Food Bioactive Compounds and Their Interference in Drug Pharmacokinetic/Pharmacodynamic Profiles

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
Methodology of the Review
2. Types of Food-Drug Interactions
- Type I interactions (pharmaceutical), which refer to ex vivo bio-inactivations that usually occur in the delivery device with reactions of hydrolysis, oxidation, neutralization, precipitation, or complexation.
- Type II interactions, which affect the function of an enzyme (subtype A interactions, pharmacokinetic) or a transport mechanism (subtype B interactions, pharmacokinetic) before systemic circulation, in turn altering the absorption and bioavailability of oral or enteral administrations. Complexation, binding, or other deactivations may occur in the gastrointestinal tract (type C interactions, pharmaceutical).
- Type III interactions (pharmacokinetic), which occur after entrance into systemic circulation and involve changes in tissues distribution, penetration, or metabolism.
- Type IV interactions (pharmacokinetic), which denote affections in drug or food component clearance because of influences upon renal or enterohepatic excretion.
3. Pharmaceutical Interactions
4. Pharmacokinetic Interactions
4.1. Pre-Systemic Interactions
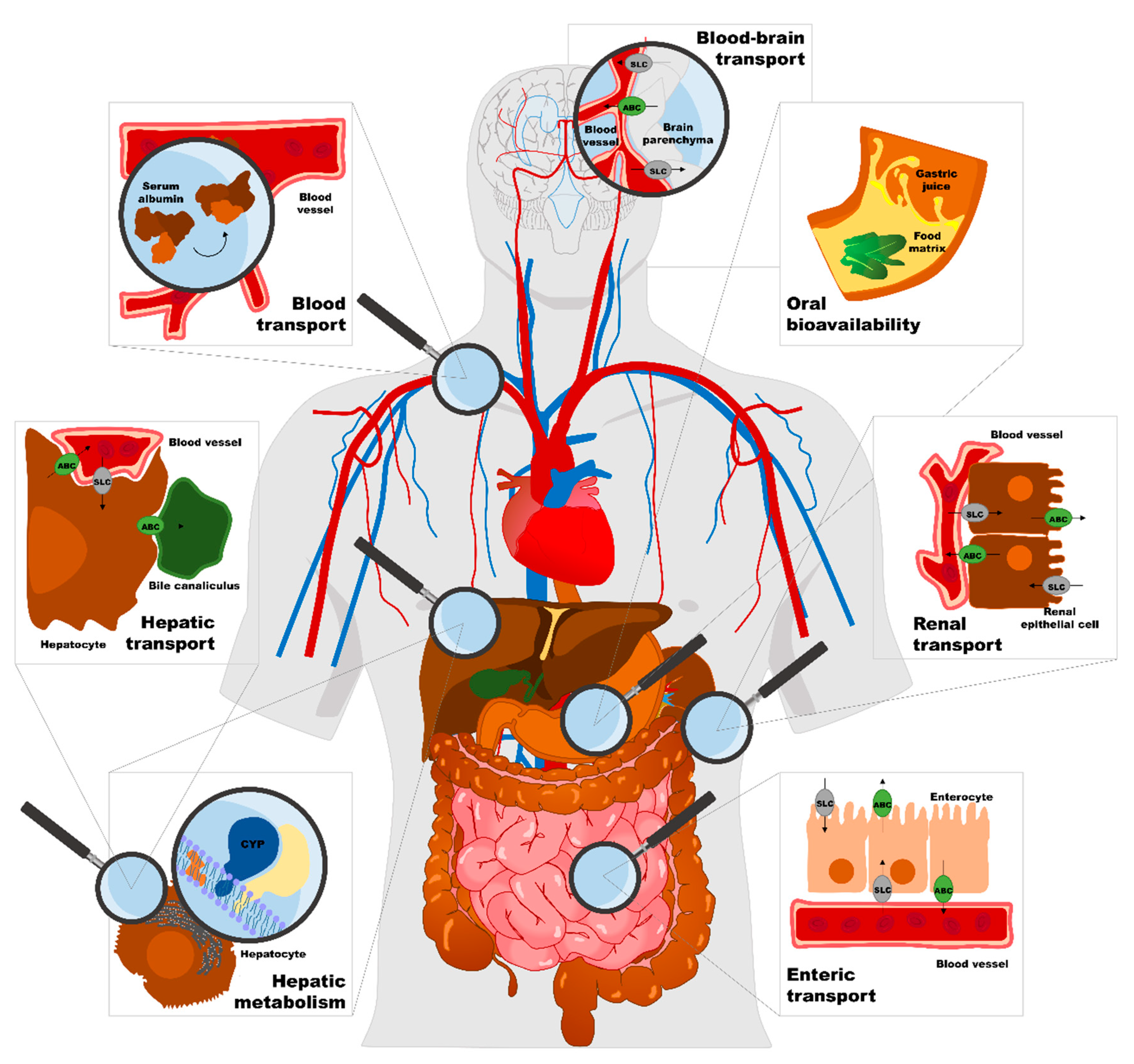
4.1.1. Food Interference with Enteric and Hepatic Transport
4.1.2. Food Interference with Enteric and Hepatic Metabolism
4.2. Post-Systemic Interactions
4.2.1. Food Interference with Blood Transport
4.2.2. Food Interference with Renal and Brain Transport, and Post-Systemic Metabolism
5. Pharmacodynamic Interactions
6. Discussion
7. Conclusions
Funding
Conflicts of Interest
References
- CfDEaR (CDER). Food-Effect Bioavailability and Fed Bioequivalence Studies—Guidance for Industry; Services USDoHaH: Washington, DC, USA, 2002. [Google Scholar]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary Neurotransmitters: A Narrative Review on Current Knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Ko, C.M. Food and Drug Interactions. J. Lifestyle Med. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, W.; Steyn, J.D.; Kotze, A.F.; Hamman, J.H. Beneficial Pharmacokinetic Drug Interactions: A Tool to Improve the Bioavailability of Poorly Permeable Drugs. Pharmaceutics 2018, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Fragoso, L.; Martinez-Arismendi, J.L.; Orozco-Bustos, D.; Reyes-Esparza, J.; Torres, E.; Burchiel, S.W. Potential risks resulting from fruit/vegetable-drug interactions: Effects on drug-metabolizing enzymes and drug transporters. J. Food Sci. 2011, 76, R112–R124. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Brinkhuis, R.F.; van Deemter, L.; Wijnholds, J.; Schinkel, A.H. Extensive contribution of the multidrug transporters P-glycoprotein and Mrp1 to basal drug resistance. Cancer Res. 2000, 60, 5761–5766. [Google Scholar] [PubMed]
- Sconce, E.; Avery, P.; Wynne, H.; Kamali, F. Vitamin K supplementation can improve stability of anticoagulation for patients with unexplained variability in response to warfarin. Blood 2007, 109, 2419–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.A.; Ernst, E. Safety of herbal supplements: A guide for cardiologists. Cardiovasc. Ther. 2010, 28, 246–253. [Google Scholar] [CrossRef]
- Briguglio, M.; Dell’Osso, B.; Galentino, R.; Zanaboni, D.C.; Banfi, G.; Porta, M. Tics and obsessive-compulsive disorder in relation to diet: Two case reports. L’Encephale 2017, 44, 479–481. [Google Scholar] [CrossRef]
- Briguglio, M.; Dina, C.Z.; Servello, D.; Porta, M. Tourette Syndrome and Nutritional Implications. In Proceedings of the 9th Meeting of European Society for the Study of Tourette Syndrome—COST International Conference for Tourette Syndrome, Warsaw, Poland, 8–11 June 2016. [Google Scholar]
- Jensen, K.; Ni, Y.; Panagiotou, G.; Kouskoumvekaki, I. Developing a molecular roadmap of drug-food interactions. PLoS Comput. Biol. 2015, 11, e1004048. [Google Scholar] [CrossRef]
- Ross, A.C.; Caballero, B.; Cousins, R.J.; Tucker, K.L.; Ziegler, T.R. Modern Nutrition in Health and Disease, 11th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014. [Google Scholar]
- Deng, J.; Zhu, X.; Chen, Z.; Fan, C.H.; Kwan, H.S.; Wong, C.H.; Shek, K.Y.; Zuo, Z.; Lam, T.N. A Review of Food-Drug Interactions on Oral Drug Absorption. Drugs 2017, 77, 1833–1855. [Google Scholar] [CrossRef]
- Administration FaD, CfDEaR (CDER). Waiver of In Vivo Bioavailability and Bioequivalence Studies for Immediate-Release Solid Oral Dosage Forms Based on a Biopharmaceutics Classification System—Guidance for Industry; Services USDoHaH: Washington, DC, USA, 2017. [Google Scholar]
- Neuvonen, P.J. Interactions with the absorption of tetracyclines. Drugs 1976, 11, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Polk, R.E. Drug-drug interactions with ciprofloxacin and other fluoroquinolones. Am. J. Med. 1989, 87, 76S–81S. [Google Scholar] [CrossRef]
- Cruz, L.; Castaneda-Hernandez, G.; Navarrete, A. Ingestion of chilli pepper (Capsicum annuum) reduces salicylate bioavailability after oral asprin administration in the rat. Can. J. Physiol. Pharmacol. 1999, 77, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Crevoisier, C.; Zerr, P.; Calvi-Gries, F.; Nilsen, T. Effects of food on the pharmacokinetics of levodopa in a dual-release formulation. Eur. J. Pharm. Biopharm. 2003, 55, 71–76. [Google Scholar] [CrossRef]
- Johansson, O.; Wahlin-Boll, E.; Lindberg, T.; Melander, A. Opposite effects of carbohydrate and protein on phenytoin absorption in man. Drug Nutr. Interact. 1983, 2, 139–144. [Google Scholar] [PubMed]
- Gonzalez Canga, A.; Fernandez Martinez, N.; Sahagun Prieto, A.M.; Garcia Vieitez, J.J.; Diez Liebana, M.J.; Diez Laiz, R.; Sierra Vega, M. Dietary fiber and its interaction with drugs. Nutr. Hosp. 2010, 25, 535–539. [Google Scholar] [PubMed]
- Le Tiec, C.; Barrail, A.; Goujard, C.; Taburet, A.M. Clinical pharmacokinetics and summary of efficacy and tolerability of atazanavir. Clin. Pharmacokinet. 2005, 44, 1035–1050. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.M.; Flood, M.R.; Mancini, R.S.; Whiteman, R.T.; Friedt, G.A.; Kramer, A.R.; Hofstetter, M.A. Oral chemotherapy food and drug interactions: A comprehensive review of the literature. J. Oncol. Pract. 2014, 10, e255–e268. [Google Scholar] [CrossRef] [PubMed]
- Prakash, U.N.; Srinivasan, K. Beneficial influence of dietary spices on the ultrastructure and fluidity of the intestinal brush border in rats. Br. J. Nutr. 2010, 104, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Bressler, R. Grapefruit juice and drug interactions. Exploring mechanisms of this interaction and potential toxicity for certain drugs. Geriatrics 2006, 61, 12–18. [Google Scholar] [PubMed]
- Bailey, D.G.; Dresser, G.K.; Leake, B.F.; Kim, R.B. Naringin is a major and selective clinical inhibitor of organic anion-transporting polypeptide 1A2 (OATP1A2) in grapefruit juice. Clin. Pharmacol. Ther. 2007, 81, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Yamashita, F.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Ohtani, H.; Sawada, Y. Citrus juices inhibit the function of human organic anion-transporting polypeptide OATP-B. Drug Metab. Dispos. 2005, 33, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.V.; Yao, M.; Zhang, Y.; Chong, S. Effect of fruit juices on the oral bioavailability of fexofenadine in rats. J. Pharm. Sci. 2005, 94, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Farkas, D.; Greenblatt, D.J. Influence of fruit juices on drug disposition: Discrepancies between in vitro and clinical studies. Expert Opin. Drug Metab. Toxicol. 2008, 4, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Shirasaka, Y.; Shichiri, M.; Mori, T.; Nakanishi, T.; Tamai, I. Major active components in grapefruit, orange, and apple juices responsible for OATP2B1-mediated drug interactions. J. Pharm. Sci. 2013, 102, 3418–3426. [Google Scholar] [CrossRef] [PubMed]
- Dresser, G.K.; Kim, R.B.; Bailey, D.G. Effect of grapefruit juice volume on the reduction of fexofenadine bioavailability: Possible role of organic anion transporting polypeptides. Clin. Pharmacol. Ther. 2005, 77, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Dresser, G.K.; Bailey, D.G.; Leake, B.F.; Schwarz, U.I.; Dawson, P.A.; Freeman, D.J.; Kim, R.B. Fruit juices inhibit organic anion transporting polypeptide-mediated drug uptake to decrease the oral availability of fexofenadine. Clin. Pharmacol. Ther. 2002, 71, 11–20. [Google Scholar] [CrossRef]
- Mandery, K.; Bujok, K.; Schmidt, I.; Keiser, M.; Siegmund, W.; Balk, B.; König, J.; Fromm, M.F.; Glaeser, H. Influence of the flavonoids apigenin, kaempferol, and quercetin on the function of organic anion transporting polypeptides 1A2 and 2B1. Biochem. Pharmacol. 2010, 80, 1746–1753. [Google Scholar] [CrossRef] [Green Version]
- Mandery, K.; Balk, B.; Bujok, K.; Schmidt, I.; Fromm, M.F.; Glaeser, H. Inhibition of hepatic uptake transporters by flavonoids. Eur. J. Pharm. Sci. 2012, 46, 79–85. [Google Scholar] [CrossRef]
- Ayrton, A.; Morgan, P. Role of transport proteins in drug absorption, distribution and excretion. Xenobiotica 2001, 31, 469–497. [Google Scholar] [CrossRef]
- Honda, Y.; Ushigome, F.; Koyabu, N.; Morimoto, S.; Shoyama, Y.; Uchiumi, T.; Kuwano, M.; Ohtani, H.; Sawada, Y. Effects of grapefruit juice and orange juice components on P-glycoprotein- and MRP2-mediated drug efflux. Br. J. Pharmacol. 2004, 143, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, T.; Satsu, H.; Hatsugai, Y.; Aizawa, K.; Inakuma, T.; Nagata, S.; Sakuda, S.H.; Nagasawa, H.; Shimizu, M. Inhibitory effect of a bitter melon extract on the P-glycoprotein activity in intestinal Caco-2 cells. Br. J. Pharmacol. 2004, 143, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, H.H.; Lee, M.; Chung, H.J.; Lee, S.K.; Kim, D.H. Effects of diosmin, a flavonoid glycoside in citrus fruits, on P-glycoprotein-mediated drug efflux in human intestinal Caco-2 cells. J. Agric. Food Chem. 2007, 55, 7620–7625. [Google Scholar] [CrossRef]
- Takanaga, H.; Ohnishi, A.; Yamada, S.; Matsuo, H.; Morimoto, S.; Shoyama, Y.; Ohtani, H.; Sawada, Y. Polymethoxylated flavones in orange juice are inhibitors of P-glycoprotein but not cytochrome P450 3A4. J. Pharmacol. Exp. Ther. 2000, 293, 230–236. [Google Scholar] [PubMed]
- Fleisher, B.; Unum, J.; Shao, J.; An, G. Ingredients in fruit juices interact with dasatinib through inhibition of BCRP: A new mechanism of beverage-drug interaction. J. Pharm. Sci. 2015, 104, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, T.; Yamaki, T.; Kitagawa, S. Effects of chemopreventive citrus phytochemicals on human P-glycoprotein and multidrug resistance protein 1. Eur. J. Pharmacol. 2008, 600, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.E.; Jeffery, E.H. Sulforaphane and erucin increase MRP1 and MRP2 in human carcinoma cell lines. J. Nutr. Biochem. 2008, 19, 246–254. [Google Scholar] [CrossRef]
- Zhang, R.; Jie, J.; Zhou, Y.; Cao, Z.; Li, W. Long-term effects of Panax ginseng on disposition of fexofenadine in rats in vivo. Am. J. Chin. Med. 2009, 37, 657–667. [Google Scholar] [CrossRef]
- Nabekura, T.; Yamaki, T.; Ueno, K.; Kitagawa, S. Inhibition of P-glycoprotein and multidrug resistance protein 1 by dietary phytochemicals. Cancer Chemother. Pharmacol. 2008, 62, 867–873. [Google Scholar] [CrossRef]
- Fan, L.; Tao, G.Y.; Wang, G.; Chen, Y.; Zhang, W.; He, Y.J.; Li, Q.; Lei, H.P.; Jiang, F.; Hu, D.L.; et al. Effects of Ginkgo biloba extract ingestion on the pharmacokinetics of talinolol in healthy Chinese volunteers. Ann. Pharmacother. 2009, 43, 944–949. [Google Scholar] [CrossRef]
- Bhardwaj, R.K.; Glaeser, H.; Becquemont, L.; Klotz, U.; Gupta, S.K.; Fromm, M.F. Piperine, a major constituent of black pepper, inhibits human P-glycoprotein and CYP3A4. J. Pharmacol. Exp. Ther. 2002, 302, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, T.; Yamaki, T.; Hiroi, T.; Ueno, K.; Kitagawa, S. Inhibition of anticancer drug efflux transporter P-glycoprotein by rosemary phytochemicals. Pharmacol. Res. 2010, 61, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.P.; Hsieh, Y.W.; Lin, S.P.; Chi, Y.C.; Hariharan, P.; Chao, P.D.; Hou, Y.C. Potential modulation on P-glycoprotein and CYP3A by soymilk and miso: In vivo and ex-vivo studies. Food Chem. 2014, 149, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Yoon, Y.J.; Shon, J.H.; Cha, I.J.; Shin, J.G.; Liu, K.H. Inhibitory effects of fruit juices on CYP3A activity. Drug Metab. Dispos. 2006, 34, 521–523. [Google Scholar] [CrossRef] [PubMed]
- De Castro, W.V.; Mertens-Talcott, S.; Rubner, A.; Butterweck, V.; Derendorf, H. Variation of flavonoids and furanocoumarins in grapefruit juices: A potential source of variability in grapefruit juice-drug interaction studies. J. Agric. Food Chem. 2006, 54, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiao, P.; Ou-Yang, D.S.; Fan, L.; Guo, D.; Wang, Y.N.; Han, Y.; Tu, J.H.; Zhou, G.; Huang, Y.F.; et al. Simultaneous action of the flavonoid quercetin on cytochrome P450 (CYP) 1A2, CYP2A6, N-acetyltransferase and xanthine oxidase activity in healthy volunteers. Clin. Exp. Pharmacol. Physiol. 2009, 36, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.R.; Chen, J.; Freiburghaus, A.U.; Follath, F. Metabolism of theophylline by cDNA-expressed human cytochromes P-450. Br. J. Clin. Pharmacol. 1995, 39, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Bencsik, T.; Boda, G.; Fliszar-Nyul, E.; Lemli, B.; Kunsagi-Mate, S.; Poór, M. Interactions of casticin, ipriflavone, and resveratrol with serum albumin and their inhibitory effects on CYP2C9 and CYP3A4 enzymes. Biomed. Pharmacother. 2018, 107, 777–784. [Google Scholar] [CrossRef]
- Piver, B.; Berthou, F.; Dreano, Y.; Lucas, D. Inhibition of CYP3A, CYP1A and CYP2E1 activities by resveratrol and other non volatile red wine components. Toxicol. Lett. 2001, 125, 83–91. [Google Scholar] [CrossRef]
- Srinivas, N.R. Cranberry juice ingestion and clinical drug-drug interaction potentials; review of case studies and perspectives. J. Pharm. Pharm. Sci. 2013, 16, 289–303. [Google Scholar] [CrossRef]
- Ansell, J.; McDonough, M.; Zhao, Y.; Harmatz, J.S.; Greenblatt, D.J. The absence of an interaction between warfarin and cranberry juice: A randomized, double-blind trial. J. Clin. Pharmacol. 2009, 49, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, M.; Okumura, M.; Fujita, K.; Ogikubo, T.; Yamasaki, K.; Iwakiri, T.; Setoguchi, N.; Arimori, K. Effects of pomegranate juice on human cytochrome p450 3A (CYP3A) and carbamazepine pharmacokinetics in rats. Drug Metab. Dispos. 2005, 33, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Hidaka, M.; Sekiya, H.; Kawano, Y.; Yamasaki, K.; Okumura, M.; Arimori, K. Effects of pomegranate juice on human cytochrome P450 2C9 and tolbutamide pharmacokinetics in rats. Drug Metab. Dispos. 2007, 35, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, M.T.; White, R.E.; Yang, C.S. Effects of bioflavonoids on hepatic P450 activities. Xenobiotica 1995, 25, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Backman, J.T.; Maenpaa, J.; Belle, D.J.; Wrighton, S.A.; Kivisto, K.T.; Neuvonen, P.J. Lack of correlation between in vitro and in vivo studies on the effects of tangeretin and tangerine juice on midazolam hydroxylation. Clin. Pharmacol. Ther. 2000, 67, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.; Will, F.; Dietrich, H.; Schrenk, D. Cytochrome P450 1A1 expression and activity in Caco-2 cells: Modulation by apple juice extract and certain apple polyphenols. J. Agric. Food Chem. 2006, 54, 10262–10268. [Google Scholar] [CrossRef]
- Anwar-Mohamed, A.; El-Kadi, A.O. Sulforaphane induces CYP1A1 mRNA, protein, and catalytic activity levels via an AhR-dependent pathway in murine hepatoma Hepa 1c1c7 and human HepG2 cells. Cancer Lett. 2009, 275, 93–101. [Google Scholar] [CrossRef]
- Fimognari, C.; Lenzi, M.; Hrelia, P. Interaction of the isothiocyanate sulforaphane with drug disposition and metabolism: Pharmacological and toxicological implications. Curr. Drug Metab. 2008, 9, 668–678. [Google Scholar] [CrossRef]
- Kot, M.; Daniel, W.A. Effect of cytochrome P450 (CYP) inducers on caffeine metabolism in the rat. Pharmacol. Rep. 2007, 59, 296–305. [Google Scholar]
- Zhou, S.; Gao, Y.; Jiang, W.; Huang, M.; Xu, A.; Paxton, J.W. Interactions of herbs with cytochrome P450. Drug Metab. Rev. 2003, 35, 35–98. [Google Scholar] [CrossRef]
- Paolini, M.; Barillari, J.; Broccoli, M.; Pozzetti, L.; Perocco, P.; Cantelli-Forti, G. Effect of liquorice and glycyrrhizin on rat liver carcinogen metabolizing enzymes. Cancer Lett. 1999, 145, 35–42. [Google Scholar] [CrossRef]
- Veeramachaneni, S.; Ausman, L.M.; Choi, S.W.; Russell, R.M.; Wang, X.D. High dose lycopene supplementation increases hepatic cytochrome P4502E1 protein and inflammation in alcohol-fed rats. J. Nutr. 2008, 138, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hamilton, S.M.; Stewart, C.; Strother, A.; Teel, R.W. Inhibition of liver microsomal cytochrome P450 activity and metabolism of the tobacco-specific nitrosamine NNK by capsaicin and ellagic acid. Anticancer Res. 1993, 13, 2341–2346. [Google Scholar] [PubMed]
- Oikawa, S.; Nagao, E.; Sakano, K.; Kawanishi, S. Mechanism of oxidative DNA damage induced by capsaicin, a principal ingredient of hot chili pepper. Free Radic. Res. 2006, 40, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Doostdar, H.; Burke, M.D.; Mayer, R.T. Bioflavonoids: Selective substrates and inhibitors for cytochrome P450 CYP1A and CYP1B1. Toxicology 2000, 144, 31–38. [Google Scholar] [CrossRef]
- Fontana, R.J.; Lown, K.S.; Paine, M.F.; Fortlage, L.; Santella, R.M.; Felton, J.S.; Knize, M.G.; Greenberg, A.; Watkins, P.B. Effects of a chargrilled meat diet on expression of CYP3A, CYP1A, and P-glycoprotein levels in healthy volunteers. Gastroenterology 1999, 117, 89–98. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, C.Q.; He, Y.J.; Chen, B.L.; Wang, G.; Zhou, G.; Zhang, W.; Tan, Z.R.; Cao, S.; Wang, L.P.; et al. Genistein alters caffeine exposure in healthy female volunteers. Eur. J. Clin. Pharmacol. 2011, 67, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, I.; Desager, J.P.; Horsmans, Y. Inhibition of chlorzoxazone metabolism, a clinical probe for CYP2E1, by a single ingestion of watercress. Clin. Pharmacol. Ther. 1998, 64, 144–149. [Google Scholar] [CrossRef]
- Lampe, J.W.; King, I.B.; Li, S.; Grate, M.T.; Barale, K.V.; Chen, C.; Feng, Z.; Potter, J.D. Brassica vegetables increase and apiaceous vegetables decrease cytochrome P450 1A2 activity in humans: Changes in caffeine metabolite ratios in response to controlled vegetable diets. Carcinogenesis 2000, 21, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Bedada, S.K.; Yellu, N.R.; Neerati, P. Effect of Resveratrol Treatment on the Pharmacokinetics of Diclofenac in Healthy Human Volunteers. Phytother. Res. 2016, 30, 397–401. [Google Scholar] [CrossRef]
- Bedada, S.K.; Neerati, P. Resveratrol Pretreatment Affects CYP2E1 Activity of Chlorzoxazone in Healthy Human Volunteers. Phytother. Res. 2016, 30, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Lhoste, E.F.; Gloux, K.; De Waziers, I.; Garrido, S.; Lory, S.; Philippe, C.; Rabot, S.A.; Knasmüller, S. The activities of several detoxication enzymes are differentially induced by juices of garden cress, water cress and mustard in human HepG2 cells. Chem. Biol. Interact. 2004, 150, 211–219. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Frye, R.F. Inhibitory effects of commonly used herbal extracts on UDP-glucuronosyltransferase 1A4, 1A6, and 1A9 enzyme activities. Drug Metab. Dispos. 2011, 39, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Saruwatari, A.; Okamura, S.; Nakajima, Y.; Narukawa, Y.; Takeda, T.; Tamura, H. Pomegranate juice inhibits sulfoconjugation in Caco-2 human colon carcinoma cells. J. Med. Food 2008, 11, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Talalay, P. Antioxidant functions of sulforaphane: A potent inducer of Phase II detoxication enzymes. Food Chem. Toxicol. 1999, 37, 973–979. [Google Scholar] [CrossRef]
- Yeh, C.T.; Yen, G.C. Effect of vegetables on human phenolsulfotransferases in relation to their antioxidant activity and total phenolics. Free Radic. Res. 2005, 39, 893–904. [Google Scholar] [CrossRef]
- Bradfield, C.A.; Chang, Y.; Bjeldanes, L.F. Effects of commonly consumed vegetables on hepatic xenobiotic-metabolizing enzymes in the mouse. Food Chem. Toxicol. 1985, 23, 899–904. [Google Scholar] [CrossRef]
- Heldt, T.; Loss, S.H. Drug-nutrient interactions in the intensive care unit: Literature review and current recommendations. Rev. Bras. Ter. Intensiv. 2013, 25, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Qi, W.; Huang, R.; Su, R.; He, Z. Elucidating the influence of gold nanoparticles on the binding of salvianolic acid B and rosmarinic acid to bovine serum albumin. PLoS ONE 2015, 10, e0118274. [Google Scholar] [CrossRef]
- Shao, X.; Ai, N.; Xu, D.; Fan, X. Exploring the interaction between Salvia miltiorrhiza and human serum albumin: Insights from herb-drug interaction reports, computational analysis and experimental studies. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 161, 1–7. [Google Scholar] [CrossRef]
- Latruffe, N.; Menzel, M.; Delmas, D.; Buchet, R.; Lancon, A. Compared binding properties between resveratrol and other polyphenols to plasmatic albumin: Consequences for the health protecting effect of dietary plant microcomponents. Molecules 2014, 19, 17066–17077. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, P.; Kanakis, C.D.; Tarantilis, P.; Pollissiou, M.G.; Tajmir-Riahi, H.A. Resveratrol, genistein, and curcumin bind bovine serum albumin. J. Phys. Chem. B 2010, 114, 3348–3354. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, B.; Sengupta, P.K. Binding of quercetin with human serum albumin: A critical spectroscopic study. Biopolymers 2003, 72, 427–434. [Google Scholar] [CrossRef]
- Wang, L.; Sweet, D.H. Renal organic anion transporters (SLC22 family): Expression, regulation, roles in toxicity, and impact on injury and disease. AAPS J. 2013, 15, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, X.; Roberts, J.; Janosky, B.; Lin, M.H. Differential role of P-glycoprotein and breast cancer resistance protein in drug distribution into brain, CSF and peripheral nerve tissues in rats. Xenobiotica 2015, 45, 547–555. [Google Scholar] [CrossRef]
- Cao, J.J.; Johnson, L.K.; Hunt, J.R. A diet high in meat protein and potential renal acid load increases fractional calcium absorption and urinary calcium excretion without affecting markers of bone resorption or formation in postmenopausal women. J. Nutr. 2011, 141, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sweet, D.H. Interaction of Natural Dietary and Herbal Anionic Compounds and Flavonoids with Human Organic Anion Transporters 1 (SLC22A6), 3 (SLC22A8), and 4 (SLC22A11). Evid. Based Complement. Alternat. Med. 2013, 2013, 612527. [Google Scholar] [CrossRef]
- Moons, T.; de Roo, M.; Claes, S.; Dom, G. Relationship between P-glycoprotein and second-generation antipsychotics. Pharmacogenomics 2011, 12, 1193–1211. [Google Scholar] [CrossRef]
- Telang, U.; Ji, Y.; Morris, M.E. ABC transporters and isothiocyanates: Potential for pharmacokinetic diet-drug interactions. Biopharm. Drug Dispos. 2009, 30, 335–344. [Google Scholar] [CrossRef]
- Wang, H.; Leung, L.K. The carotenoid lycopene differentially regulates phase I and II enzymes in dimethylbenz[a]anthracene-induced MCF-7 cells. Nutrition 2010, 26, 1181–1187. [Google Scholar] [CrossRef]
- Yellapu, R.K.; Mittal, V.; Grewal, P.; Fiel, M.; Schiano, T. Acute liver failure caused by ‘fat burners’ and dietary supplements: A case report and literature review. Can. J. Gastroenterol. 2011, 25, 157–160. [Google Scholar] [PubMed]
- Yanagida, K.; Hla, T. A dark side to omega-3 fatty acids. Nature 2017, 552, 180–181. [Google Scholar] [CrossRef] [PubMed]
- Kompoliti, K.; Fan, W.; Leurgans, S. Complementary and alternative medicine use in Gilles de la Tourette syndrome. Mov. Disord. 2009, 24, 2015–2019. [Google Scholar] [CrossRef] [PubMed]
- Levy, I.; Attias, S.; Ben-Arye, E.; Goldstein, L.; Matter, I.; Somri, M.; Schiff, E. Perioperative Risks of Dietary and Herbal Supplements. World J. Surg. 2017, 41, 927–934. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Tinz, J. Multivitamin/mineral supplements: Rationale and safety—A systematic review. Nutrition 2016, 33, 76–82. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef]
- Silveira, C.M.; Moreira, A.V.; Martino, H.S.; Gomide, R.S.; Pinheiro, S.S.; Della Lucia, C.M.; Pinheiro-Sant’ana, H.M. Effect of cooking methods on the stability of thiamin and folic acid in fortified rice. Int. J. Food Sci. Nutr. 2016, 68, 179–187. [Google Scholar] [CrossRef]
- Barichella, M.; Cereda, E.; Cassani, E.; Pinelli, G.; Iorio, L.; Ferri, V.; Privitera, G.; Pasqua, M.; Valentino, A.; Monajemi, F.; et al. Dietary habits and neurological features of Parkinson’s disease patients: Implications for practice. Clin. Nutr. 2017, 36, 1054–1061. [Google Scholar] [CrossRef]
- Tomita, T.; Goto, H.; Sumiya, K.; Yoshida, T.; Tanaka, K.; Kohda, Y. Stabilization of the Serum Lithium Concentration by Regulation of Sodium Chloride Intake: Case Report. Yakugaku Zasshi 2016, 136, 517–521. [Google Scholar] [CrossRef] [Green Version]
- Flockhart, D.A. Dietary restrictions and drug interactions with monoamine oxidase inhibitors: An update. J. Clin. Psychiatry 2012, 73 (Suppl. 1), 17–24. [Google Scholar] [CrossRef]
- Bailey, D.G.; Dresser, G.K.; Bend, J.R. Bergamottin, lime juice, and red wine as inhibitors of cytochrome P450 3A4 activity: Comparison with grapefruit juice. Clin. Pharmacol. Ther. 2003, 73, 529–537. [Google Scholar] [CrossRef]
- Benet, L.Z.; Hoener, B.A. Changes in plasma protein binding have little clinical relevance. Clin. Pharmacol. Ther. 2002, 71, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zeng, M.; Sun, R.; Hu, M. Disposition of flavonoids impacts their efficacy and safety. Curr. Drug Metab. 2014, 15, 841–864. [Google Scholar] [CrossRef] [PubMed]
- Actis-Goretta, L.; Leveques, A.; Rein, M.; Teml, A.; Schafer, C.; Hofmann, U.; Li, H.; Schwab, M.; Eichelbaum, M.; Williamson, G. Intestinal absorption, metabolism, and excretion of (−)-epicatechin in healthy humans assessed by using an intestinal perfusion technique. Am. J. Clin. Nutr. 2013, 98, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.C.; Saville, D.J.; Coville, P.F.; Wanwimolruk, S. Content of CYP3A4 inhibitors, naringin, naringenin and bergapten in grapefruit and grapefruit juice products. Pharm. Acta Helv. 2000, 74, 379–385. [Google Scholar] [CrossRef]
- National Consumers League USFaDA. Avoid Food-Drug Interactions—A Guide; Services USDoHaH: Washington, DC, USA, 2013. [Google Scholar]
- Bouraoui, A.; Brazier, J.L.; Zouaghi, H.; Rousseau, M. Theophylline pharmacokinetics and metabolism in rabbits following single and repeated administration of Capsicum fruit. Eur. J. Drug Metab. Pharmacokinet. 1995, 20, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.I.; Real, R.; Perez, M.; Mendoza, G.; Prieto, J.G.; Merino, G. Modulation of the activity of ABC transporters (P-glycoprotein, MRP2, BCRP) by flavonoids and drug response. J. Pharm. Sci. 2010, 99, 598–617. [Google Scholar] [CrossRef]


{kind=link}
| Transport Enzyme | Food Bioactive Compound | Reference |
|---|---|---|
| OATP1A2 | - grapefruit compounds | - [24,27,28,30,31] |
| - naringin, hesperidin | - [25] | |
| - apigenin, kaempferol, quercetin | - [32] | |
| OATP1B1 | - naringenin, naringin, quercetin, daidzein, genistein, glycyrrhizin, caffeine | - [11] |
| OATP1B3 | - apigenin, kaempferol, quercetin, naringenin, naringin, rutin | - [33] |
| - naringenin, naringin, quercetin, daidzein, genistein, glycyrrhizin, caffeine | - [11] | |
| OATP2B1 | - grapefruit, orange, and apple compounds | - [26,27,28,29,31] |
| - apigenin, kaempferol, quercetin, naringenin, naringin, rutin | - [33] | |
| - naringenin, naringin, quercetin, daidzein, genistein, glycyrrhizin, caffeine | - [11] | |
| OCT1 | - caffeine | - [11] |
| Transport enzyme | Food bioactive compound | Reference |
|---|---|---|
| MDR1 | - grapefruit, orange, tangerine, bitter melon compounds, diosmin, nobiletin | - [35,36,37,38,40] |
| - Welsh onion compounds | - [36] | |
| - Ginseng compounds | - [42] | |
| - sesamin, ginkgolic acid, glycyrrhetinic acid, glabridin | - [43,44] | |
| - piperine | - [45] | |
| - carnosic acid | - [46] | |
| - soybean, dokudami, soymilk, and miso | - [36,47] | |
| - resveratrol | - [11] | |
| MRP1 | - nobiletin | - [40] |
| - sulforaphane | - [41] | |
| - glycyrrhetinic acid | - [43] | |
| MRP2 | - grapefruit and orange compounds, naringenin sulforaphane | - [11,35] |
| - ginkgo compounds | - [41] | |
| - apigenin, biochanin A, genistein, glycyrrhizin, | - [44] | |
| - quercetin, resveratrol | - [11] | |
| BCRP | - bergamottin, tangeretin, nobiletin | - [39] |
| Metabolizing Enzyme | Food bioactive compound | Reference |
|---|---|---|
| CYP1A | - apigenin, apple, caffeine, capsaicin, charcoal-broiled meat, diallyl disulphide, ellagic acid, epicatechin gallate, esperetin, galangin, genistein, grapefruit, hesperetin, isorhamnetin, kaempferol, liquorice compounds, luteolin, naringenin, pinocembrin, piperine, quercetin, sulforaphane, tangeretin, teophillyne | - [11,24,50,51,58,60,61,63,64,65,68,69,70,71,73] |
| CYP1B | - hesperetin | - [69] |
| CYP2A | - bergapten, capsaicin, ellagic acid, furanocoumarin, genistein, quercetin, xanthotoxin | - [11,50,67,71] |
| CYP2B | - bergapten, capsaicin, diallyl disulphide, ellagic acid, furanocoumarin, piperine, xanthotoxin | - [11,64,67] |
| CYP2C | - apigenin, bergapten, biochanin A, caffeic acid, caffeine, capsaicin, casticin, cranberry, curcumin, daidzein, ellagic acid, galangin, genistein, hesperetin, kaempferol, luteolin, myricetin, naringenin, nicotine, pinocembrin, quercetin, resveratrol, sesamin, xanthotoxin. | - [11,52,54,63,67,74] |
| CYP2D | - 6-shogaol, apigenin, bergapten, β-sitosterol, capsaicin, curcumin, daidzein, formononetin, galangin, harmine, hesperetin, kaempferol, luteolin, myricetin, pinocembrin, quercetin, resveratrol, serotonin, sesamin, xanthotoxin | - [11] |
| CYP2E | - diallyl disulphide, ethanol, lycopene, piperine, resveratrol, watercress compounds | - [51,53,64,66,72,75] |
| CYP3A | - 6-shogaol, apigenin, bergamottin, bergapten, biochanin A, black mulberry, caffeic acid, caffeine, capsaicin, casticin, curcumin, daidzein, diallyl disulphide, diosgenin, ellagic acid, estragole, galangin, epicatechin gallate, genistein, geraniol, glycyrrhizin, harmine, hesperetin, imperatorin, isopimpinellin, kaempferol, luteolin, myricetin, naringenin, naringin, piperine, pomegranate, pyrocatechol, quercetin, resveratrol, rutoside, safrole, sesamin, sulforaphane, tangeretin, taxifolin, theobromine, xanthotoxin | - [4,11,24,28,45,47,48,49,52,54,56,57,62,63,64,67] |
| Metabolizing Enzyme | Food/Bioactive Compound | Reference |
|---|---|---|
| SULT | - punicalagin | - [78] |
| - sulforaphane, asparagus, eggplant, cauliflower, celery,carrot, and potato compounds | - [80] | |
| UGT | - garden cress and mustard | - [76] |
| - milk thistle, epicatechin gallate, saw palmetto,cranberry, quercetin, kaempferol | - [77] | |
| - (+)-catechin, apigenin, ergosterol, galangin, menthol, naringenin, carvacrol diosgenin, eugenol linalool naringin quercetin, tea polyphenol, borneol citronellol ethinyl estradiol shikimic acid, caproic acid | - [11] | |
| DT-diaphorase | - watercress, garden cress, mustard | - [76] |
| - sulforaphane | - [79] | |
| EPH | - soya bean, Brussels sprouts, cauliflower, and onion | - [81] |
| GST | - sulforaphane | - [79] |
| - soya bean, Brussels sprouts, cauliflower, onion | - [81] | |
| NAT | - quercetin | - [50] |
| - mustard | - [76] |
| Types of Food-Drug Interactions | Targets | Food Components |
|---|---|---|
| a. Pharmaceutical | ||
| - solubilization, stabilization, and dissolution properties | - alcohol (wine, beer, spirits), capsaicin, cations from milk and dairy products, insoluble fibers (cellulose, bran), soluble fibers (pectin, glucomannan, psyllium, guar gum), lipids, phytoserotonin, proteins | |
| b. Pharmacokinetic | ||
| Pre-systemic | - enteric and hepatic transporters of SLC and ABC | - 1-monopalmitin, apigenin, bergamottin, biochanin A, caffeine, carnosic acid, daidzein, genistein, ginkgolic acid, glabridin, glycyrrhetinic acid, glycyrrhizin, hesperidin, kaempferol, naringenin, naringin, nobiletin, piperine, quercetin, resveratrol, rutin, sesamin, soymilk and miso components, sulforaphane, tangeretin, tangeretin |
| - enteric and hepatic metabolizing enzymes of CYPP450 families and phase II enzyme reactions, such as SULT, UGT, DT-diaphorase, EPH, GST, NAT | - (+)-catechin, 6-shogaol, apigenin, asparagus components, bergamottin, bergapten, biochanin A, black mulberry components, borneol, caffeic acid, caffeine, caproic acid, capsaicin,, carrot components, carvacrol, casticin, charcoal-broiled meat, citronellol, cranberry components, curcumin, daidzein, diallyl disulphide, dihydrocapsaicin, diosgenin, eggplant components, ellagic acid, epicatechin gallate, ergosterol, estragole, ethanol, ethinyl estradiol, eugenol, formononetin, freeze-dried soya bean components, furanocoumarin, furocoumarin, galangin, genistein, geraniol, glycyrrhizin, harmine, hesperetin, imperatorin, isopimpinellin, isorhamnetin, kaempferol, linalool, luteolin,, lycopene, menthol, milk thistle components, mustard, myricetin, naringenin, naringin, nicotine, onion components, phytoserotonin, pinocembrin, piperine, pomegranate components, potato components, punicalagin, pyrocatechol, quercetin, resveratrol, rutoside, safrole, saw palmetto components, sesamin, shikimic acid, soymilk and miso components, sulforaphane, tangeretin, taxifolin, tea polyphenols, theobromine, theophylline, watercress and garden cress components, xanthotoxin, β-sitosterol | |
| Post-systemic | - transporters, such as albumin, SLC, and ABC. Metabolizing enzymes of CYPP450 families and phase II enzyme reactions, such as UGT | - apigenin, caffeine, casticin, curcumin, genistein, ginseng, isothiocyanates, licorice, lycopene, naringenin, nicotinic acid, proteins, quercetin, resveratrol, resveratrol, rosmarinic acid, salvianolic acid B |
| c. Pharmacodynamic | ||
| - aromatase, epidermal growth factor, 5-hydroxytryptamine, carbonic anhydrase, estrogen, prostaglandin G/H synthase 2, acetylcholinesterase, cholinesterase, glutamate receptors, D(2) and D(1A), and β2 adrenergic receptors | - α-aminoadipic acid, apigenin, caffeic acid, daidzein, dihydrocapsaicin, eriodictyol, galangin, genistein, glutamate/glutamic acid, harmine, hesperetin, higenamine, kaempferol, liquiritin, luteolin, myricetin, naringenin, P-hydroxybenzoic acid, dopamine, phytoserotonin, quercetin, resveratrol, rutoside, taxifolin |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briguglio, M.; Hrelia, S.; Malaguti, M.; Serpe, L.; Canaparo, R.; Dell’Osso, B.; Galentino, R.; De Michele, S.; Zanaboni Dina, C.; Porta, M.; et al. Food Bioactive Compounds and Their Interference in Drug Pharmacokinetic/Pharmacodynamic Profiles. Pharmaceutics 2018, 10, 277. https://doi.org/10.3390/pharmaceutics10040277
Briguglio M, Hrelia S, Malaguti M, Serpe L, Canaparo R, Dell’Osso B, Galentino R, De Michele S, Zanaboni Dina C, Porta M, et al. Food Bioactive Compounds and Their Interference in Drug Pharmacokinetic/Pharmacodynamic Profiles. Pharmaceutics. 2018; 10(4):277. https://doi.org/10.3390/pharmaceutics10040277
Chicago/Turabian StyleBriguglio, Matteo, Silvana Hrelia, Marco Malaguti, Loredana Serpe, Roberto Canaparo, Bernardo Dell’Osso, Roberta Galentino, Sara De Michele, Carlotta Zanaboni Dina, Mauro Porta, and et al. 2018. "Food Bioactive Compounds and Their Interference in Drug Pharmacokinetic/Pharmacodynamic Profiles" Pharmaceutics 10, no. 4: 277. https://doi.org/10.3390/pharmaceutics10040277