Dye Activatability upon Damage
3.3. Activatable Targeted Liposomes Are Selective for the Respective Target Cells In Vitro
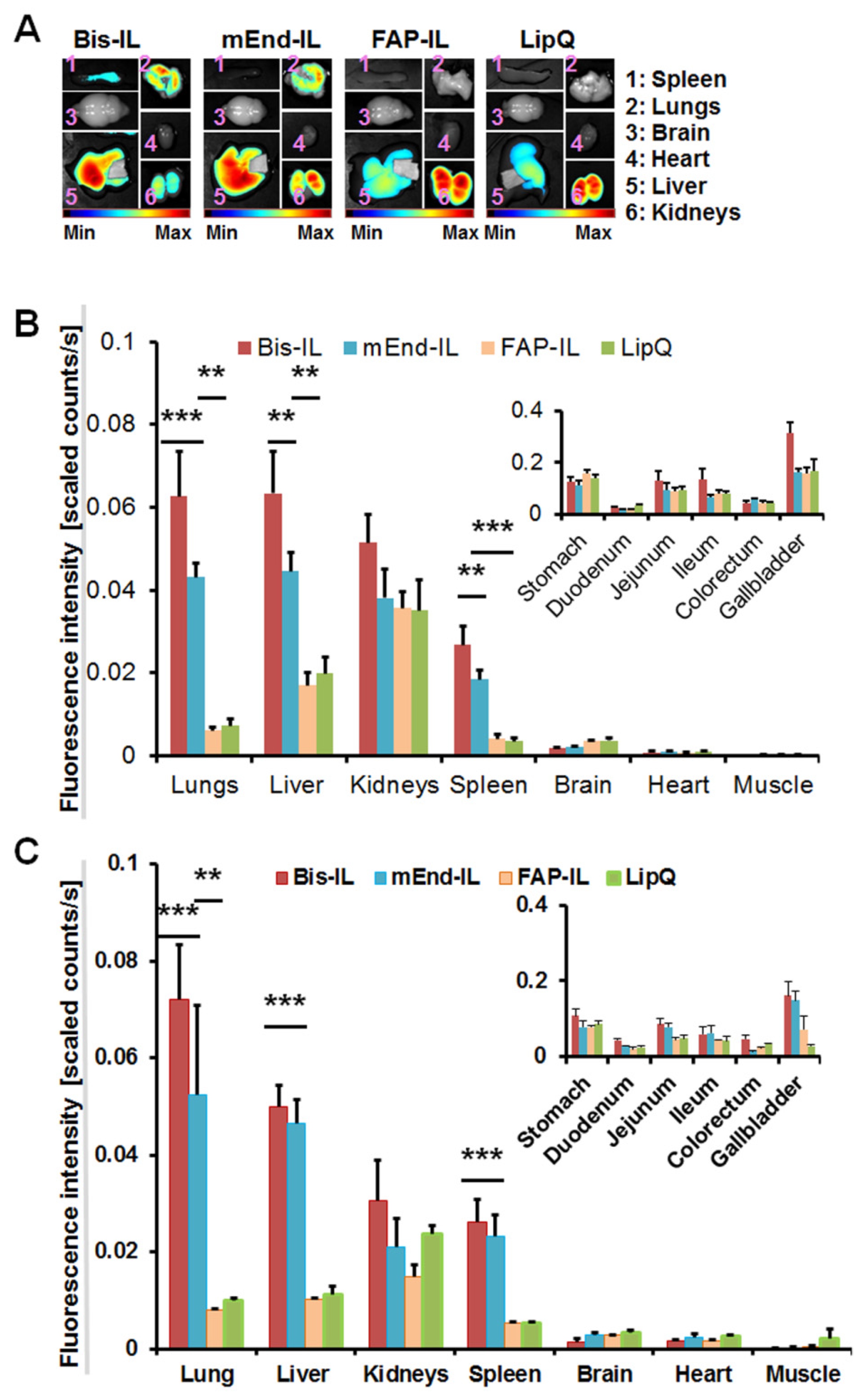
3.4. The Activatable Targeted Liposomes Show Distinct Biodistribution in Immune Deficient and Immune Competent Mice
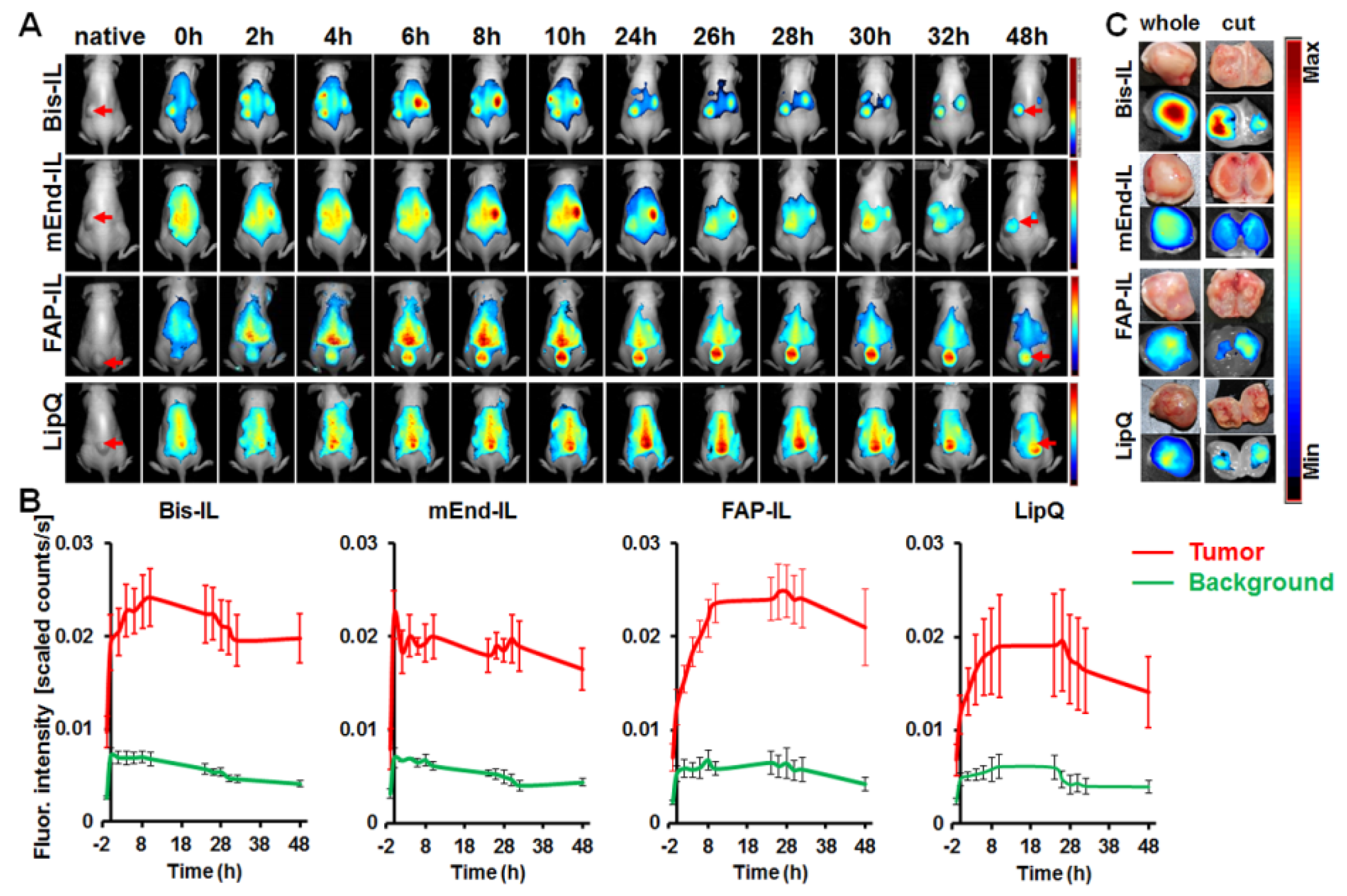
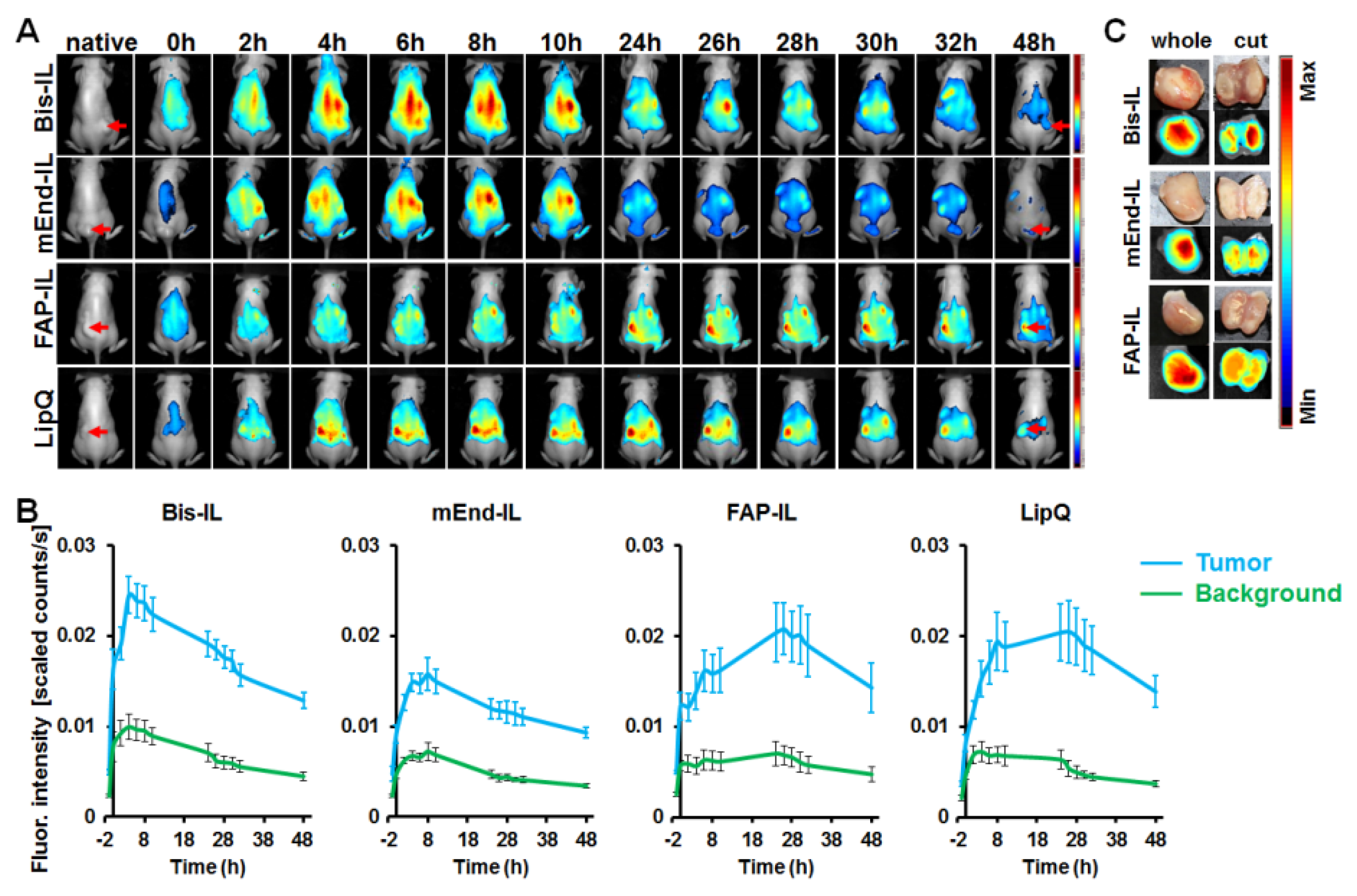
3.5. The Activatable Targeted Liposomes Show Distinct Accumulation in Tumor Models In Vivo
3.5.1. Liposome-Based Imaging of High FAP Expressing Tumor Models
3.5.2. Liposome-Based Imaging of FAP/Endoglin Negative Tumor Models
3.6. Accumulation of Bispecific Liposomes in Tumors Is Based on Target Binding, Whereas Accumulation in Mice Liver and Kidneys Is Based on Both Target Binding and Elimination
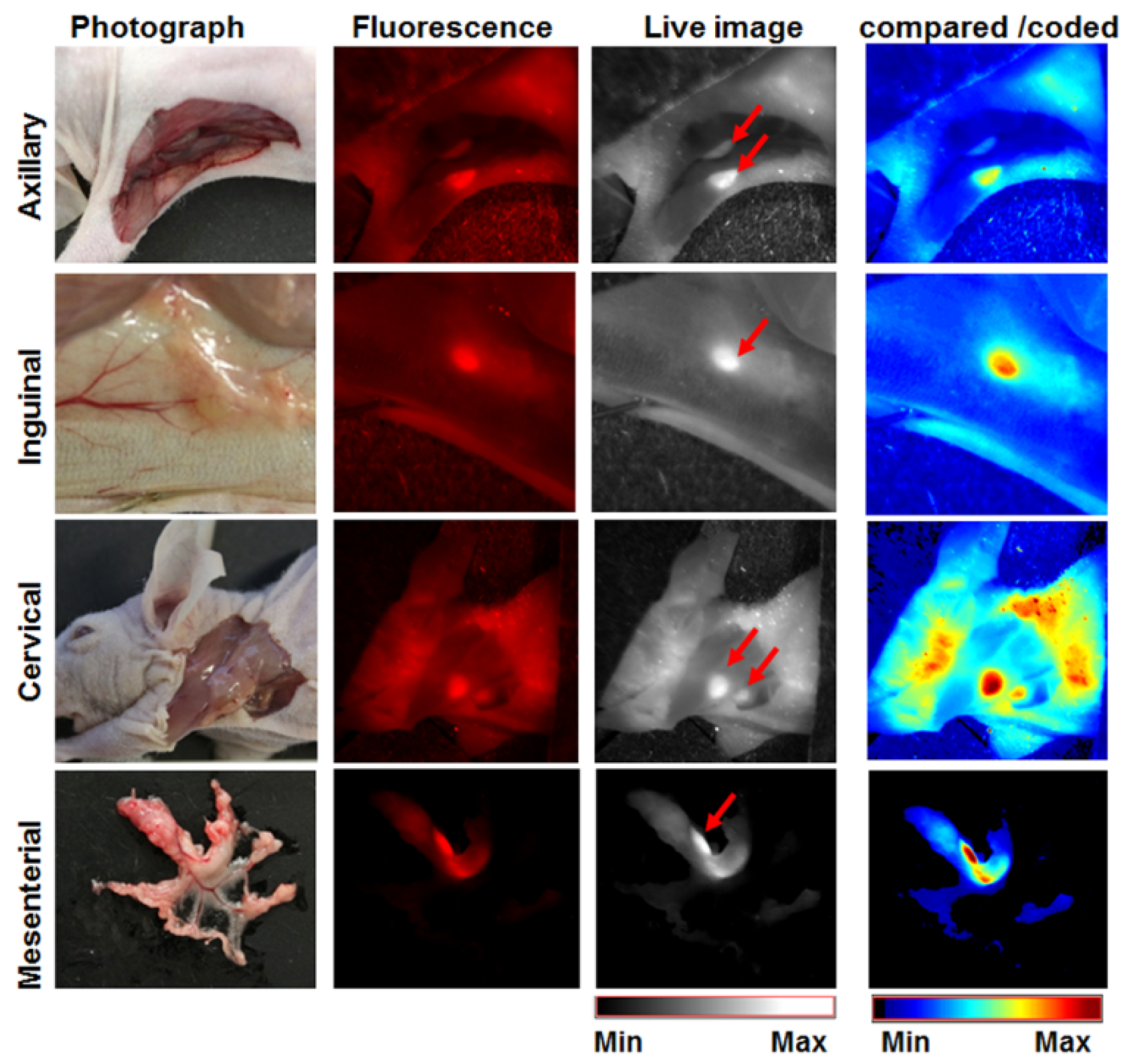
3.7. The Activatable Bispecific Liposome Is Suitable for Intraoperative Detection
4. Discussion
4.1. Physical Properties of the Liposomal Probes
4.2. Selectivity of Bispecific Liposomes to the Targets on Cultured Cells
4.3. Biodistribution, Selectivity and Potential Applications of Bispecific Liposomes In Vivo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fahr, A.; Liu, X. Drug delivery strategies for poorly water-soluble drugs. Expert Opin. Drug Deliv. 2007, 4, 403–416. [Google Scholar] [CrossRef]
- Aryasomayajula, B.; Salzano, G.; Torchilin, V.P. Multifunctional Liposomes. Methods Mol. Biol. 2017, 1530, 41–61. [Google Scholar] [CrossRef]
- Fahr, A.; van Hoogevest, P.; May, S.; Bergstrand, N.; ML, S.L. Transfer of lipophilic drugs between liposomal membranes and biological interfaces: Consequences for drug delivery. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2005, 26, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Gabizon, A.; Shmeeda, H.; Grenader, T. Pharmacological basis of pegylated liposomal doxorubicin: Impact on cancer therapy. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2012, 45, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Rüger, R.; Tansi, F.L.; Rabenhold, M.; Steiniger, F.; Kontermann, R.E.; Fahr, A.; Hilger, I. In vivo near-infrared fluorescence imaging of FAP-expressing tumors with activatable FAP-targeted, single-chain Fv-immunoliposomes. J. Control. Release Off. J. Control. Release Soc. 2014, 186C, 1–10. [Google Scholar] [CrossRef]
- Khorana, A.A.; Ryan, C.K.; Cox, C.; Eberly, S.; Sahasrabudhe, D.M. Vascular endothelial growth factor, CD68, and epidermal growth factor receptor expression and survival in patients with Stage II and Stage III colon carcinoma: A role for the host response in prognosis. Cancer 2003, 97, 960–968. [Google Scholar] [CrossRef] [Green Version]
- Funada, Y.; Noguchi, T.; Kikuchi, R.; Takeno, S.; Uchida, Y.; Gabbert, H.E. Prognostic significance of CD8+ T cell and macrophage peritumoral infiltration in colorectal cancer. Oncol. Rep. 2003, 10, 309–313. [Google Scholar] [CrossRef]
- Zhou, Q.; Peng, R.Q.; Wu, X.J.; Xia, Q.; Hou, J.H.; Ding, Y.; Zhou, Q.M.; Zhang, X.; Pang, Z.Z.; Wan, D.S.; et al. The density of macrophages in the invasive front is inversely correlated to liver metastasis in colon cancer. J. Transl. Med. 2010, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M. Cancer nanotechnology: Opportunities and challenges. Nat. Rev. Cancer 2005, 5, 161–171. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.P.; Schier, R.; McCall, A.M.; Simmons, H.H.; Horak, E.M.; Alpaugh, R.K.; Marks, J.D.; Weiner, L.M. High affinity restricts the localization and tumor penetration of single-chain fv antibody molecules. Cancer Res. 2001, 61, 4750–4755. [Google Scholar] [PubMed]
- Li, J.; Chen, K.; Liu, H.; Cheng, K.; Yang, M.; Zhang, J.; Cheng, J.D.; Zhang, Y.; Cheng, Z. Activatable near-infrared fluorescent probe for in vivo imaging of fibroblast activation protein-alpha. Bioconjugate Chem. 2012, 23, 1704–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Hong, K.; Kirpotin, D.B.; Colbern, G.; Shalaby, R.; Baselga, J.; Shao, Y.; Nielsen, U.B.; Marks, J.D.; Moore, D.; et al. Anti-HER2 immunoliposomes: Enhanced efficacy attributable to targeted delivery. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2002, 8, 1172–1181. [Google Scholar]
- Laginha, K.; Mumbengegwi, D.; Allen, T. Liposomes targeted via two different antibodies: Assay, B-cell binding and cytotoxicity. Biochim. Biophys. Acta 2005, 1711, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Bauer, S.; Jendro, M.C.; Wadle, A.; Kleber, S.; Stenner, F.; Dinser, R.; Reich, A.; Faccin, E.; Godde, S.; Dinges, H.; et al. Fibroblast activation protein is expressed by rheumatoid myofibroblast-like synoviocytes. Arthritis Res. Ther. 2006, 8, R171. [Google Scholar] [CrossRef] [Green Version]
- Brennen, W.N.; Isaacs, J.T.; Denmeade, S.R. Rationale behind targeting fibroblast activation protein-expressing carcinoma-associated fibroblasts as a novel chemotherapeutic strategy. Mol. Cancer Ther. 2012, 11, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T. Fibroblast activation protein-alpha and dipeptidyl peptidase IV (CD26): Cell-surface proteases that activate cell signaling and are potential targets for cancer therapy. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer Chemother. 2005, 8, 51–58. [Google Scholar] [CrossRef]
- Zhi, K.; Shen, X.; Zhang, H.; Bi, J. Cancer-associated fibroblasts are positively correlated with metastatic potential of human gastric cancers. J. Exp. Clin. Cancer Res. CR 2010, 29, 66. [Google Scholar] [CrossRef] [Green Version]
- Hofheinz, R.D.; al-Batran, S.E.; Hartmann, F.; Hartung, G.; Jager, D.; Renner, C.; Tanswell, P.; Kunz, U.; Amelsberg, A.; Kuthan, H.; et al. Stromal antigen targeting by a humanised monoclonal antibody: An early phase II trial of sibrotuzumab in patients with metastatic colorectal cancer. Onkologie 2003, 26, 44–48. [Google Scholar] [CrossRef]
- Jiang, G.M.; Xu, W.; Du, J.; Zhang, K.S.; Zhang, Q.G.; Wang, X.W.; Liu, Z.G.; Liu, S.Q.; Xie, W.Y.; Liu, H.F.; et al. The application of the fibroblast activation protein alpha-targeted immunotherapy strategy. Oncotarget 2016, 7, 33472–33482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Simms, A.E.; Mazur, A.; Wang, S.; Leon, N.R.; Jones, B.; Aziz, N.; Kelly, T. Fibroblast activation protein-alpha promotes tumor growth and invasion of breast cancer cells through non-enzymatic functions. Clin. Exp. Metastasis 2011, 28, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.O.; Mullins, S.R.; Franco-Barraza, J.; Valianou, M.; Cukierman, E.; Cheng, J.D. FAP-overexpressing fibroblasts produce an extracellular matrix that enhances invasive velocity and directionality of pancreatic cancer cells. BMC Cancer 2011, 11, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein-Goldberg, A.; Maman, S.; Witz, I.P. The role played by the microenvironment in site-specific metastasis. Cancer Lett. 2013. [Google Scholar] [CrossRef]
- Santos, A.M.; Jung, J.; Aziz, N.; Kissil, J.L.; Pure, E. Targeting fibroblast activation protein inhibits tumor stromagenesis and growth in mice. J. Clin. Investig. 2009, 119, 3613–3625. [Google Scholar] [CrossRef] [PubMed]
- Ge, A.Z.; Butcher, E.C. Cloning and expression of a cDNA encoding mouse endoglin, an endothelial cell TGF-beta ligand. Gene 1994, 138, 201–206. [Google Scholar] [CrossRef]
- Bernabeu, C.; Lopez-Novoa, J.M.; Quintanilla, M. The emerging role of TGF-beta superfamily coreceptors in cancer. Biochim. Biophys. Acta 2009, 1792, 954–973. [Google Scholar] [CrossRef]
- Aird, W.C. Molecular heterogeneity of tumor endothelium. Cell Tissue Res. 2009, 335, 271–281. [Google Scholar] [CrossRef]
- Gregory, A.L.; Xu, G.; Sotov, V.; Letarte, M. Review: The enigmatic role of endoglin in the placenta. Placenta 2014, 35, S93–S99. [Google Scholar] [CrossRef]
- Fonsatti, E.; Altomonte, M.; Arslan, P.; Maio, M. Endoglin (CD105): A target for anti-angiogenetic cancer therapy. Curr. Drug Targets 2003, 4, 291–296. [Google Scholar] [CrossRef]
- Fonsatti, E.; Jekunen, A.P.; Kairemo, K.J.; Coral, S.; Snellman, M.; Nicotra, M.R.; Natali, P.G.; Altomonte, M.; Maio, M. Endoglin is a suitable target for efficient imaging of solid tumors: In vivo evidence in a canine mammary carcinoma model. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 2037–2043. [Google Scholar]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A marker of tumor vasculature and potential target for therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansi, F.L.; Rüger, R.; Rabenhold, M.; Steiniger, F.; Fahr, A.; Kaiser, W.A.; Hilger, I. Liposomal Encapsulation of a Near-Infrared Fluorophore Enhances Fluorescence Quenching and Reliable Whole Body Optical Imaging Upon Activation In Vivo. Small 2013. [Google Scholar] [CrossRef]
- Tansi, F.L.; Rüger, R.; Rabenhold, M.; Steiniger, F.; Fahr, A.; Hilger, I. Fluorescence-quenching of a liposomal-encapsulated near-infrared fluorophore as a tool for in vivo optical imaging. J. Vis. Exp. Jove 2015, e52136. [Google Scholar] [CrossRef] [Green Version]
- Tansi, F.L.; Rüger, R.; Böhm, C.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. Potential of activatable FAP-targeting immunoliposomes in intraoperative imaging of spontaneous metastases. Biomaterials 2016, 88, 70–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocks, B.; Garin-Chesa, P.; Behrle, E.; Park, J.E.; Rettig, W.J.; Pfizenmaier, K.; Moosmayer, D. Species-crossreactive scFv against the tumor stroma marker “fibroblast activation protein” selected by phage display from an immunized FAP-/- knock-out mouse. Mol. Med. 2001, 7, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Matsuno, F.; Haruta, Y.; Kondo, M.; Tsai, H.; Barcos, M.; Seon, B.K. Induction of lasting complete regression of preformed distinct solid tumors by targeting the tumor vasculature using two new anti-endoglin monoclonal antibodies. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1999, 5, 371–382. [Google Scholar]
- Rüger, R.; Müller, D.; Fahr, A.; Kontermann, R.E. Generation of immunoliposomes using recombinant single-chain Fv fragments bound to Ni-NTA-liposomes. J. Drug Target. 2005, 13, 399–406. [Google Scholar] [CrossRef]
- Müller, D.; Trunk, G.; Sichelstiel, A.; Zettlitz, K.A.; Quintanilla, M.; Kontermann, R.E. Murine endoglin-specific single-chain Fv fragments for the analysis of vascular targeting strategies in mice. J. Immunol. Methods 2008, 339, 90–98. [Google Scholar] [CrossRef]
- Müller, D.; Karle, A.; Meissburger, B.; Höfig, I.; Stork, R.; Kontermann, R.E. Improved pharmacokinetics of recombinant bispecific antibody molecules by fusion to human serum albumin. J. Biol. Chem. 2007, 282, 12650–12660. [Google Scholar] [CrossRef] [Green Version]
- Iden, D.L.; Allen, T.M. In vitro and in vivo comparison of immunoliposomes made by conventional coupling techniques with those made by a new post-insertion approach. Biochim. Biophys. Acta 2001, 1513, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Allen, T.M.; Sapra, P.; Moase, E. Use of the post-insertion method for the formation of ligand-coupled liposomes. Cell. Mol. Biol. Lett. 2002, 7, 889–894. [Google Scholar]
- Feldman, J.P.; Goldwasser, R.; Mark, S.; Schwartz, J.; Orion, I. A Mathematical Model for Tumor Volume Evaluation using Two-Dimensions. J. Appl. Quant. Methods 2009, 4, 8. [Google Scholar]
- Tansi, F.L.; Rüger, R.; Kollmeier, A.M.; Böhm, C.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. A fast and effective determination of the biodistribution and subcellular localization of fluorescent immunoliposomes in freshly excised animal organs. BMC Biotechnol. 2017, 17, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansi, F.L.; Rüger, R.; Böhm, C.; Steiniger, F.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. Activatable bispecific liposomes bearing fibroblast activation protein directed single chain fragment/Trastuzumab deliver encapsulated cargo into the nuclei of tumor cells and the tumor microenvironment simultaneously. Acta Biomater. 2017, 54, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Tansi, F.L.; Ruger, R.; Kollmeier, A.M.; Rabenhold, M.; Steiniger, F.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. Endoglin based in vivo near-infrared fluorescence imaging of tumor models in mice using activatable liposomes. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1389–1400. [Google Scholar] [CrossRef]
- Tansi, F.L.; Ruger, R.; Kollmeier, A.M.; Rabenhold, M.; Steiniger, F.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. Dataset on the role of endoglin expression on melanin production in murine melanoma and on the influence of melanin on optical imaging. Data Brief 2018, 20, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, S.J.; Turner, C.; Heath, A.; Feavers, I.; Vatn, I.; Natvig, J.B.; Thompson, K.M. Clonal analysis of a human antimouse antibody (HAMA) response. Scand. J. Immunol. 2003, 57, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, M.; Kosaka, N.; Choyke, P.L.; Kobayashi, H. H-type dimer formation of fluorophores: A mechanism for activatable, in vivo optical molecular imaging. ACS Chem. Biol. 2009, 4, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.K.; Cook, R.M. Intramolecular dimers: A new design strategy for fluorescence-quenched probes. Chemistry 2003, 9, 3466–3471. [Google Scholar] [CrossRef]
- Wu, P.; Brand, L. Resonance energy transfer: Methods and applications. Anal. Biochem. 1994, 218, 1–13. [Google Scholar] [CrossRef]
- Weissleder, R.; Ntziachristos, V. Shedding light onto live molecular targets. Nat. Med. 2003, 9, 123–128. [Google Scholar] [CrossRef]
- Zonios, G.; Dimou, A. Melanin optical properties provide evidence for chemical and structural disorder in vivo. Opt. Express 2008, 16, 8263–8268. [Google Scholar] [CrossRef]
- Ito, A.S.; Azzellini, G.C.; Silva, S.C.; Serra, O.; Szabo, A.G. Optical absorption and fluorescence spectroscopy studies of ground state melanin-cationic porphyrins complexes. Biophys. Chem. 1992, 45, 79–89. [Google Scholar] [CrossRef]
- Saw, P.E.; Park, J.; Lee, E.; Ahn, S.; Lee, J.; Kim, H.; Kim, J.; Choi, M.; Farokhzad, O.C.; Jon, S. Effect of PEG pairing on the efficiency of cancer-targeting liposomes. Theranostics 2015, 5, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.E.; Chen, Z.G.; Shin, D.M. Nanoparticle therapeutics: An emerging treatment modality for cancer. Nat. Rev. Drug Discov. 2008, 7, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Paauwe, M.; Schoonderwoerd, M.J.A.; Helderman, R.; Harryvan, T.J.; Groenewoud, A.; van Pelt, G.W.; Bor, R.; Hemmer, D.M.; Versteeg, H.H.; Snaar-Jagalska, B.E.; et al. Endoglin Expression on Cancer-Associated Fibroblasts Regulates Invasion and Stimulates Colorectal Cancer Metastasis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 6331–6344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mufamadi, M.S.; Pillay, V.; Choonara, Y.E.; Du Toit, L.C.; Modi, G.; Naidoo, D.; Ndesendo, V.M. A review on composite liposomal technologies for specialized drug delivery. J. Drug Deliv. 2011, 2011, 939851. [Google Scholar] [CrossRef]
- Balza, E.; Castellani, P.; Zijlstra, A.; Neri, D.; Zardi, L.; Siri, A. Lack of specificity of endoglin expression for tumor blood vessels. Int. J. Cancer. J. Int. Du Cancer 2001, 94, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Rabenhold, M.; Steiniger, F.; Fahr, A.; Kontermann, R.E.; Ruger, R. Bispecific single-chain diabody-immunoliposomes targeting endoglin (CD105) and fibroblast activation protein (FAP) simultaneously. J. Control. Release Off. J. Control. Release Soc. 2015, 201, 56–67. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Z-Average [nm] | PDI | Zeta Potential [mV] | DY-676 (µg/mL) |
|---|---|---|---|---|
| LipQ * | 128.1 ± 2 | 0.072 ± 0.02 | −10.9 ± 0.7 | 161 |
| LipQ | 133.0 ± 1 | 0.081 ± 0.10 | −17.0 ± 1.1 | 152 |
| mEnd-IL | 137.0 ± 2 | 0.180 ± 0.01 | −13.0 ± 0.4 | 158 |
| FAP-IL | 139.1 ± 2.5 | 0.079 ± 0.02 | −15.4 ± 4.9 | 148 |
| Bi-FAP/mEnd-IL | 154.5 ± 5.3 | 0.107 ± 0.02 | −20.5 ± 0.64 | 178 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tansi, F.L.; Rüger, R.; Kollmeier, A.M.; Rabenhold, M.; Steiniger, F.; Kontermann, R.E.; Teichgräber, U.K.; Fahr, A.; Hilger, I. Targeting the Tumor Microenvironment with Fluorescence-Activatable Bispecific Endoglin/Fibroblast Activation Protein Targeting Liposomes. Pharmaceutics 2020, 12, 370. https://doi.org/10.3390/pharmaceutics12040370
Tansi FL, Rüger R, Kollmeier AM, Rabenhold M, Steiniger F, Kontermann RE, Teichgräber UK, Fahr A, Hilger I. Targeting the Tumor Microenvironment with Fluorescence-Activatable Bispecific Endoglin/Fibroblast Activation Protein Targeting Liposomes. Pharmaceutics. 2020; 12(4):370. https://doi.org/10.3390/pharmaceutics12040370
Chicago/Turabian StyleTansi, Felista L., Ronny Rüger, Ansgar M. Kollmeier, Markus Rabenhold, Frank Steiniger, Roland E. Kontermann, Ulf K. Teichgräber, Alfred Fahr, and Ingrid Hilger. 2020. "Targeting the Tumor Microenvironment with Fluorescence-Activatable Bispecific Endoglin/Fibroblast Activation Protein Targeting Liposomes" Pharmaceutics 12, no. 4: 370. https://doi.org/10.3390/pharmaceutics12040370