Identification of Catalposide Metabolites in Human Liver and Intestinal Preparations and Characterization of the Relevant Sulfotransferase, UDP-glucuronosyltransferase, and Carboxylesterase Enzymes

 , , ,
, , ,  , and
, and 
Abstract
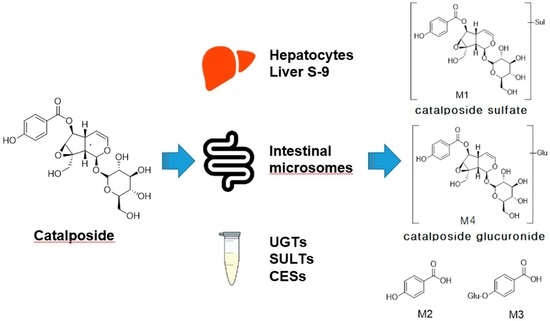
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. In Vitro Metabolism of Catalposide in Cryopreserved Human Hepatocytes
2.3. In Vitro Metabolism of Catalposide in Human Liver S9 Fractions and Intestinal Microsomes
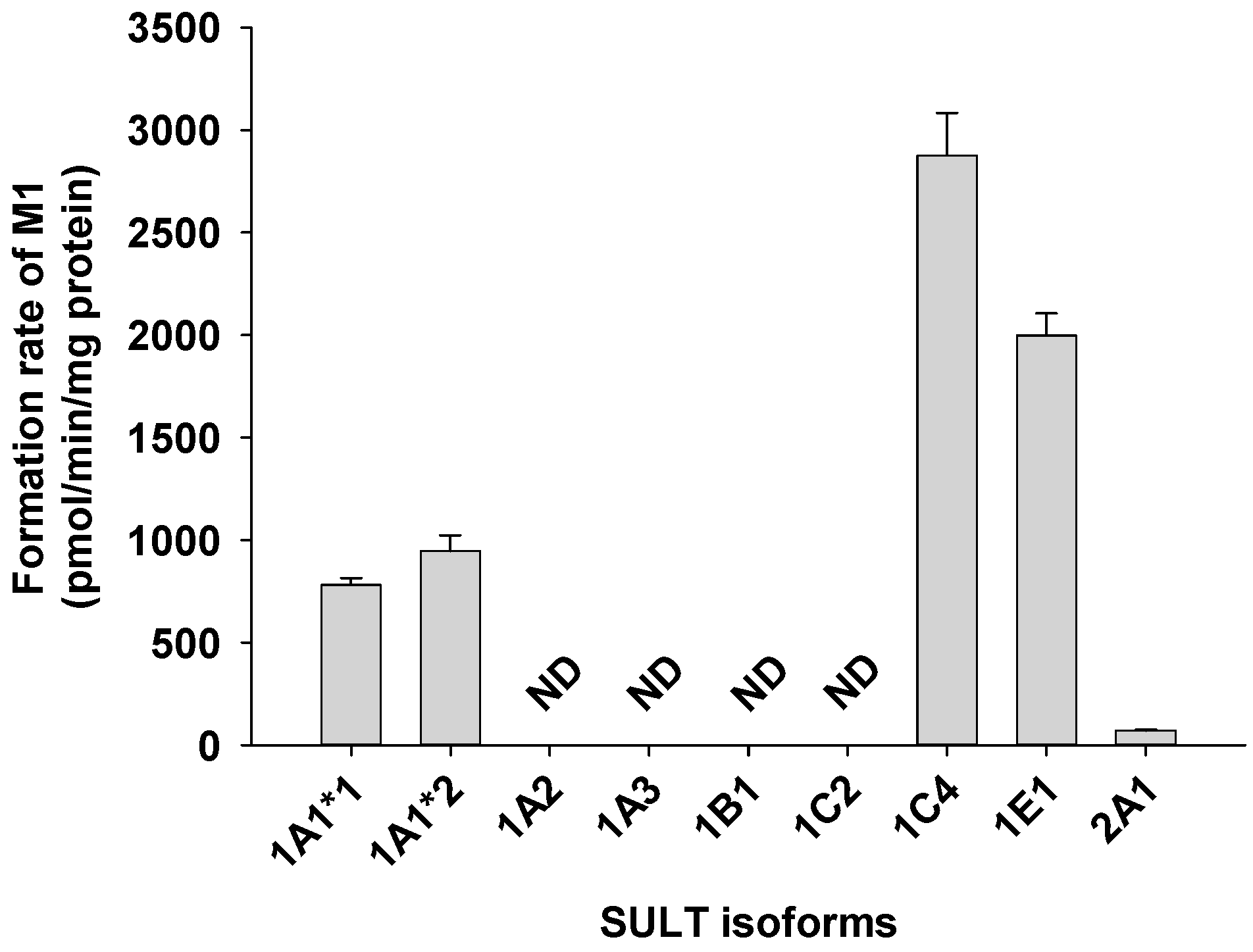
2.4. Characterization of Human SULTs Involved in Catalposide Sulfation
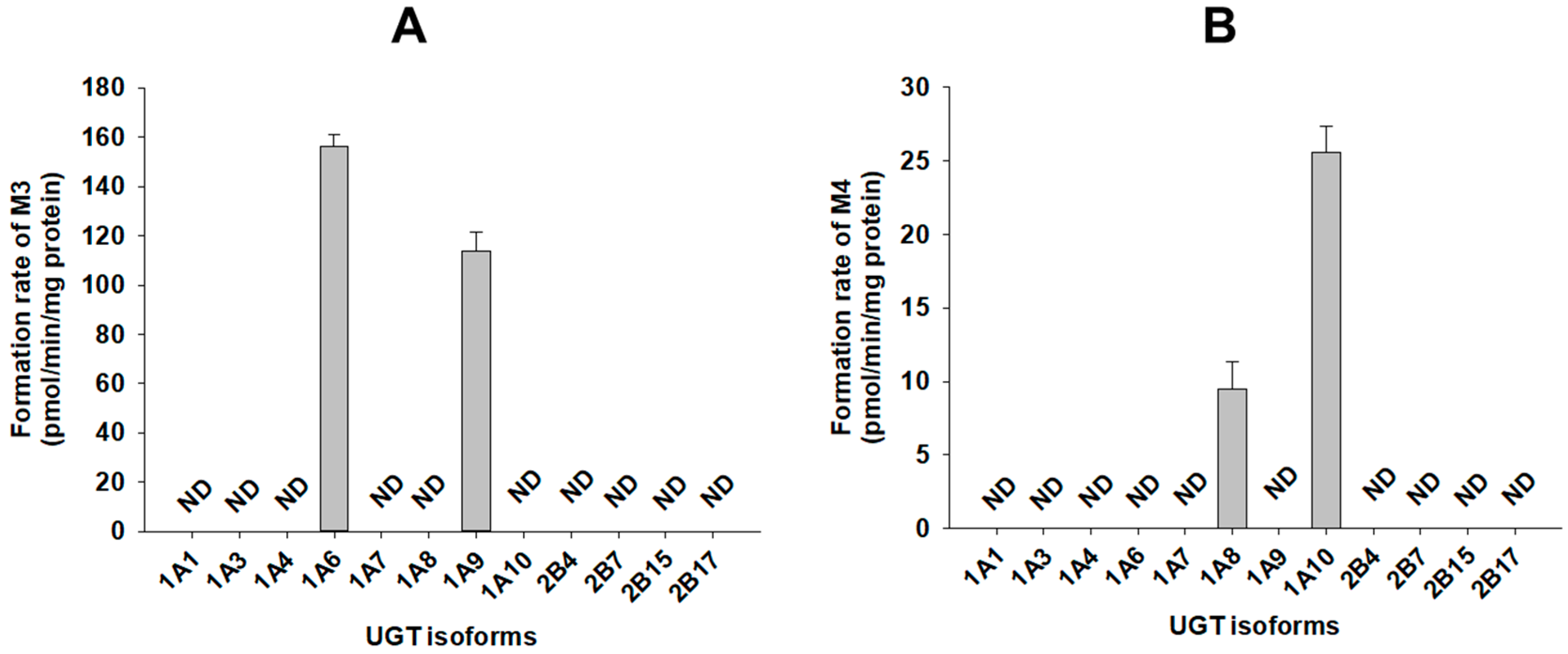
2.5. Characterization of Human UGTs Involved in Catalposide and 4-Hydroxybenzoic Acid Glucuronidation
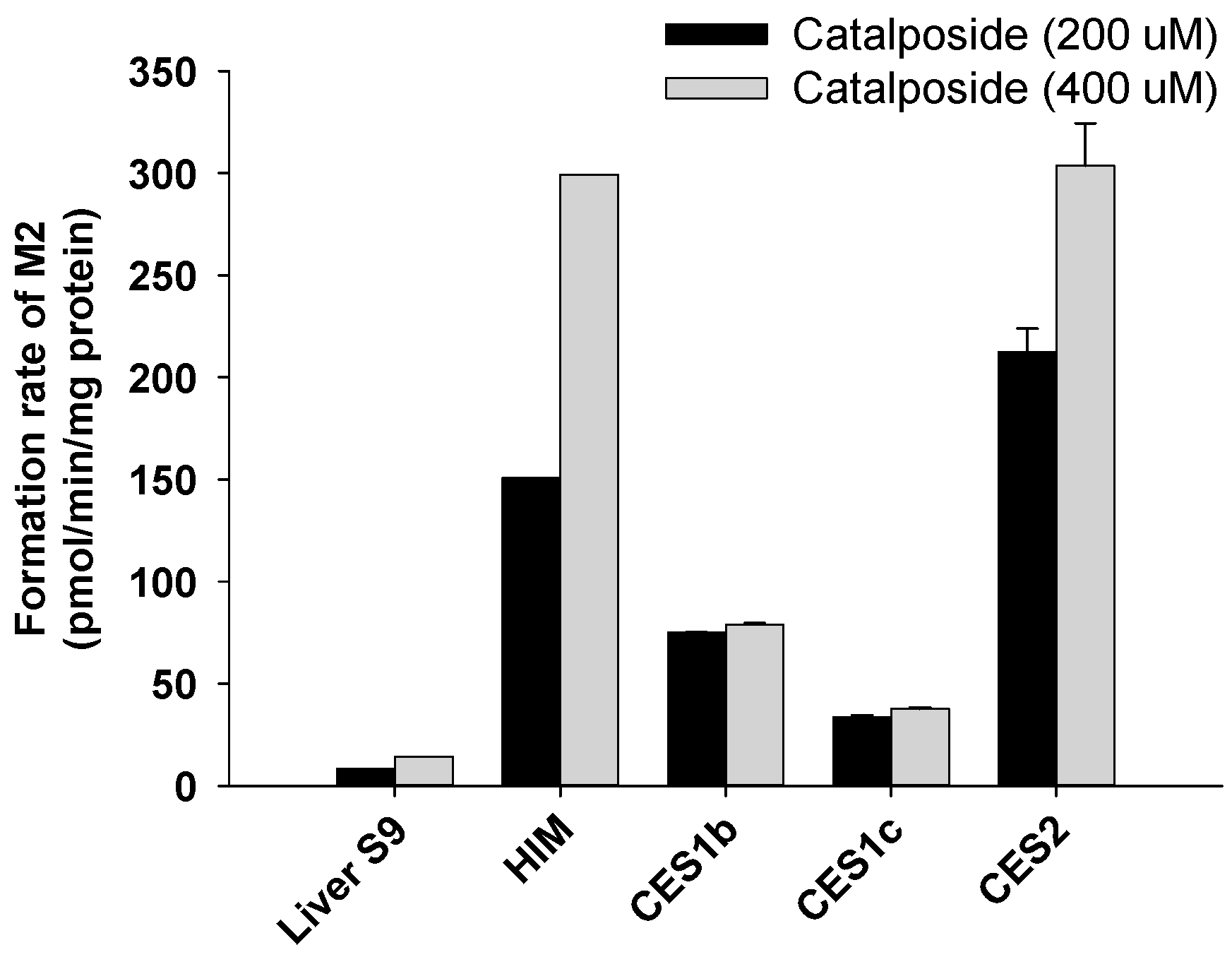
2.6. Characterization of Carboxylesterases Involved in the Formation of 4-Hydroxybenzoic Acid from Catalposide
2.7. LC-HRMS Analysis of Catalposide and Metabolites
2.8. Data Analysis
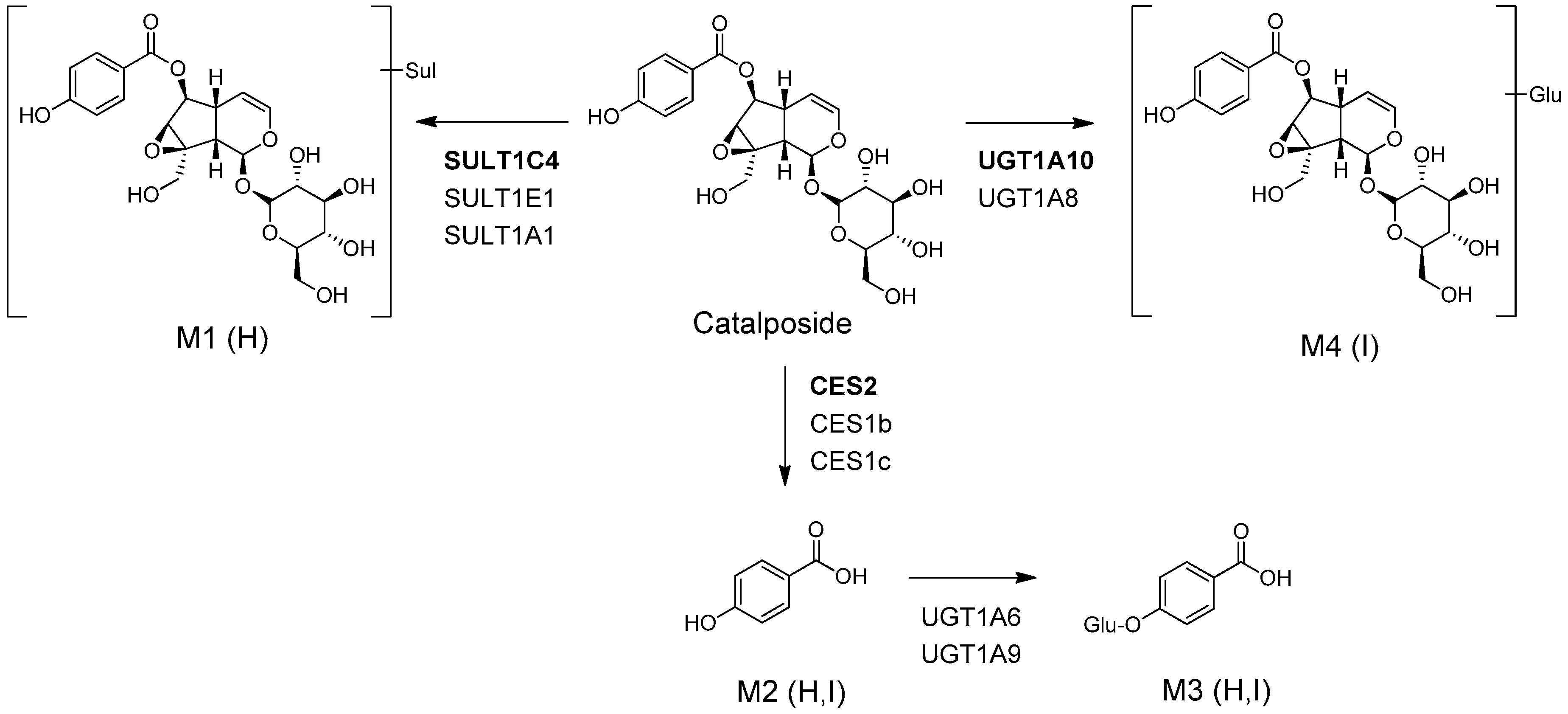
3. Results
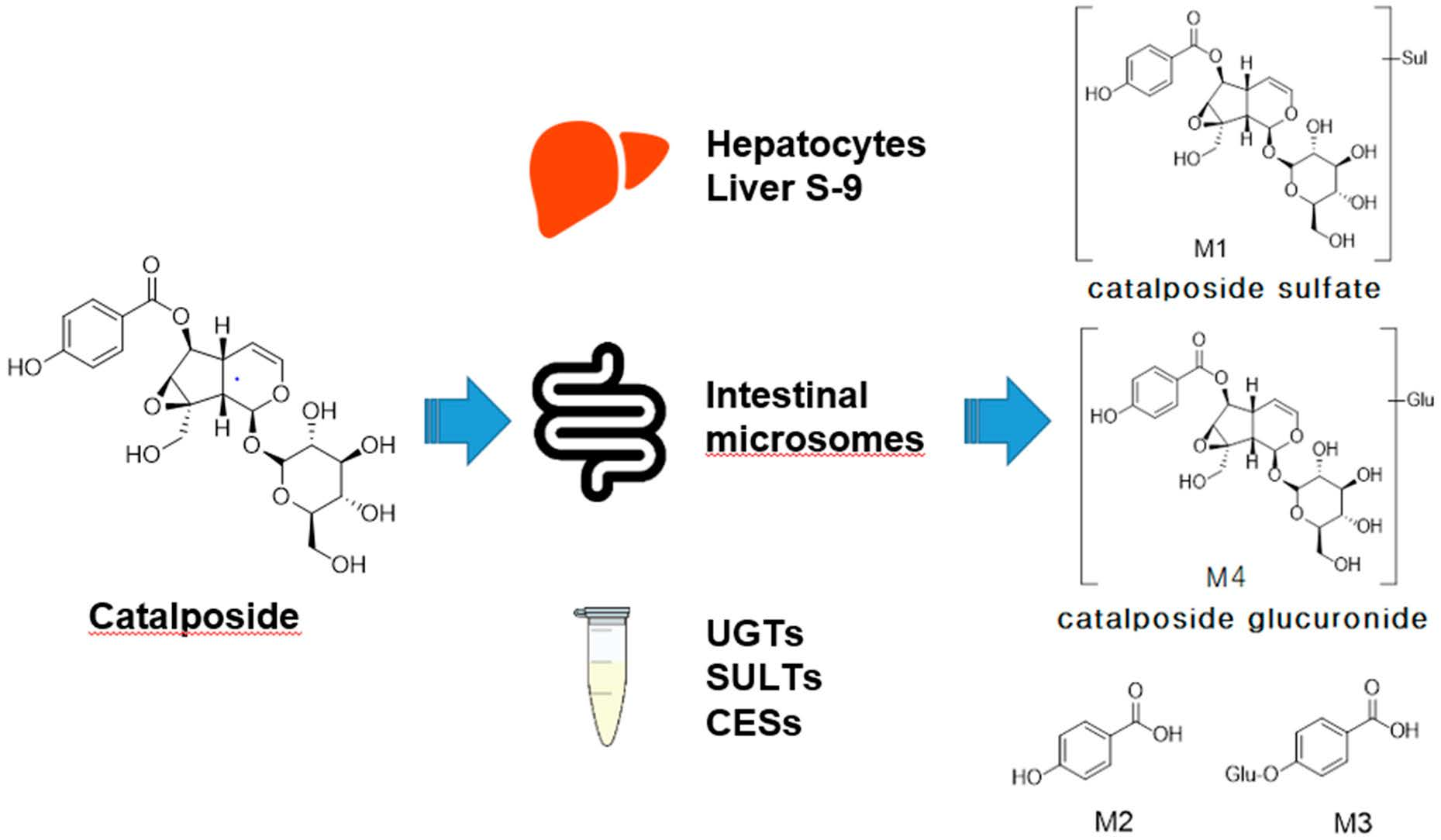
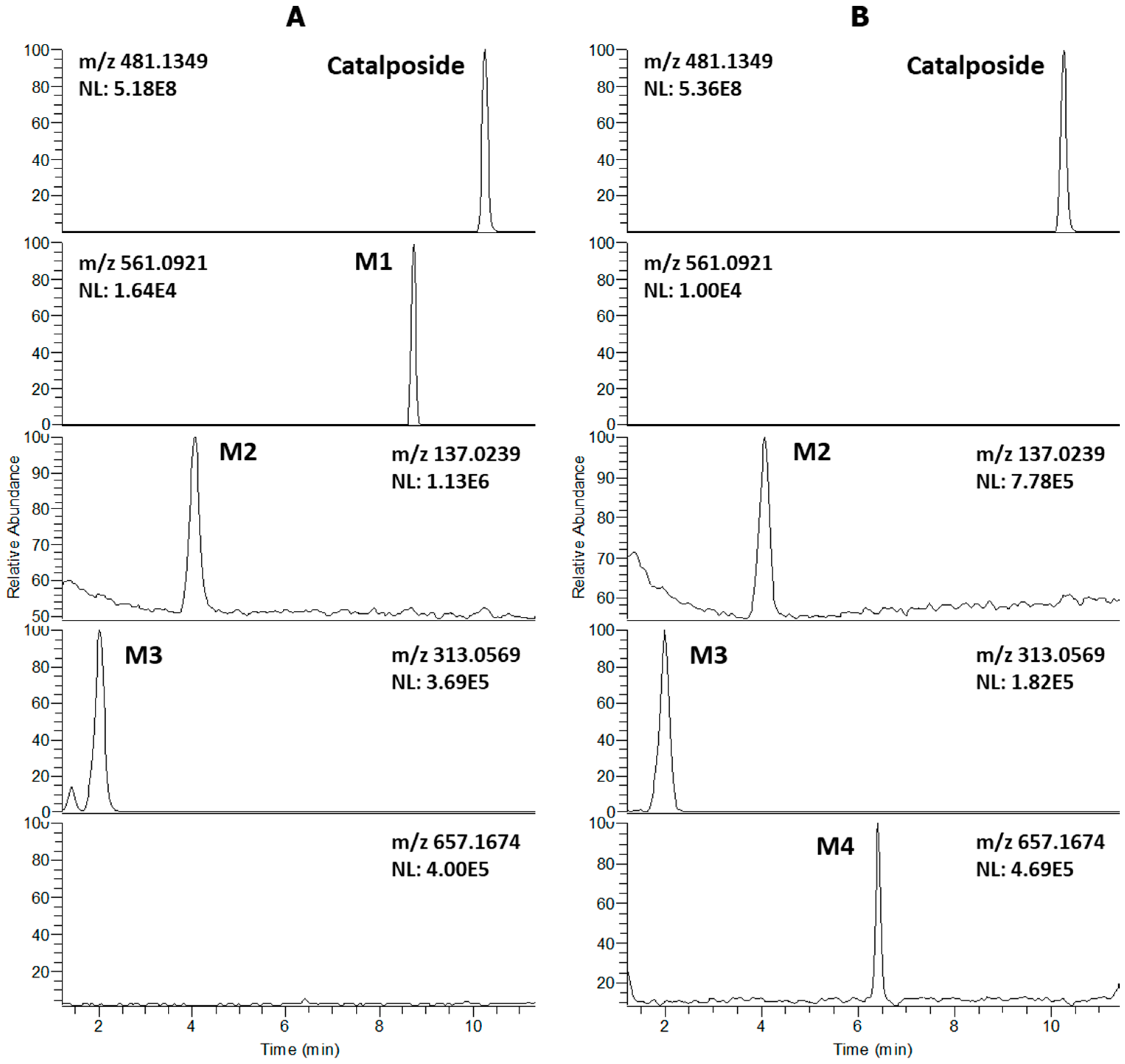
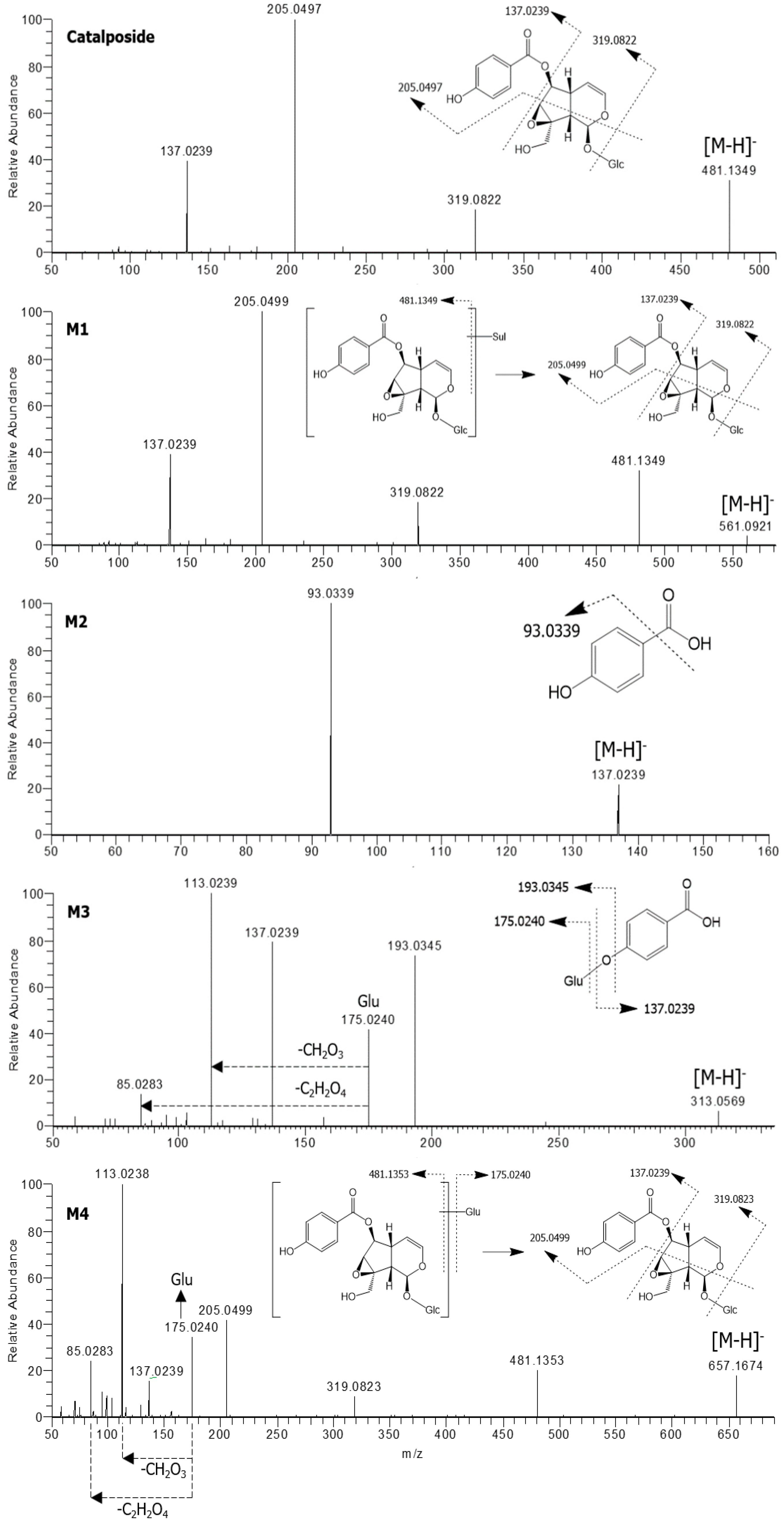
3.1. In Vitro Metabolic Profiles of Catalposide Incubated with Human Hepatocytes and Intestinal Microsomes
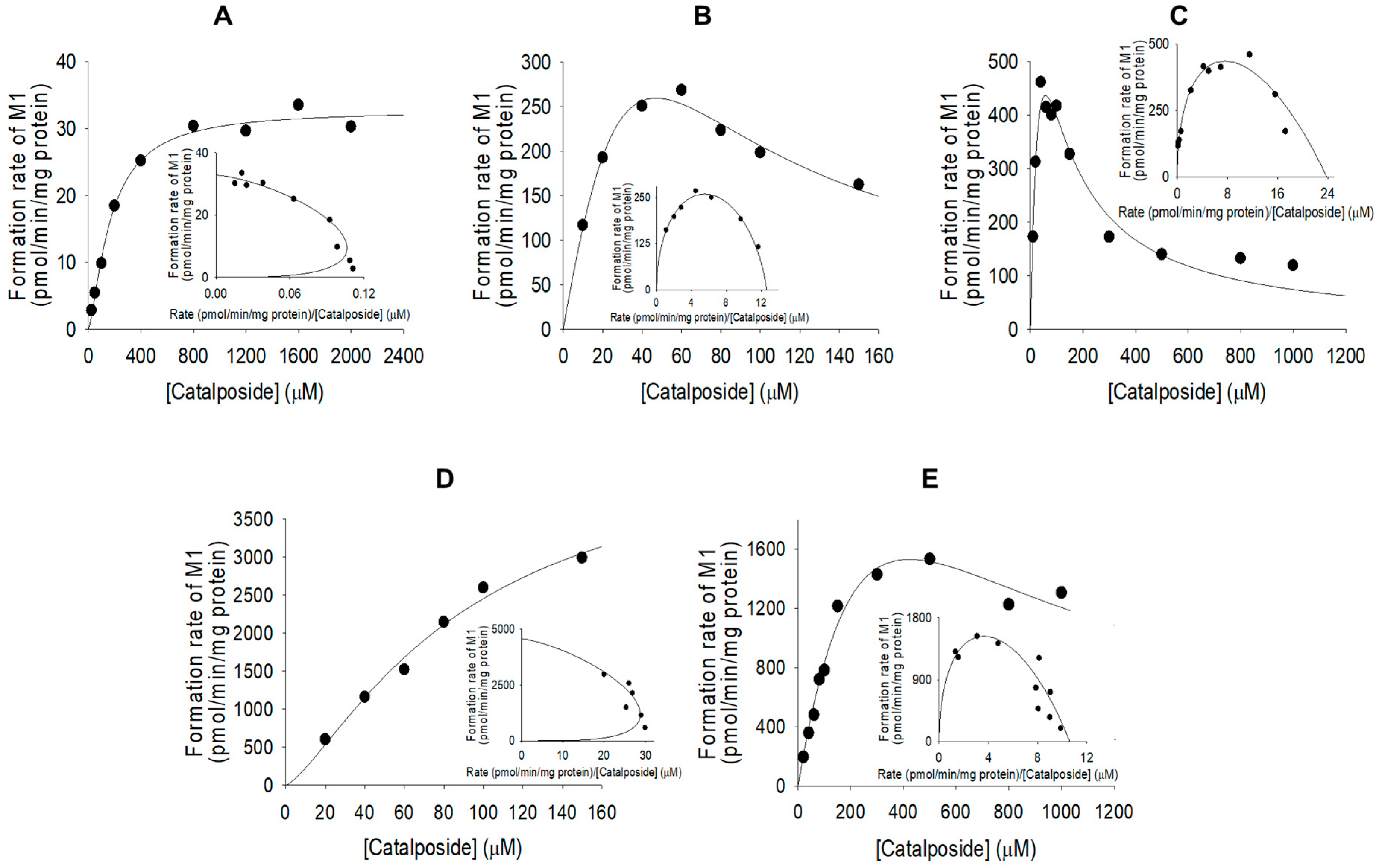
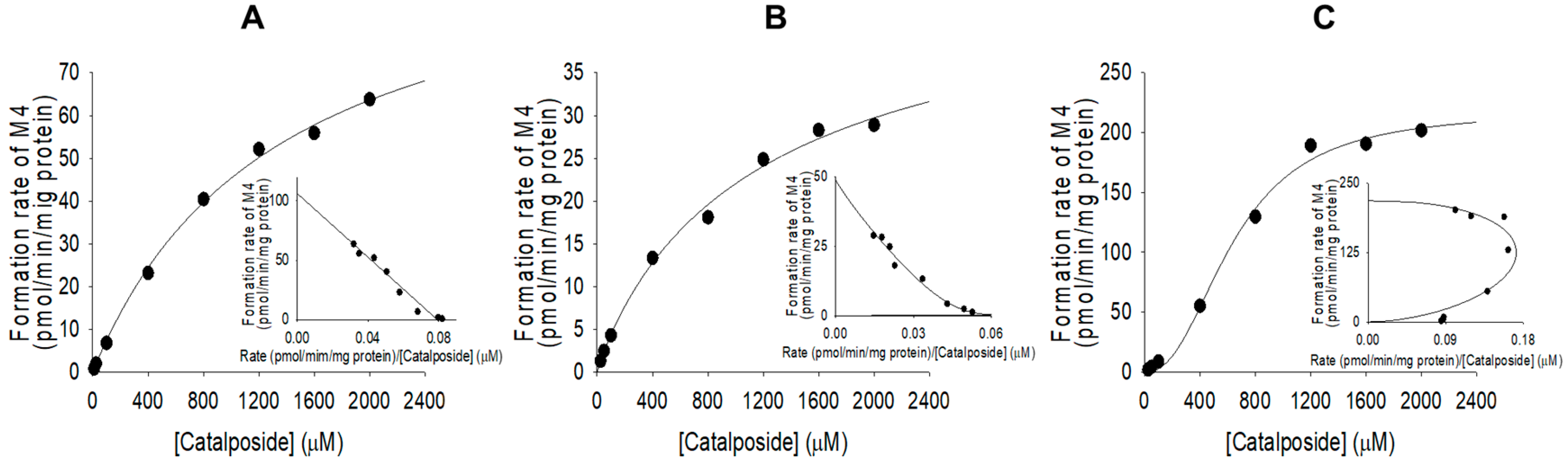
3.2. Characterization of Human SULT, UGT, and CES Enzymes Involved in Catalposide Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Harput, U.S.; Saracoglu, I.; Inoue, M.; Ogihara, Y. Phenylethanoid and Iridoid Glycosides from Veronica persica. Chem. Pharm. Bull. 2002, 50, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Harput, U.S.; Saracoglu, I.; Nagatsu, A.; Ogihara, Y. Iridoid Glucosides from Veronica hederifolia. Chem. Pharm. Bull. 2002, 50, 1106–1108. [Google Scholar] [CrossRef] [PubMed]
- Harput, U.; Varel, M.; Nagatsu, A.; Saracoglu, I. Acylated iridoid glucosides from Veronica anagallis-aquatica. Phytochemistry 2004, 65, 2135–2139. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Pae, H.O.; Oh, G.S.; Lee, S.Y.; Chai, K.Y.; Song, C.E.; Kwon, T.O.; Chung, H.T.; Lee, H.S. Inhibition of Inducible Nitric Oxide Synthesis by Catalposide from Catalpa ovata. Planta Med. 2002, 68, 685–689. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Pae, H.; Oh, G.; Choi, B.; Jeong, S.; Jang, S.; Oh, H.; Kwon, T.; Song, C.E.; Chung, H. Inhibition of TNF-α, IL-1β, and IL-6 productions and NF-κB activation in lipopolysaccharide-activated RAW 264.7 macrophages by catalposide, an iridoid glycoside isolated from Catalpa ovata G. Don (Bignoniaceae). Int. Immunopharmacol. 2002, 2, 1173–1181. [Google Scholar] [CrossRef]
- Moon, M.K.; Choi, B.M.; Oh, G.S.; Pae, H.O.; Kim, J.D.; Oh, H.; Oh, C.S.; Kim, D.H.; Rho, Y.D.; Shin, M.K.; et al. Catalposide protects Neuro 2A cells from hydrogen peroxide-induced cytotoxicity via the expression of heme oxygenase-1. Toxicol. Lett. 2003, 145, 46–54. [Google Scholar] [CrossRef]
- Kim, S.W.; Choi, S.C.; Choi, E.Y.; Kim, K.S.; Oh, J.M.; Lee, H.J.; Oh, H.M.; Kim, S.; Oh, B.S.; Kimm, K.C.; et al. Catalposide, a Compound Isolated from Catalpa Ovata, Attenuates Induction of Intestinal Epithelial Proinflammatory Gene Expression and Reduces the Severity of Trinitrobenzene Sulfonic Acid-Induced Colitis in Mice. Inflamm. Bowel Dis. 2004, 10, 564–572. [Google Scholar] [CrossRef]
- Kupeli, E.; Harput, U.S.; Varel, M.; Yesilada, E.; Saracoglu, I. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. J. Ethnopharmacol. 2005, 102, 170–176. [Google Scholar] [CrossRef]
- Lu, Q.; Sun, Y.; Shu, Y.; Tan, S.; Yin, L.; Guo, Y.; Tang, L. HSCCC Separation of the Two Iridoid Glycosides and Three Phenolic Compounds from Veronica ciliata and Their in Vitro Antioxidant and Anti-Hepatocarcinoma Activities. Molecules 2016, 21, 1234. [Google Scholar] [CrossRef]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharmacal Res. 2009, 32, 207–213. [Google Scholar] [CrossRef]
- Saracoglu, I.; Harput, U.S. In vitro cytotoxic activity and structure activity relationships of iridoid glucosides derived from Veronica species. Phytother. Res. 2012, 26, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jun, H.J.; Hoang, M.H.; Jia, Y.; Han, X.H.; Lee, D.H.; Lee, H.J.; Hwang, B.Y.; Lee, S.J. Catalposide is a natural agonistic ligand of peroxisome proliferator-activated receptor-α. Biochem. Biophys. Res. Commun. 2012, 422, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Lu, Q.; Shu, Y.; Sun, Y.; Chen, F.; Tang, L. Iridoid Glycosides Fraction Isolated from Veronica ciliata Fisch. Protects against Acetaminophen-Induced Liver Injury in Mice. Evid. Based Complement. Alternat. Med. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.Y.; Lee, H.W.; Kim, H.H.; Lee, H.I.; Chung, H.T.; Lee, H.S. Determination of Catalposide in Rat Plasma by Liquid Chromatography-Mass Spectrometry. Anal. Lett. 2003, 36, 2999–3009. [Google Scholar] [CrossRef]
- Jeong, H.U.; Kwon, M.; Lee, Y.; Yoo, J.S.; Shin, D.H.; Song, I.S.; Lee, H.S. Organic anion transporter 3- and organic anion transporting polypeptides 1B1- and 1B3-mediated transport of catalposide. Drug Des. Dev. Ther. 2015, 9, 643–653. [Google Scholar]
- Choi, W.G.; Kim, J.H.; Kim, D.K.; Lee, Y.; Yoo, J.S.; Shin, D.H.; Lee, H.S. Simultaneous Determination of Chlorogenic Acid Isomers and Metabolites in Rat Plasma Using LC-MS/MS and Its Application to A Pharmacokinetic Study Following Oral Administration of Stauntonia Hexaphylla Leaf Extract (YRA-1909) to Rats. Pharmaceutics 2018, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Cerny, M.A. Prevalence of Non-Cytochrome P450-Mediated Metabolism in Food and Drug Administration-Approved Oral and Intravenous Drugs: 2006–2015. Drug Metab. Dispos. 2016, 44, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Foti, R.S.; Dalvie, D.K. Cytochrome P450 and non-Cytochrome P450 Oxidative Metabolism: Contributions to the Pharmacokinetics, Safety and Efficacy of Xenobiotics. Drug Metab. Dispos. 2016, 44, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hwang, D.K.; Moon, J.Y.; Lee, Y.; Yoo, J.S.; Shin, D.H.; Lee, H.S. Multiple UDP-Glucuronosyltransferase and Sulfotransferase Enzymes are Responsible for the Metabolism of Verproside in Human Liver Preparations. Molecules 2017, 22, 670. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.U.; Kim, J.H.; Kong, T.Y.; Choi, W.G.; Lee, H.S. Comparative metabolism of honokiol in mouse, rat, dog, monkey, and human hepatocytes. Arch. Pharmacal Res. 2016, 39, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Suiko, M.; Kurogi, K.; Hashiguchi, T.; Sakakibara, Y.; Liu, M.C. Updated perspectives on the cytosolic sulfotransferases (SULTs) and SULT-mediated sulfation. Biosci. Biotech. Biochem. 2017, 81, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidry, A.L.; Tibbs, Z.E.; Runge-Morris, M.; Falany, C.N. Expression, Purification, and Characterization of Human Cytosolic Sulfotransferase (Sult) 1c4. Horm. Mol. Boil. Clin. Investig. 2017, 29, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Coughtrie, M.W. Function and organization of the human cytosolic sulfotransferase (SULT) family. Chem. Interact. 2016, 259, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W.H. Quantitative Evaluation of the Expression and Activity of Five Major Sulfotransferases (SULTs) in Human Tissues: The SULT “Pie”. Drug Metab. Dispos. 2009, 37, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Marto, N.; Morello, J.; Monteiro, E.C.; Pereira, S.A.; Morello, J.B. Implications of Sulfotransferase Activity in Interindividual Variability in Drug Response: Clinical Perspective on Current Knowledge. Drug Metab. Rev. 2017, 49, 357–371. [Google Scholar] [CrossRef] [PubMed]
- James, M.O.; Ambadapadi, S. Interactions of cytosolic sulfotransferases with xenobiotics. Drug Metab. Rev. 2013, 45, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Chen, Y.; Huang, C.; Chen, G. Caffeine Induction of Sulfotransferases in Rat Liver and Intestine. J. Appl. Toxicol. 2012, 32, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Zhang, J.B.; Hou, J.; Dou, T.Y.; Ge, G.B.; Hu, W.Z.; Yang, L. Chemical probes fpr human UDP-glucuronosyltransferases: A comprehensive review. Biotechnol. J. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, G.; Wang, G.; Hao, H. Regulation of Mammalian UDP-Glucuronosyltransferases. Curr. Drug Metab. 2018, 19, 490–501. [Google Scholar] [CrossRef]
- Meech, R.; Hu, D.G.; McKinnon, R.A.; Mubarokah, S.N.; Haines, A.Z.; Nair, P.C.; Rowland, A.; MacKenzie, P.I. The UDP-Glycosyltransferase (UGT) Superfamily: New Members, New Functions, and Novel Paradigms. Physiol. Rev. 2019, 99, 1153–1222. [Google Scholar] [CrossRef]
- Fallon, J.K.; Neubert, H.; Goosen, T.C.; Smith, P.C. Targeted Precise Quantification of 12 Human Recombinant Uridine-Diphosphate Glucuronosyl Transferase 1A and 2B Isoforms Using Nano-Ultra-High-Performance Liquid Chromatography/Tandem Mass Spectrometry with Selected Reaction Monitoring. Drug Metab. Dispos. 2013, 41, 2076–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argikar, U.A.; Potter, P.M.; Hutzler, J.M.; Marathe, P.H. Challenges and opportunities with Non-CYP enzymes aldehyde oxidase, carboxylesterase, and UDP-Glucuronosyltransferase: Focus on reaction phenotyping and prediction of human clearance. AAPS J. 2016, 18, 1391–1405. [Google Scholar] [CrossRef] [PubMed]
- Kiang, T.K.; Ensom, M.H.; Chang, T.K. UDP-glucuronosyltransferases and clinical drug-drug interactions. Pharmacol. Ther. 2005, 106, 97–132. [Google Scholar] [CrossRef]
- Miners, J.O.; Mackenzie, P.I.; Knights, K.M. The prediction of drug-glucuronidation parameters in humans: UDP-glucuronosyltransferase enzyme-selective substrate and inhibitor probes for reaction phenotyping and in vitro-in vivo extrapolation of drug clearance and drug-drug interaction potential. Drug Metab. Rev. 2010, 42, 196–208. [Google Scholar] [CrossRef]
- Parkinson, A.; Kazmi, F.; Buckley, D.B.; Yerino, P.; Ogilvie, B.W.; Paris, B.L. System-Dependent Outcomes during the Evaluation of Drug Candidates as Inhibitors of Cytochrome P450 (CYP) and Uridine Diphosphate Glucuronosyltransferase (UGT) Enzymes: Human Hepatocytes versus Liver Microsomes versus Recombinant Enzymes. Drug Metab. Pharmacok. 2010, 25, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhang, B.; Parker, R.B.; Laizure, S.C. Clinical implications of genetic variation in carboxylesterase drug metabolism. Expert Opin. Drug Metab. Toxicol. 2018, 14, 131–142. [Google Scholar] [CrossRef]
- Abbas, S.; Greige-Gerges, H.; Karam, N.; Piet, M.H.; Netter, P.; Magdalou, J. Metabolism of Parabens (4-Hydroxybenzoic Acid Esters) by Hepatic Esterases and UDP-Glucuronosyltransferases in Man. Drug Metab. Pharmacok. 2010, 25, 568–577. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Formula | Exact Mass [M−H]− (m/z) | Error (ppm) | tR (min) | Product Ions (m/z) |
|---|---|---|---|---|---|
| Catalposide | C22H26O12 | 481.1349 | −0.6 | 10.26 | 319.0822, 205.0497, 137.0239 |
| M1 | C22H26SO15 | 561.0921 | 0.2 | 8.74 | 481.1349, 319.0822, 205.0499, 137.0239 |
| M2 | C7H6O3 | 137.0239 | −3.6 | 4.07 | 93.0339 |
| M3 | C13H14O9 | 313.0569 | 1.3 | 2.02 | 193.0345, 175.0240, 137.0239, 113.0239, 85.0283 |
| M4 | C28H34O18 | 657.1674 | 0.3 | 6.41 | 481.1353, 319.0823, 205.0499, 175.0240, 137.0239, 113. 0238, 85.0283 |
| Enzyme | Km (μM) | Vmax | Ki (μM) | n | Clint |
|---|---|---|---|---|---|
| Sulfation of catalposide to M1 | |||||
| SULT1A1*1 | 162.0 | 2046.8 | 13.7 | - | 12.6 |
| SULT1A1*2 | 50.5 | 1203.2 | 65.5 | - | 23.8 |
| SULT1C4 | 89.8 | 4576.2 | - | 1.4 | 51.0 |
| SULT1E1 | 456.6 | 4840.0 | 391.4 | - | 10.6 |
| Human liver S9 fractions | 169.7 | 32.8 | - | 1.4 | 0.19 |
| Glucuronidation of catalposide to M4 | |||||
| UGT1A8 | 1230.1 | 48.7 | - | 0.9221 | 0.0396 |
| UGT1A10 | 641.7 | 218.1 | - | 2.3 | 0.3399 |
| Human intestinal microsomes | 1341.3 | 106.2 | - | - | 0.0792 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.-K.; Kim, J.-H.; Shin, Y.; Choi, W.-G.; Kim, S.; Cho, Y.-Y.; Lee, J.Y.; Kang, H.C.; Lee, H.S. Identification of Catalposide Metabolites in Human Liver and Intestinal Preparations and Characterization of the Relevant Sulfotransferase, UDP-glucuronosyltransferase, and Carboxylesterase Enzymes. Pharmaceutics 2019, 11, 355. https://doi.org/10.3390/pharmaceutics11070355
Hwang D-K, Kim J-H, Shin Y, Choi W-G, Kim S, Cho Y-Y, Lee JY, Kang HC, Lee HS. Identification of Catalposide Metabolites in Human Liver and Intestinal Preparations and Characterization of the Relevant Sulfotransferase, UDP-glucuronosyltransferase, and Carboxylesterase Enzymes. Pharmaceutics. 2019; 11(7):355. https://doi.org/10.3390/pharmaceutics11070355
Chicago/Turabian StyleHwang, Deok-Kyu, Ju-Hyun Kim, Yongho Shin, Won-Gu Choi, Sunjoo Kim, Yong-Yeon Cho, Joo Young Lee, Han Chang Kang, and Hye Suk Lee. 2019. "Identification of Catalposide Metabolites in Human Liver and Intestinal Preparations and Characterization of the Relevant Sulfotransferase, UDP-glucuronosyltransferase, and Carboxylesterase Enzymes" Pharmaceutics 11, no. 7: 355. https://doi.org/10.3390/pharmaceutics11070355