1. Introduction
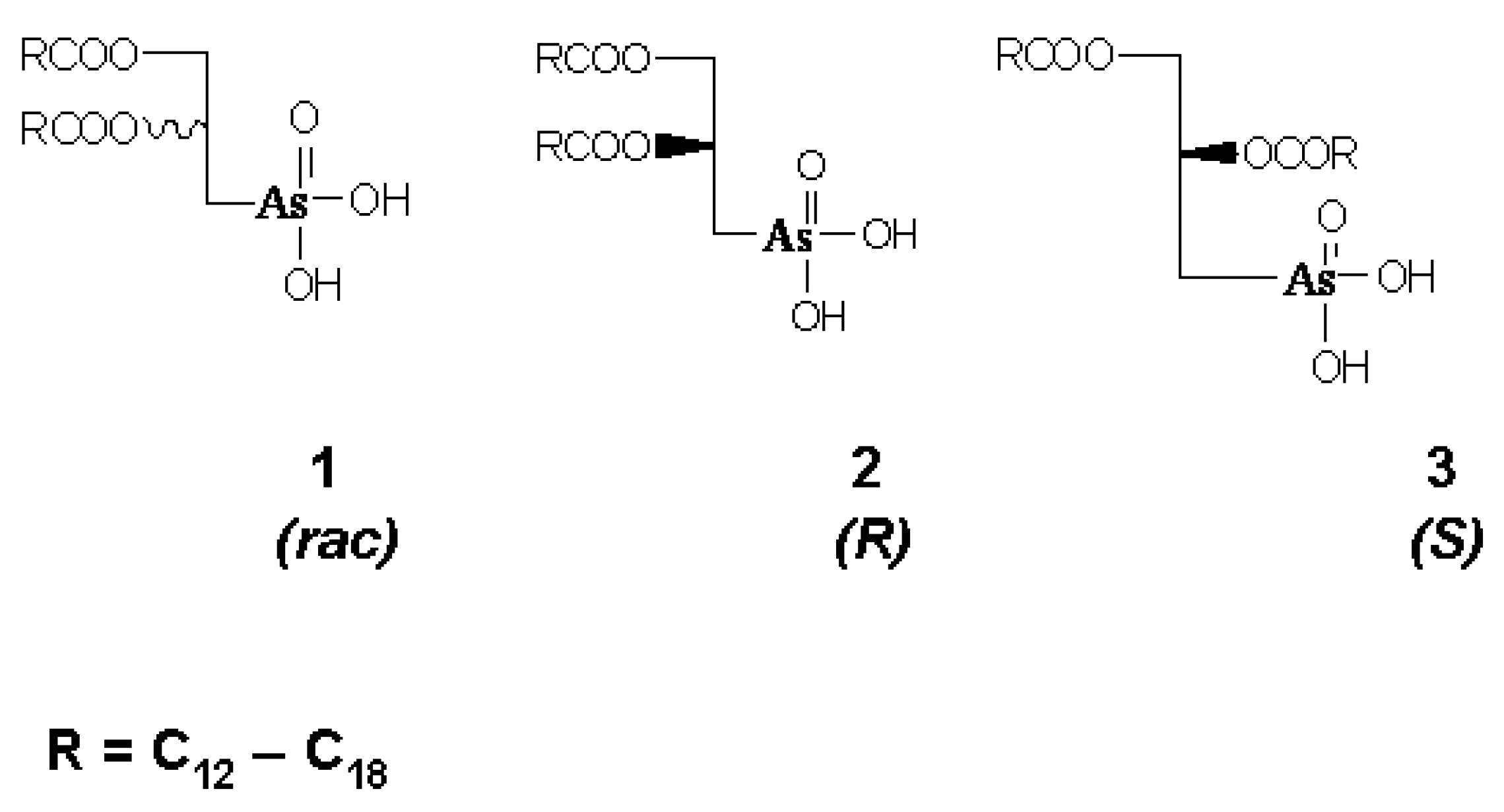
Arsonolipids are analogues of phosphonolipids in which P has been replaced by as in the lipid head group. (
Scheme 1) [
1,
2]. Arsonoliposomes (ARSL) constitute a particular class of liposomes, which are consisted of mixtures of arsonolipids, phospholipids, and cholesterol [
2,
3]. Nanosized ARSL demonstrate high toxicity against specific cancer cells, while being substantially less toxic towards normal cells, as proven by in vitro [
4,
5,
6] and in vivo studies [
7].
Specific ARSL compositions that include a polyethylene-glycol (PEG) coating on the lipid vesicle surface were demonstrated to have very high integrity during incubation in presence of serum proteins (compared to other ARSL compositions) [
8,
9], making them suitable for in vivo administration as cancer therapeutics [
10,
11]. In more detail, the in vitro integrity of ARSL composed of arsonolipid (C16), DSPC, and cholesterol (at 8:12:10 mole ratio) during incubation in serum proteins was highly increased when 8 mole% of DSPE-conjugated PEG was added in their membrane [
9].
The in vitro integrity tests for ARSL formulations was proven to be a good predictive tool for the in vivo bioavailability of arsenic; ARSL types that demonstrated higher integrity were also found to have significantly improved pharmacokinetics (compare to the ARSL types with lower in vitro integrity) [
10,
11]. In more detail, the levels of arsenic measured in abdominal tissues and in blood were significantly higher after administration of the more stable (with higher in vitro integrity) DSPC-containing ARSL [
11], compared to the less stable PC-containing ARSL [
10]. Furthermore, when the ARSL were coated with PEG molecules (in the above mentioned lipid composition, which conferred highly stable vesicles [
9]), the pharmacokinetic profile of arsenic was influenced even more, resulting in higher blood-circulation times, compared to those obtained by other (less stable) ARSL formulations [
10,
11].
Nevertheless, arsonoliposomes have some drawbacks, which if encountered successfully may increase their potential applicability. The two most important drawbacks are their low toxicity towards some cancer types, and their—up-to-date—inability to overcome the BBB. Indeed, while being highly toxic towards some cancer cell types, such as human leukemia cells (NB4 and HL60), prostatic cancer PC3 cells, and rat brain glioma C6 cells, ARSL did not exhibit the same high toxicity for other cancer cells, such as human breast adenocarcinoma MDA-MB-468 cells, and rat pituitary tumor GH3 cells [
5,
6]. Additionally, arsenic was not detected in the brain of animals injected with ARSL, in all the cases of ARSL which were studied in vivo [
10,
11], thus posing an interesting challenge for further exploitation. Indeed, if ARSL could overcome the blood brain barrier, they could potentially kill parasites in the central nervous system or act as anticancer therapeutics against difficult or even impossible to treat brain tumors.
Based on the identified most stable ARSL lipid composition, we attempted herein to investigate the potential to develop novel ARSL formulations that incorporate a curcumin lipidic derivative (TREG), as a method to increase their distribution in the brain; and/or encapsulate Doxorubicin (DOX), as a method to increase their toxicity towards cancer cells.
DOX is a highly potent anticancer drug; however, its serious side-effects, and especially its cardiotoxicity, restricts its use in many situations [
12]. The latter problem has been resolved by the construction of PEGylated DOX-loaded liposomes, which realize altered DOX pharmacokinetics and consequently, reduced toxicity [
12,
13,
14]. The easy loading of high amounts of DOX in pre-formed empty PEG-liposomes by an active-loading protocol [
15,
16] has highly contributed to the development of the long-circulating liposomal formulation of DOX, which is in clinical use for more than 25 years.
Curcumin (CUR) is a polyphenol that demonstrates interesting anti-inflammatory activity, while its potential anticancer activity is a controversial issue [
17]. A lipid derivative of curcumin, TREG, was previously synthesized and incorporated in liposomes, as an approach for development of nanoparticulate therapeutics for Alzheimer’s disease (AD) [
18]. In addition to their high affinity for amyloid species [
18,
19], TREG-liposomes demonstrated increased brain targeting potential, while the integrity of TREG-incorporating liposomes during incubation in presence of serum proteins was increased, as demonstrated in the case of non-targeted as well as brain-targeted liposomes (compared to control liposomes without TREG, in both cases) [
20,
21].
For the reasons mentioned above, we investigated herein the ability to incorporate TREG in ARSL, as well as the potential to load ARSL-types with DOX. Since synergistic anticancer activity between CUR and DOX has been reported before [
22,
23], perhaps the presence of TREG on DOX-loaded ARSL may confer increased anticancer activity in addition to any potential increase of the vesicle stability and/or targeting capability.
For the loading of DOX in both, ARSL and TREG-ARSL, the active loading procedure was applied [
15,
16]. Preliminary evaluation of the toxicity of some of the novel vesicle types towards cancer cell lines, and normal cells, was finally performed.
2. Materials and Methods
1,2-Distearoyl-sn-glycerol-3-phosphatidyl-choline [DSPC], and 1,2-Distearoyl-sn-glycerol-3-phosphatidyl-ethanolamine-
N-[methoxy(polyethylene-glycol)-2000] [PEG2000] were purchased from Lipoid, Germany. Cholesterol (Chol) was purchased from Sigma-Aldrich (Darmstadt, Germany). The rac-2,3-dipalmitoyl-oxypropylarsonic acid [ARS] (C
16), was synthesized as described in detail before [
1,
24,
25]; TREG lipid was also synthesized using the method reported in detail before [
18,
19]. Doxorubicin, hydrochloric salt (DOX) was purchased by Tocris Bioscience, UK. Fetal Calf Serum (FCS) was from Sigma (Darmstadt, Germany).
A bath sonicator (Branson) and a microtip-probe sonicator (Sonics and Materials, Leicestershire, UK) were used for liposome preparation. Protein concentrations were measured by Bradford microassay (1–10 µg protein/mL). The Cell Viability MTT assay was carried out with 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reagent, which was purchased by Sigma-Aldrich. All other reagents and chemicals used were of analytical grade, and were purchased from Sigma-Aldrich. The osmolarity of solutions used for liposome formation was measured by a Roebling osmometer and adjusted to 300 mOsm with NaCl (if needed).
2.1. Preparation of TREG-Incorporating ARSL (TREG-ARSL)
Liposomes (ARSL and TREG-incorporating arsonoliposomes (TREG-ARSL)) with the lipid compositions reported in
Table 1 were prepared as mentioned below. The specific ARSL composition used (DSPC/ARS/Chol (12:8:10 mol/mol containing 8 mole% PEG2000)) was the one that was previously identified to confer the highest integrity during incubation in presence of FCS (compared to other ARSL compositions) [
9]. Additionally, conventional liposomes (without arsonolipids) (LIP) and TREG-incorporating liposomes (TREG-LIP) were also formulated to be used as control formulations. For all liposome types, the thin-film-hydration method followed by probe sonication for size reduction was applied [
26]. In brief, lipids were dissolved in chloroform/methanol (2:1
v/
v), and appropriate volumes from the corresponding organic solutions, in order to confer the specific lipid compositions required (as presented in
Table 1), were added in a round bottomed flask; the total lipid concentration in the liposome dispersions was always equal to 20 mg/mL. The organic solvents were then evaporated by rotary evaporation (30 min; 105 rpm, 60 °C), and the lipids formed a thin lipid film on the sides of the flask. Any residual organic solvent was removed by subjection of the flask under a stream of N
2 for 15 min.
The resulting lipid film was subsequently hydrated at 60 °C with 1 mL of ammonium sulfate solution (120 mM, pH 5.5, 300 mOsm), if the vesicles were to be used for DOX loading, or with 1 mL calcein solution (100 mM, pH 7.40, 300 mOsm), if the vesicles were to be used for vesicle integrity studies, to form multilamellar vesicles (MLV). For physicochemical characterization, liposomes were hydrated with PBS buffer (pH 7.40, 300 mOsm). Vesicle size was then reduced by subjection to a high intensity (750 Watt) probe sonicator (Sonics and Materials, Leicestershire, UK) for (at least) two 15-min cycles, until the dispersion became completely clear, and then left at 60 °C for 1 h, to anneal any structural defects. Any traces of titanium, which may have leaked from the probe microtip, or any lipid aggregates present in the liposome dispersions, were removed by centrifugation at RCF 1600× g for 5 min (Scilogex 2012 microcentrifuge, Rocky Hill, CT, USA).
The exact lipid content of the resulting liposomes was measured by the Stewart assay, a colorimetric method used routinely for the quantification of phospholipids [
27]. Liposomes were purified from non-encapsulated solutes (calcein or DOX) by size exclusion chromatography (SEC), using a Sepharose 4B-CL column (40 × 1 cm), which was eluted with PBS buffer (pH 7.40), or by repeated ultracentrifugations for 1 h (each) at 60,000 rpm (Sorvall WX90 Ultra, Thermo Scientific, Waltham, MA, USA), depending on the need to re-concentrate the sample (or not) for the specific study that followed (if dilution occurring during SEC would cause a need for re-concentration, ultracentrifugation was preferred).
2.2. Physicochemical Properties of Liposomes
All the liposome types prepared were characterized for their lipid concentration, mean diameter, size distribution, and zeta-potential. For measurement of their size, the liposome dispersions were diluted to a final concentration of 0.4 mg/mL, and measured by dynamic light scattering (Malvern Instruments, Zetasizer Nano SZ, Malvern, UK), which enables the mass distribution of particle size to be obtained in the range between 0.3 nm–10 µm. Phosphate buffered saline (PBS 10 mM), pH 7.40 was used for dilution of LIP dispersions, after being filtered through polycarbonate filters (0.22 μm) (Millipore, UK). Particle size measurements were carried out with a fixed angle of 173° for backscatter correction, at 25 °C. The sizes reported correspond to the z-average means of the hydrodynamic diameters of the liposomes.
For ζ-potential values, the electrophoretic mobility of the liposome dispersions was measured at 25 °C, by the same instrument. Zeta potential values were obtained (by the instrument) from the electrophoretic mobility, according to the Smoluchowski equation.
The percent incorporation of TREG in liposomes (compared to the initial amount of TREG added in the samples during liposome preparation), was quantified as reported before [
18,
19], in order to verify if the complete amount of TREG was indeed incorporated in the liposomes, and thus exclude any potential of micelle of small lipid aggregate formation. In brief, HPLC analysis of a specific quantity of liposomes (lipid amount) was carried out, before as well as after purification of the liposome dispersions, both by ultracentrifugation and size exclusion chromatography (as described above); integration of the corresponding peaks of TREG followed (see
Supplementary data). For this, the liposome samples were dissolved in MeOH and HPLC was performed with a Lichrosphere 100 RP-18 (5 mm) column; eluted with MeOH (as mobile phase) at a 1 mL/min flow rate, by a Shimatzu, LC-20AB Prominence Liquid Chromatography System. TREG elution was monitored at 330 nm. Blank liposomes (with no TREG) did not give any reading at the time period that TREG eluted from the column.
The physical stability of ARSL and TREG-ARSL was monitored by measuring their mean hydrodynamic diameter, polydispersity index, and zeta potential (as mentioned above) at specific time periods during their incubation at 4 °C, for up to 56 days.
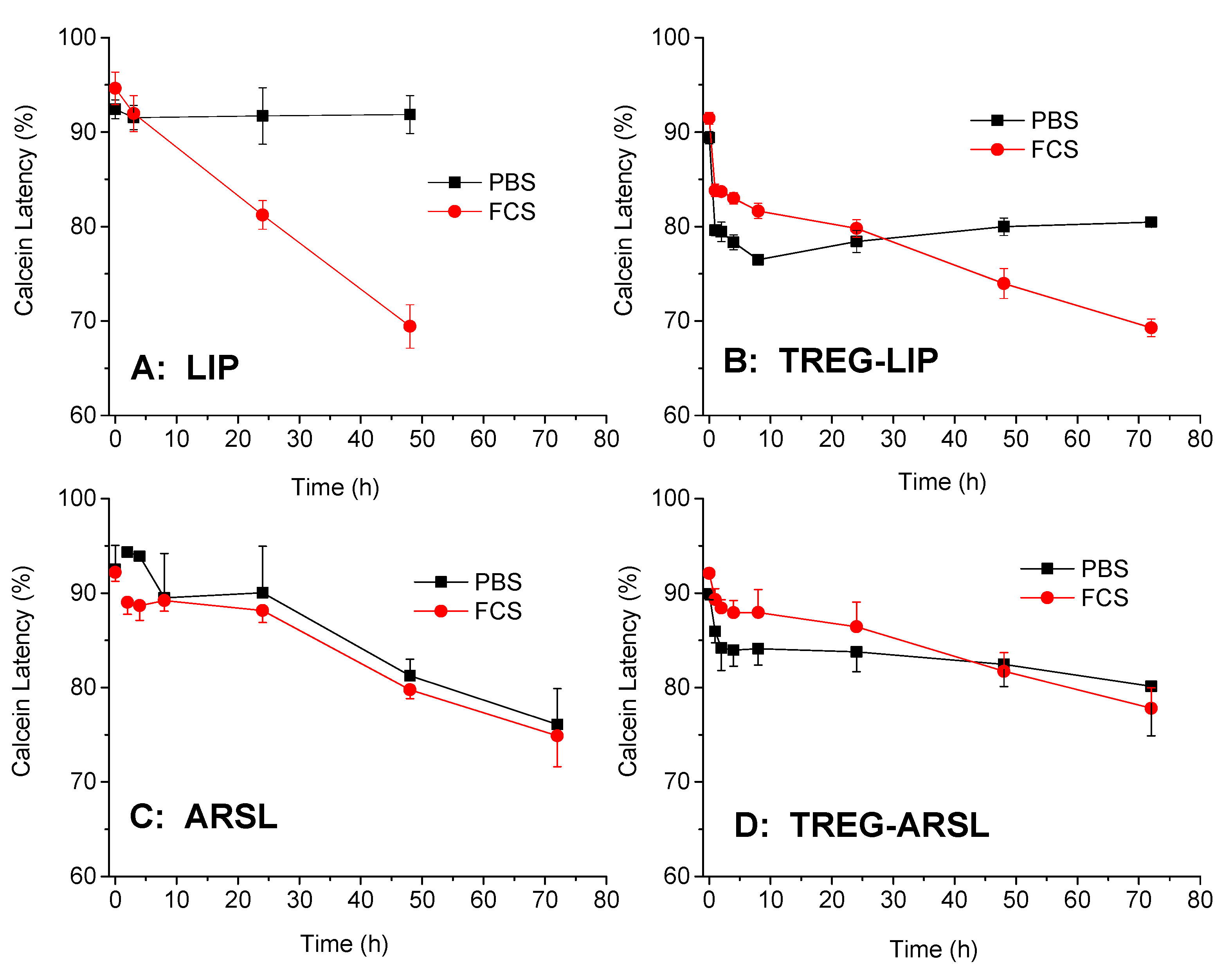
2.3. Liposome Integrity Studies (Calcein Latency)
Calcein latency was monitored during incubation of the various liposome types in buffer as well as in the presence of serum proteins (80% FCS) as a measure of the liposome integrity in blood [
28]. Calcein was encapsulated in the vesicles at a quenched concentration (100 mM) and for latency calculation, liposome samples (20 μL) were diluted with 4 mL buffer, pH 7.40, and the fluorescence intensity (FI) of the diluted samples was measured (EM 470 nm, EX 520 nm, 5 nm slits), before and after disruption of the liposomes by addition of Triton X-100 at a final concentration of 1%
v/
v (that ensures liposome disruption and release of all encapsulated dye). Percent latency (%
latency) values were then calculated according to Equation (1):
where,
FBT and
FAT are the calcein fluorescence intensities, before and after vesicle disruption by Triton X-100, respectively.
FAT was multiplied with 1.1 for correction due to the dilution by Triton.
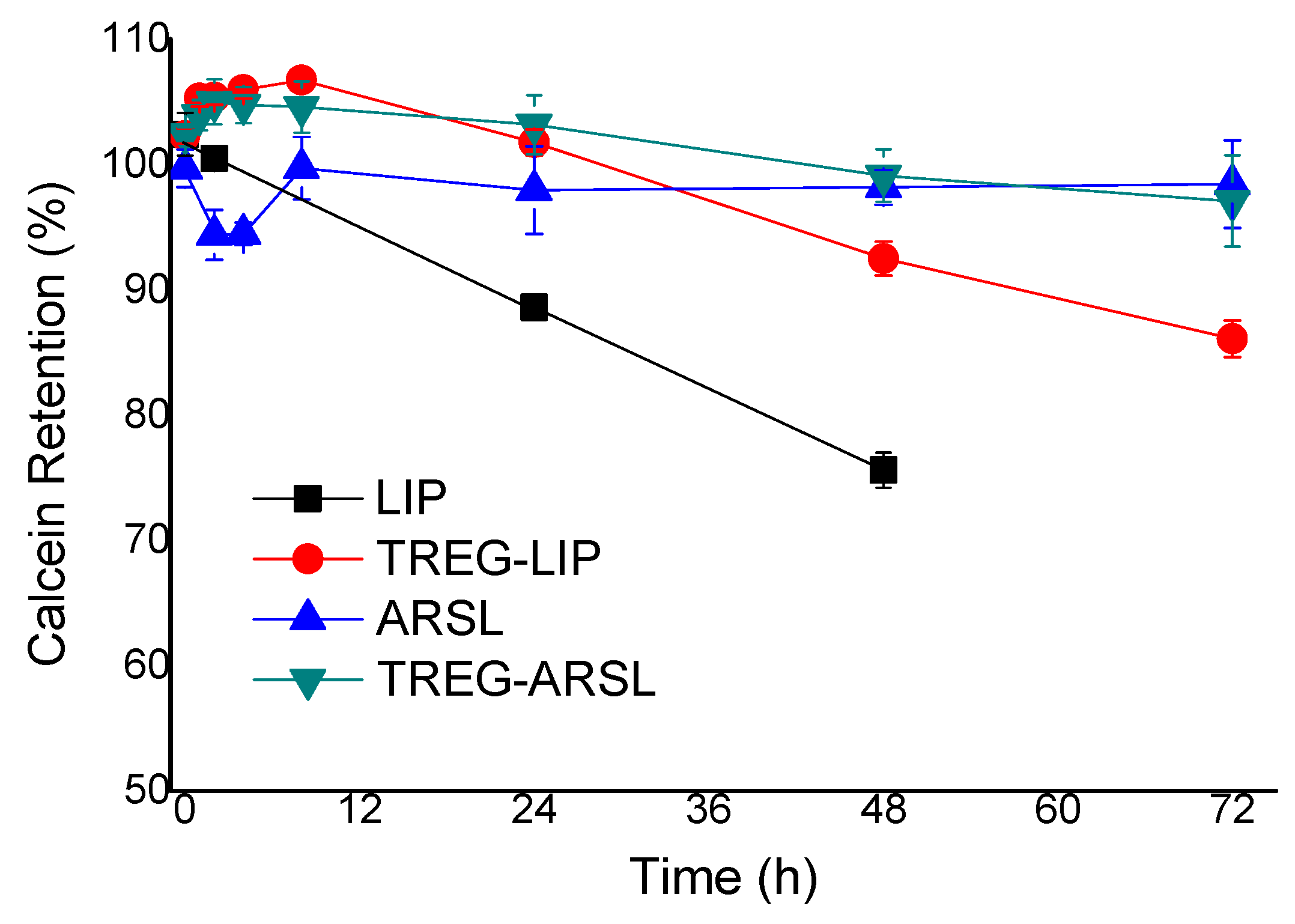
For evaluation of the specific disruptive effect that serum proteins cause towards liposome membranes, calcein
retention (%) of vesicles was calculated from the
latency of the liposomes during incubation in buffer, and the corresponding (at the same time point)
latency in FCS, according to Equation (2):
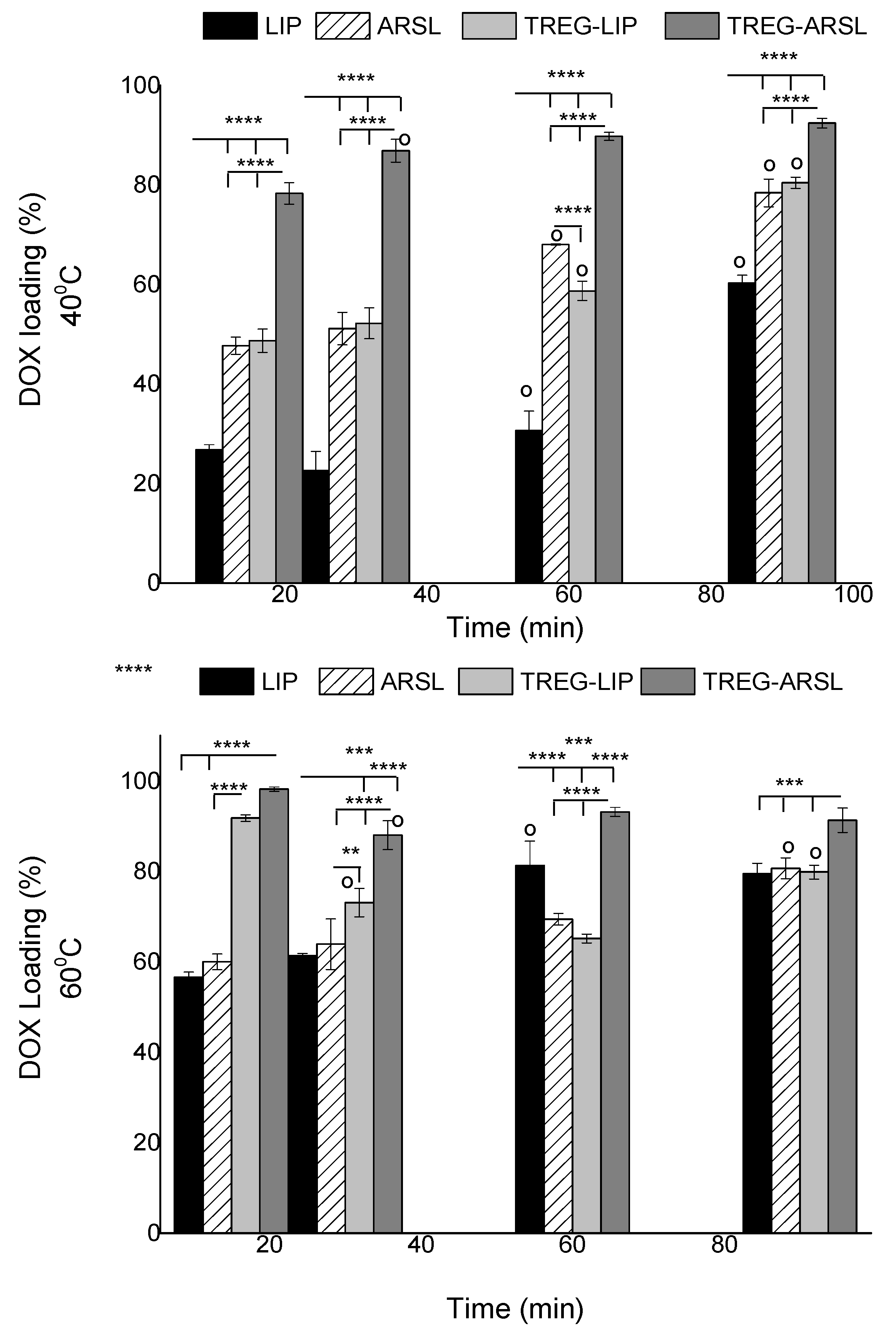
2.4. Loading/Release of DOX in/from Liposomes
DOX was loaded in LIP and TREG-LIP (control formulations) as well as in ARSL and TREG-ARSL by the active loading method, as described in detail elsewhere [
16]. In brief, empty vesicles were pre-formulated in ammonium sulfate ([NH
4]
2SO
4 (120 mM)), as described above. The vesicles were then ultra-centrifuged (Sorvall WX90 Ultra, Thermo Scientific) at 60,000 rpm for 1 h and re-suspended in PBS pH 7.4, for exchange of the dispersion media. Ultracentrifugation was used for external buffer exchange instead of size-exclusion chromatography, in order to avoid an extra step to re-concentrate the sample, since chromatography results in 3–5 times dilution of liposome dispersions. For the active loading of DOX, all types of (empty) liposomes, at a lipid concentration of 1.4 mg/mL (in PBS) were incubated in presence of a 0.2 mg/mL DOX solution (in PBS) (corresponding to a lipid/DOX ratio equal to 7:1 [
w/
w]), for various time periods (15, 30, 60 and 90 min) at 40 °C and 60 °C. After this, the liposomes were purified from non-encapsulated drug by two repeated ultracentrifugation steps (60,000 rpm for 1 h, each), in order to achieve high purification, without dilution of samples. Supernatants containing non-encapsulated DOX were collected, as well as the DOX-loaded liposome pellets, which were re-suspended in 1 mL of PBS, for measurement of DOX loading efficiency. For this, the liposome lipid concentration was measured by the Stewart assay, and the DOX concentration in free DOX solutions and liposomes was calculated by measuring the fluorescence intensity (FI, EX 485 nm, EM 590 nm, slits 5 nm) of DOX in every sample by a Shimadzu RF-Fluorescence Spectrophotometer. Liposomal samples were measured in the presence of Triton X-100 (1%) in order to dissolve the liposomes. To estimate the drug concentration, DOX calibration curves in the concentration range between 2.5 and 40 μg/mL were prepared, in media with similar composition as the samples (1% Triton X-100 in PBS, after ensuring that the presence of the dissolved lipids within the concentration range they were present in the samples, did not modify the calibration curve).
The final D/L ratio (
w/
w) was estimated and compared with the initial one, for calculation of the DOX encapsulation efficiency (%) of each liposome type, according to Equation (3):
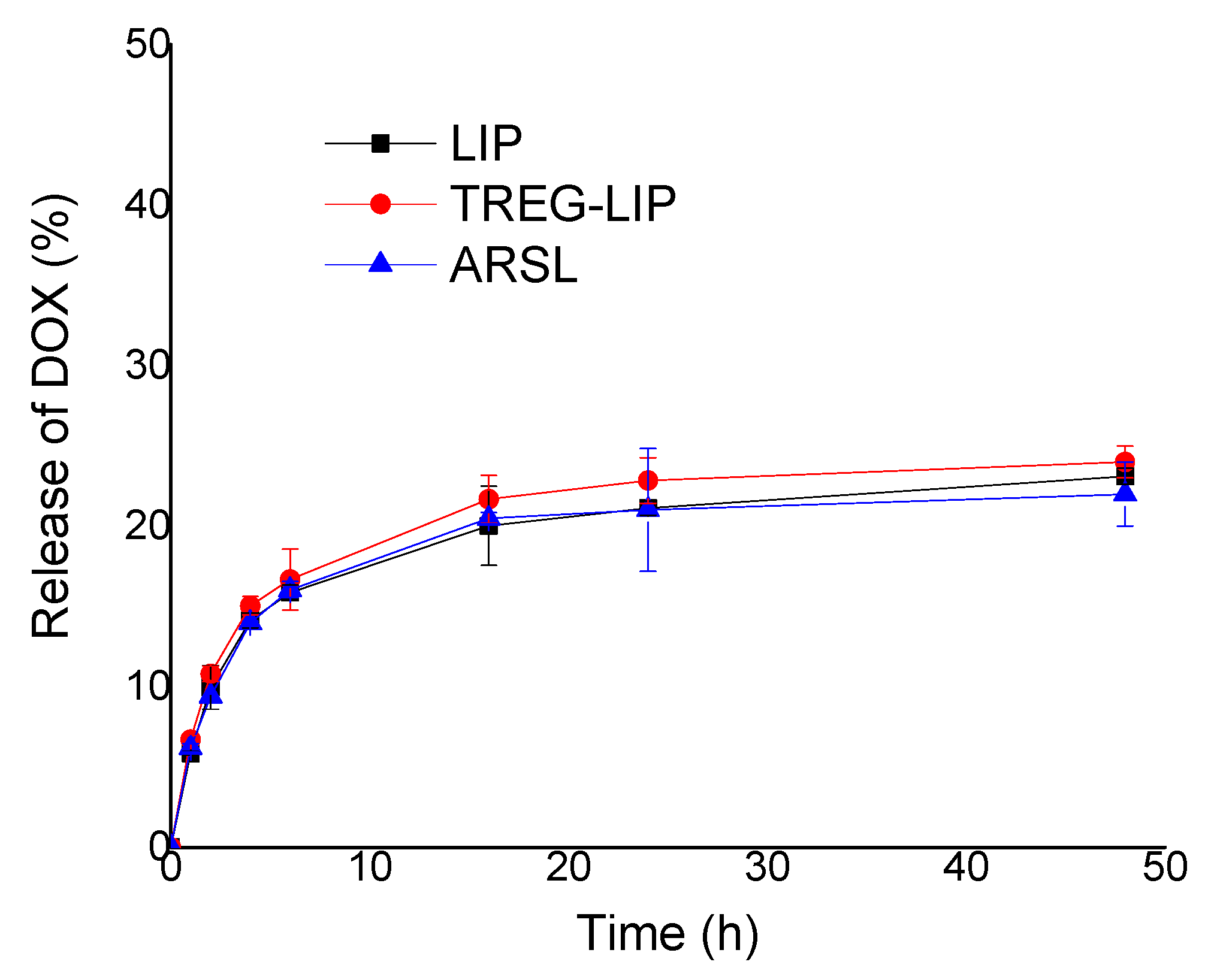
The release of DOX from TREG-LIP, ARSL, and LIP (as control formulation) was studied by adding 0.5 mL of DOX-loaded liposome formulations at a lipid concentration of 1 mg/mL in dialysis tubing sacs (Servapor, with MW cutoff 14,000 Daltons) and placing the sacs in capped test tubes containing 10 mL of PBS buffer, pH 7.40. The test tubes were then closed and placed in a shaking incubator (Stuart Orbital Incubator) at 60 rpm and 37 °C for 48 h. At specified time points (0, 1, 2, h, 6, 18, 24, 48 h) 0.5 mL samples were taken from the buffer (volume was replaced with PBS) and DOX was quantified by measuring the sample FI, as described above. Sink conditions applied throughout the study.
2.5. Cell Culture Studies
Three types of cells were used in this study: (i) C57BL/6 mouse B16F10 skin melanoma cells (B16) (National Cancer Institute Tumor Repository, Frederick, MD, USA); (ii) Lewis lung carcinoma cells (LLC), and (iii) Human HEK-293 embryonic kidney cells (HEK) (American Type Culture Collection, Manassas, VA, USA), provided by Prof. G.T. Stathopoulos (Medical School, University of Patras). All cells were grown in RPMI 1640 medium supplemented with 10% FBS and 1% antibiotic-antimycotic solution (Invitrogen, Carlsbad, CA, USA). The cells were cultured at 37 °C, 5% CO2/saturated humidity. Medium was changed every 2–3 days.
The toxicity of the various types of liposomes towards cancer and normal cells was evaluated by the corresponding reduction of cell viability after 24 h incubation with test (DOX-ARSL) or control formulations (DOX-solution, empty ARSL, and, DOX-solution—empty ARSL mixtures), by the MTT assay [
29]. In more detail, for each experiment, cells were seeded overnight at 37 °C at a density of 3 × 10
4 cells per well, in 24-well plates, until almost confluent, and then incubated for 24 h at 37 °C (5% CO
2/saturated humidity) with 0.4 mL RPMI and 0.1 mL of the formulation evaluated in each case: (i) DOX-solution; (ii) DOX-ARSL; (iii) Empty ARSL (with the same final lipid concentrations with (ii)); and (iv) PBS (control). All formulations were pre-filtrated through a 0.22 μm Millipore filter. The medium/PBS (
v/
v) ratio was kept constant. Cell viability was determined by the MTT method, which is a colorimetric assay for assessment of the cell metabolic activity. For this, after the 24 h incubations with the cells, the medium was removed and the cells were washed three times with PBS before adding 0.5 mL of a 0.5 mg/mL solution of MTT in PBS. Two hours later, 0.5 mL of acidified isopropanol (0.33%) was added in each well, in order to disrupt the cells and solubilize the colored formazan crystals that formed. The optical density of the controls and samples was measured at 570 nm (Multiscan EX plate reader, Thermo). Viability (%) was calculated by the equation:
where, OD-570
control corresponds to untreated cells (or PBS control) and OD-570
background to MTT without cells. 1% Triton X-100 was used as a positive control of cytotoxicity, and resulted in viability values <5% for all cells.
For the cytotoxicity studies, the lipid/DOX ratios used during the loading of DOX into the liposomes were different from the ratio mentioned above (in
Section 2.4), in order to achieve the specific DOX and ARSL concentrations required for induction of significant toxicity towards cancer cells. Test experiments were initially carried out for selection of the specific concentrations required, and finally the DOX concentrations selected were 1 µM and 3 µΜ, and the phospholipid concentrations of ARSL (empty or DOX-loaded) ranged between 0.5 µΜ and 50 µΜ.
2.6. DOX Uptake by Cells
The uptake of DOX by B16 and LLC cells was measured. For this, cells were seeded overnight at 5 × 104 cells/mL in RPMI at 37 °C (5% CO2/saturated humidity), and then incubated with DOX-liposomes or free DOX (control) for 3 h. DOX concentrations of 1 µM and 3 μM were used in the uptake experiments (after confirming that they are non-toxic towards the cells following a 3 h co-incubation period). After treatment of the cells with DOX-liposomes or DOX solution, the cells were washed twice with ice-cold PBS, and then detached and lysed by the addition of 3 mL of PBS and 2 mL of Triton X-100 (10%). The samples were collected and their fluorescence intensity was measured (EX 485 and EM 590 nm), for calculation of the DOX concentration from a calibration curve that was constructed by spiking known concentrations of DOX (between 0.25 µΜ and 4.5µM in similar media with that of the samples (Triton X-100 in PBS); the presence of lysed cells or liposomes did not modulate the FI of DOX control solutions (in the concentration range used). The auto-fluorescence of non-treated cells was also measured under identical conditions, and found to be very low (always below 0.1% of the FIs of the samples). The protein concentration in every sample was measured by the Bradford microassay.
Finally, DOX uptake by cells was expressed as % Uptake (by comparison of the DOX measured in the cells and the total amount of DOX incubated with the cells), and also as nmoles DOX/μg protein.
2.7. Transmission Electron Microscopy (TEM)
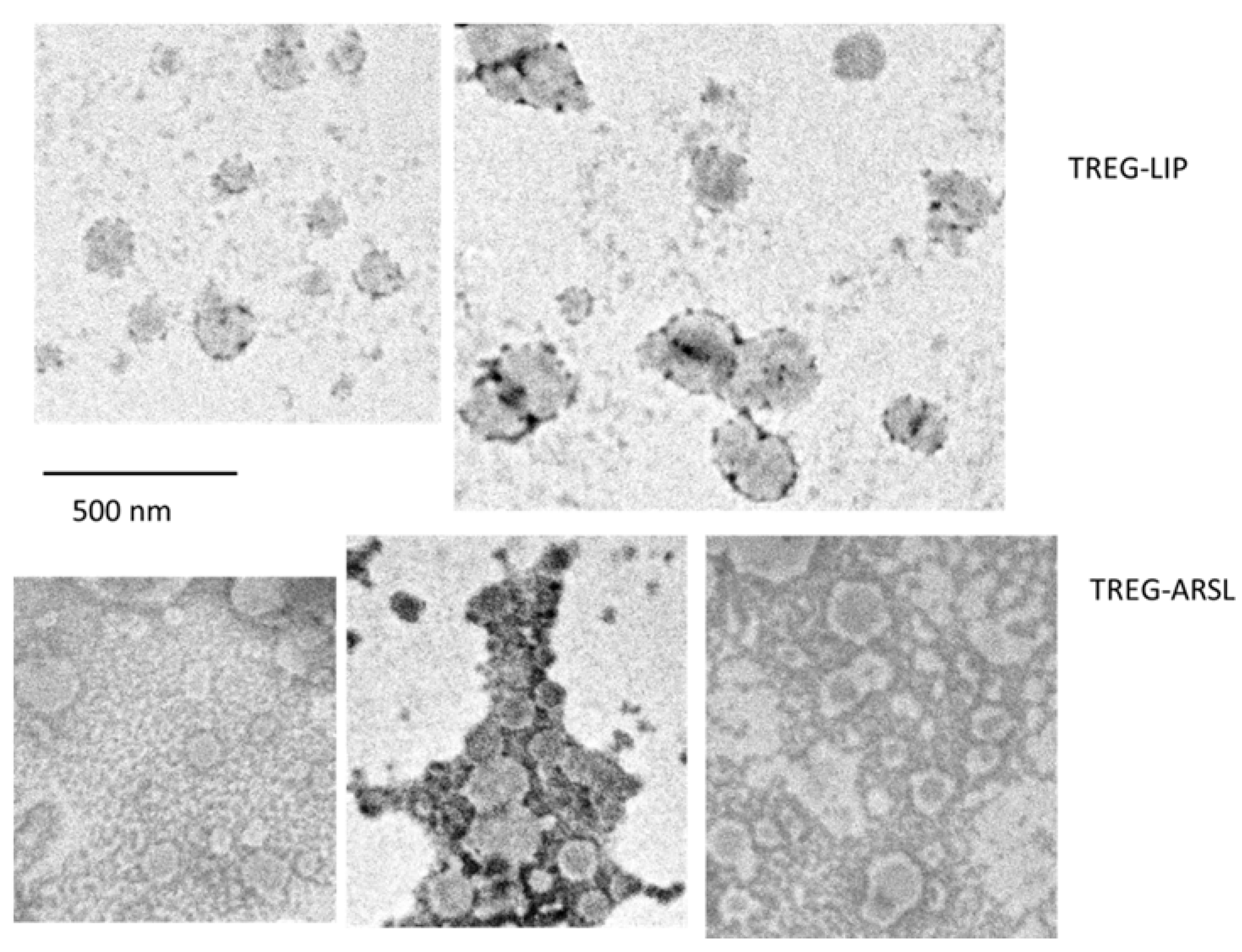
The morphology of ARSL and TREG-ARSL was studied by TEM after negative staining with 1% neutral phosphotungstic acid (PTA), washing twice with dH2O, and draining with a tip of a tissue paper. Samples were observed at 100.000 eV with JEOL (JEM-2100) TEM (Jeol, Tokyo, Japan)
2.8. Statistical Analysis
All results are expressed as mean ± SD from at least three independent experiments. The significance of variability between results from various groups was determined by two-way-ANOVA and individual differences between groups were tested by one-way ANOVA and Tukey’s multiple comparisons test.
4. Discussion
Herein, we investigated the potential to incorporate TREG (a previously synthesized and studied lipidic derivative of curcumin) [
18] in ARSL and its effect on ARSL stability. Furthermore, we evaluated the ability to load ARSL and TREG-ARSL with DOX using the remote liposome loading method, in order to develop potential anticancer chemotherapeutic systems that will combine the anticancer activities of ARSL and DOX, together with the potentially enhanced pharmacokinetics and targeting capability provided by TREG.
TREG-LIP and TREG-ARSL, as well as LIP and ARSL (which were prepared as control vesicles), had mean diameters in the nano-range, between 101 and 145 nm (
Table 2), polydispersity values below <0.270, and slightly negative zeta potential values. The low zeta potential value measured for ARSL despite the highly negative charge of arsonolipids, was attributed to the fact that the particular ARSL were PEGylated, and was consistent with previous results [
9]. For the liposomes with no arsonolipids in their composition, the differences with other reported values [
30] were attributed to the different measurement conditions used [
31]. The complete incorporation of TREG in the liposome membranes was verified by HPLC measurements that were carried out before and after purification of the lipid dispersions from non-liposome associated materials (by size exclusion chromatography) (
Figure S2), ruling out any suspicion that TREG micelles or aggregates could be present in the formulations, as also evident in the analytical size reports of DLS measurements (
Figure S1), and their morphology (
Figure 1).
After confirming the capability to formulate stable nanosized TREG-ARSL that incorporate the complete amount of TREG added in the lipid phase during their preparation, and demonstrate high physical stability during storage at 4 °C (
Figure 2), their integrity in the presence of serum proteins was evaluated. The current results demonstrated that the incorporation of TREG in ARSL had a marginal effect to increase their stability towards any disruptive actions of serum proteins, and confirmed previous findings about the stabilizing (towards disruption by serum proteins) effect of TREG on TREG-LIP [
20,
21].
Concerning the capability of using the active loading protocol [
15,
16] to load DOX into ARSL and TREG-ARSL, the current results (
Figure 5) confirmed for the first time that high amounts of DOX can be loaded into ARSL, opening the path for exploration of potential synergistic anticancer effects of the two compounds when co-existing in a vesicular format. Additionally, it was proven that when liposomes or ARSL incorporate TREG, this does not prevent in any way their capability to be remotely loaded with DOX. On the contrary, high DOX loadings were achieved faster in TREG-LIP and ARSL compared to the corresponding control vesicles. Whether the latter results are attributed to modulations in the liquid ordered (LO) and liquid disordered transition of the membranes, or to the decrease of the hydrophilic properties of the vesicle surface that may result in faster attraction of DOX molecules into the lipid membrane, or perhaps to chemical interactions (e.g., hydrogen bonding or dipole-dipole interactions) between specific groups of the TREG and/or the arsonolipid with DOX, we cannot be sure. Importantly, the release profile of DOX from all the vesicle types was found to be similar, proving that TREG or arsonolipids do not cause faster (compared with DOX-LIP) leakage of DOX (
Figure 6).
The in vitro experiments carried out with DOX-ARSL on B16, LLC and HEK cells demonstrated interesting anticancer potential of DOX-ARSL toward both cancer cell types at specific DOX and ARSL combinations (
Figure 7). In most cases, the cytotoxic effect of DOX-ARSL towards cancer cells was higher than that demonstrated when the corresponding mixtures of free DOX and empty ARSL were used, indicating the importance of combining the two compounds in the same vesicular structure. Oppositely, the situation was different for HEK cells, since DOX-ARSL (at all concentrations) were found to be less toxic compared to the corresponding mixtures. The previous finding, together with the fact that in all cases DOX-ARSL were less toxic towards HEK cells and significantly more toxic towards the cancer cells (the less resistant LLC cells as well as the more resistant B16 cells) indicate some very strong advantages of using the particular formulations as anticancer therapeutics.
The uptake of free DOX by both cell types is substantially higher compared to that of ARSL-entrapped DOX (
Table 3), suggesting that the cytotoxic effect of the comparatively low amounts of DOX taken-up by the cells (when they are incubated with DOX-ARSL) is highly enhanced when appropriate amounts of arsonolipids are taken up at the same time (in the form of DOX-ARSL).
In order to understand if the current results indicate synergism, additive effect or antagonism between DOX and ARSL, we calculated the corresponding combination index (CI), for each case, according to the previously reported equation: CI = (D1/Dm1) + (D2/Dm2) [
22,
32]. Here, D1 and D2 represent the dose of the drugs in combined administration (DOX-ARSL) and Dm1 and Dm2 represent the dose required to produce the same effect when used alone (free DOX or empty ARSL). It has been reported that when CI < 1, synergism is indicated; when CI = 1, additive nature is indicated; and when CI > 1, antagonism is indicated [
32,
33]. The CI values that could be calculated from the current results are reported in
Table 4. The values for the free DOX and empty ARSL (Dm1 and Dm2) were taken from the corresponding results of cell viability in presence of increasing concentrations of free compounds (
Figure S3, Supplementary Data file). For the B16 cells, some values for the free compounds were not available, since even at very high concentrations of empty ARSL, the viability of B16 cells was never below 45–50%; therefore, some CI values could not be estimated. As seen in
Table 4, in two cases for B16 cells and in one case for LLC cells, the CI values indicate synergism between arsonolipids and DOX; while in other cases in LLC cells, antagonism is indicated.
The current preliminary in vitro studies are not sufficient to fully explain the mechanisms of the relative cytotoxic effects demonstrated by DOX-ARSL towards cancer and normal cells. Nevertheless, the highly interesting current finding in regards to the cytotoxic properties of DOX-ARSL towards cancer cells and their reduced toxicity towards normal (HEK) cells fully justify further in vivo exploitation of this particular combination therapy nano-system. Furthermore, the fact that the particular (DOX-loaded) ARSL can also incorporate TREG in their membranes, with a slight improvement in their in vitro integrity during incubation in the presence of proteins, unlocks a series of potential applications for difficult-to-treat pathologies, such as brain tumors, that should be additionally investigated in future projects. In fact, the additive or synergistic anticancer activity of DOX/curcumin combinations suggested by several recent studies [
34,
35,
36,
37] provide an extra reason to further investigate the anticancer activity of DOX-loaded TREG-ARSL.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}