
Low Replication Efficiency of a Japanese Rabbit Hepatitis E Virus Strain in the Human Hepatocarcinoma Cell Line PLC/PRF/5
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rabbit HEV Strains and Preparation of the Virus Suspensions
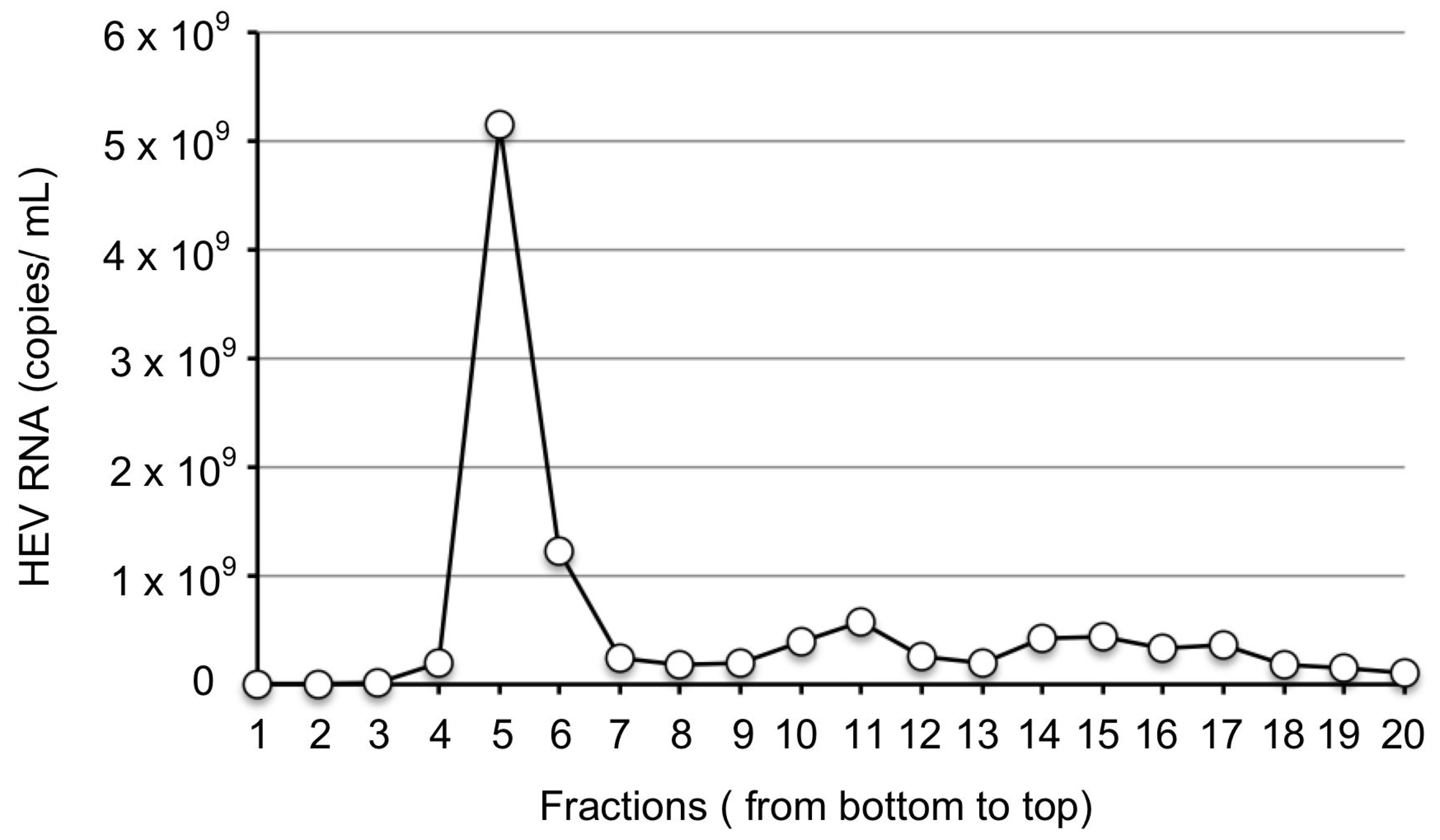
2.2. Purification of JP-59 from Stool Suspension
2.3. Cell Culture and Virus Infection
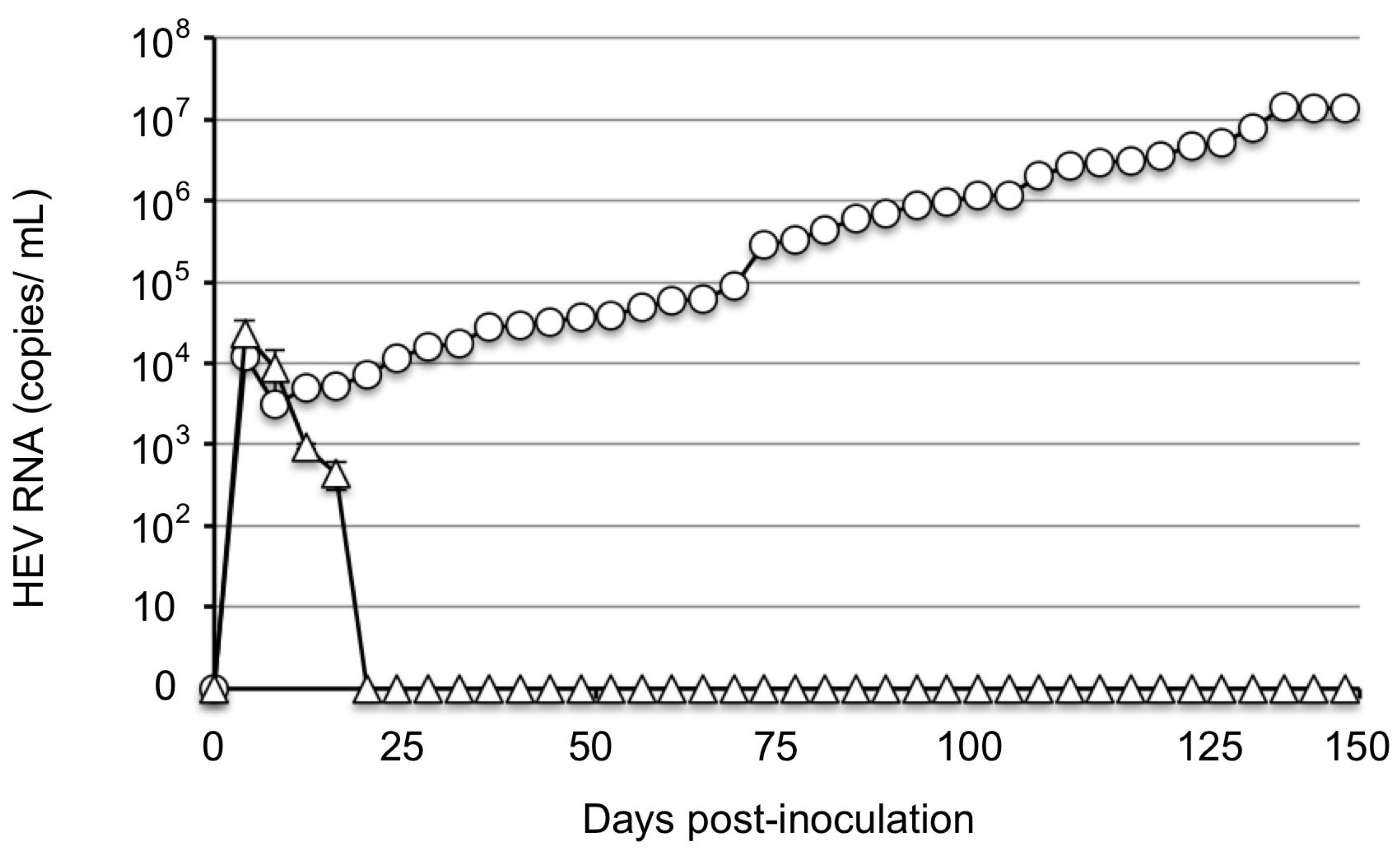
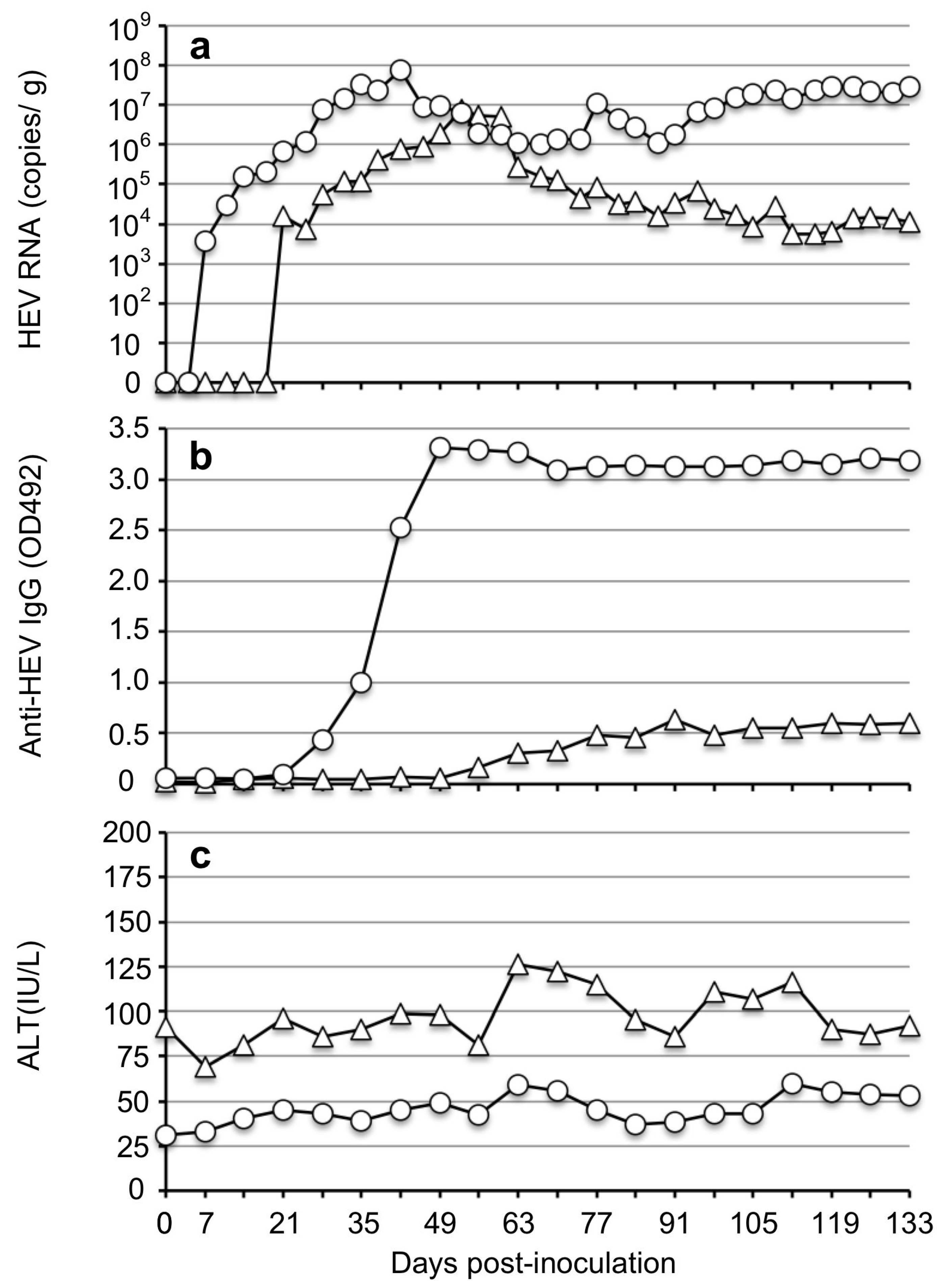
2.4. Inoculation of Rabbits and Sample Collection
2.5. Detection of Rabbit HEV RNA
2.6. Detection of Rabbit Anti-HEV IgG Antibodies
2.7. Liver Enzyme Level
2.8. Next-Generation Sequencing (NGS) for the Entire JP-59 Genome
3. Results
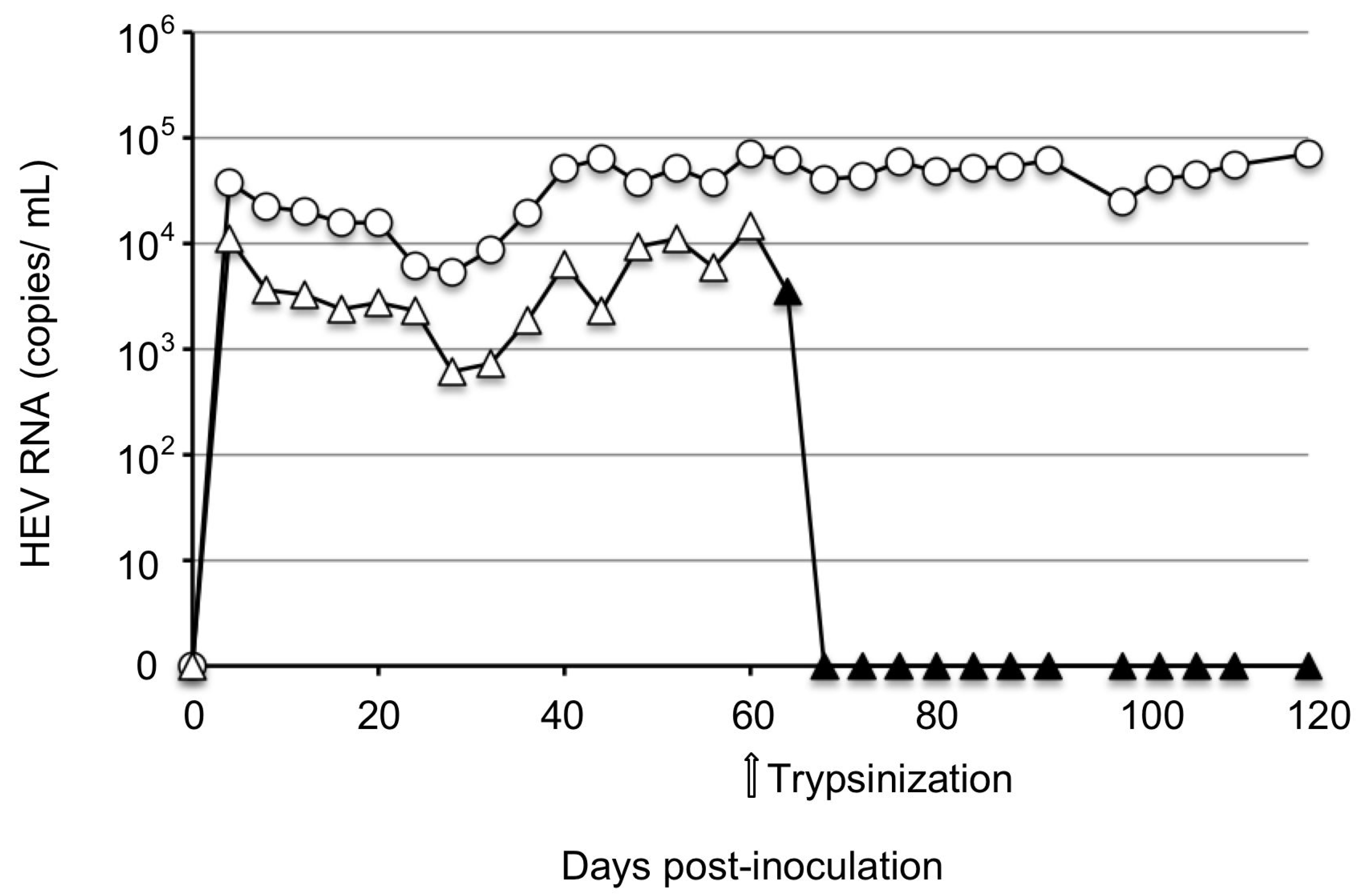
3.1. No Virus Was Recovered from PLC/PRF/5 Cells Inoculated with JP-59s
3.2. JP-59 Replicates in PLC/PRF/5 Cells When a Purified High-Titer Virus Was Used
3.3. Infectivity of JP59c
3.4. Nucleotide Sequence Analyses of JP-59 Strains
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purdy, M.A.; Drexler, J.F.; Meng, X.-J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV virus taxonomy profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef]
- Meng, X.J. Hepatitis E virus: Animal reservoirs and zoonotic risk. Vet. Microbiol. 2010, 140, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.-H.; Tan, B.-H.; Teo, E.C.-Y.; Lim, S.-G.; Dan, Y.-Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.; Tan, C.-K.; et al. Chronic infection with camelid hepatitis E virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.; Chan, J.F.; Zhao, P.S.; Chan, W.; Poon, R.W.; Tsoi, H.; et al. Transmission of Rat hepatitis E virus infection to humans in Hong Kong: A clinical and epidemiological analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ma, Z.; Harrison, T.J.; Feng, R.; Zhang, C.; Qiao, Z.; Fan, J.; Ma, H.; Li, M.; Song, A.; et al. A novel genotype of hepatitis E virus prevalent among farmed rabbits in China. J. Med. Virol. 2009, 81, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; International Committee on Taxonomy of Viruses Hepeviridae Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; Izopet, J.; Oliveira-Filho, E.F.; Ulrich, R.G.; Johne, R.; Koenig, M.; Jameel, S.; Harrison, T.J.; Meng, X.-J.; et al. Proposed reference sequences for hepatitis E virus subtypes. J. Gen. Virol. 2016, 97, 537–542. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.-J.; Norder, H.; Okamoto, H.; van der Poel, W.H.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Geng, J.; Fu, H.; Wang, L.; Bu, Q.; Liu, P.; Wang, M.; Sui, Y.; Wang, X.; Zhu, Y.; Zhuang, H. Phylogenetic analysis of the full genome of rabbit hepatitis E virus (rbHEV) and molecular biologic study on the possibility of cross species transmission of rbHEV. Infect. Genet. Evol. 2011, 11, 2020–2025. [Google Scholar] [CrossRef]
- Cossaboom, C.M.; Córdoba, L.; Cao, D.; Ni, Y.-Y.; Meng, X.-J. Complete genome sequence of hepatitis E virus from rabbits in the United States. J. Virol. 2012, 86, 13124–13125. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Zhang, H.; Li, J.; Huang, W.; Harrison, T.J.; Zhao, C.; Zhou, Y.; Lian, H.; Wang, Y. Comparison of hepatitis E virus genotypes from rabbits and pigs in the same geographic area: No evidence of natural cross-species transmission between the two animals. Infect. Genet. Evol. 2013, 13, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Burt, S.A.; Veltman, J.; Der Honing, R.H.-V.; Schmitt, H.; Van Der Poel, W.H.M. Hepatitis E virus in farmed rabbits, wild rabbits and petting farm rabbits in the Netherlands. Food Environ. Virol. 2016, 8, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Di Bartolo, I.; De Sabato, L.; Marata, A.; Martinelli, N.; Magistrali, C.F.; Monini, M.; Ponterio, E.; Ostanello, F.; Ruggeri, F.M. Serological survey of hepatitis E virus infection in farmed and pet rabbits in Italy. Arch. Virol. 2016, 161, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, F.; Schwaiger, K.; Dähnert, L.; Vina-Rodriguez, A.; Höper, D.; Gareis, M.; Groschup, M.H.; Eiden, M. Hepatitis E virus in wild rabbits and European brown hares in Germany. Zoonoses Public Health 2017, 64, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-H.; Park, B.-J.; Ahn, H.-S.; Kim, Y.-H.; Go, H.-J.; Kim, D.-H.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; et al. Evidence of hepatitis E virus infection in specific pathogen-free rabbits in Korea. Virus Genes 2018, 54, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.V.; Yonemitsu, K.; Ishijima, K.; Minami, S.; Supriyono; Tran, N.T.; Kuroda, Y.; Tatemoto, K.; Inoue, Y.; Okada, A.; et al. Characterization of rabbit hepatitis E virus isolated from a feral rabbit. Vet. Microbiol. 2021, 263, 109275. [Google Scholar] [CrossRef]
- Jirintai, S.; Jinshan; Tanggis; Manglai, D.; Mulyanto; Takahashi, M.; Nagashima, S.; Kobayashi, T.; Nishizawa, T.; Okamoto, H. Molecular analysis of hepatitis E virus from farm rabbits in Inner Mongolia, China and its successful propagation in A549 and PLC/PRF/5 cells. Virus Res. 2012, 170, 126–137. [Google Scholar] [CrossRef]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Jirintai, S.; Takahashi, M.; Okamoto, H.; Takeda, N.; Muramatsu, M.; Li, T. Persistent infection with a rabbit hepatitis E virus created by a reverse genetics system. Transbound. Emerg. Dis. 2020, 8, 615–625. [Google Scholar] [CrossRef]
- Cossaboom, C.M.; Córdoba, L.; Sanford, B.J.; Piñeyro, P.; Kenney, S.P.; Dryman, B.A.; Wang, Y.; Meng, X.-J. Cross-species infection of pigs with a novel rabbit, but not rat, strain of hepatitis E virus isolated in the United States. J. Gen. Virol. 2012, 93, 1687–1695. [Google Scholar] [CrossRef]
- Liu, P.; Bu, Q.-N.; Wang, L.; Han, J.; Du, R.-J.; Lei, Y.-X.; Ouyang, Y.-Q.; Li, J.; Zhu, Y.-H.; Lu, F.-M.; et al. Transmission of hepatitis E virus from rabbits to cynomolgus macaques. Emerg. Infect. Dis. 2013, 19, 559–565. [Google Scholar] [CrossRef]
- Abravanel, F.; Lhomme, S.; El Costa, H.; Schvartz, B.; Peron, J.-M.; Kamar, N.; Izopet, J. Rabbit hepatitis E virus infections in humans, France. Emerg. Infect. Dis. 2017, 23, 1191–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahli, R.; Fraga, M.; Semela, D.; Moradpour, D.; Gouttenoire, J. Rabbit HEV in immunosuppressed patients with hepatitis E acquired in Switzerland. J. Hepatol. 2019, 70, 1023–1025. [Google Scholar] [CrossRef] [Green Version]
- Lhomme, S.; Dubois, M.; Abravanel, F.; Top, S.; Bertagnoli, S.; Guerin, J.-L.; Izopet, J. Risk of zoonotic transmission of HEV from rabbits. J. Clin. Virol. 2013, 58, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Bai, H.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.-C. Immunogenicity and antigenicity of rabbit hepatitis E virus-like particles produced by recombinant baculoviruses. Viruses 2021, 13, 1573. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-C.; Yang, T.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Ishii, K.; Haga, K.; Nakamura, T.; Ochiai, S.; Takaji, W.; et al. Construction and characterization of an infectious cDNA clone of rat hepatitis E virus. J. Gen. Virol. 2015, 96, 1320–1327. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | For the First PCR | Name | For the Second PCR | Products, nt |
|---|---|---|---|---|
| F38 | TCGATGCCATGGAGGCCCA | F105 | GCCTTGGCGAATGCTGTGG | 105–469 |
| R560 | CATTGCCTCTGCCACATCAG | R469 | TCAAAGCAATAGGTTCGATC | |
| F373 | GCGATGGTATTCTGCCCCTA | F540 | CTGATGTGGCAGAGGCAATG | 373–1100 |
| R1128 | CATCTTCAGAGGCATTCCAG | R1110 | AGCTACAAGAGCACCAACAG | |
| F1337 | GCTTTCTGCCGGCTTTCATT | F1378 | TTGACGAATCTGTGCACTGC | 1378–1759 |
| R1805 | CTGAGCAGTACGTTCTCTCA | R1759 | TCAGTGATTGTGGTGCGGAA | |
| F2089 | GGGAGTCTACTAACCCGTTT | F2160 | AGCGACTTTTCACCACCTGA | 2160–2439 |
| R2519 | CTTGGTATGGTCGAAAGACT | R2439 | ATGCGTTAACCAGCCAGTCA | |
| F2501 | GTCTTTCGACCATACCAAGT | F2581 | TCGCCCCTGATTATAGAGTT | 2581–3040 |
| R3074 | CTGCTGTATGGACCTCGATT | R3040 | AACGCCTGCAGTGAATTGATA | |
| F3221 | CCACCTGCTTTTGTTGCATA | F3301 | TTGAGCATGCCGGTCTAGTC | 3301–3699 |
| R3840 | GATAGGCACTAATCTGGCAG | R3699 | ATATGCCCACCTCACGAAGT | |
| F4048 | GTATGGGCGTCGAACAAAGT | F4141 | CTACAACGTGCGAGCTTTAT | 4141–4359 |
| R4440 | CAGAGTCTTCATAGGCGTCA | R4359 | GCCCAAATAAGGCGCAGAAT | |
| F5020 | CGAATGTTGCTCAGGTGTGT | F5091 | CTTATTGGCATGTTACAGAC | 5091–5440 |
| R5280 | CAGCATAGGCAAACACACGA | R5440 | CGGAATGTGAGTCAACGTCA | |
| F5986 | TATCGTAACCAGGGGTGGCG | F6022 | GGTGTAGCCGAGGAGGAGGC | 6022–6650 |
| R6702 | AGGGTTAGTTGACGAGCCAT | R6650 | GAGAGCCACAACACATCGTT |
| Nucleotide Changes | Amino Acid Changes | |||||
|---|---|---|---|---|---|---|
| a Position | JP-59 | JP-59cr | b Position | ORF1 | ORF2 | ORF3 |
| 249 | C | T | ||||
| 368 | C | T | ||||
| 551 | G | A | ||||
| 800 | T | C | ||||
| 1040 | G | A | ||||
| 1439 | C | T | ||||
| 1613 | A | G | ||||
| 1652 | G | A | ||||
| 1723 | C | T | 566 | Thr/Ile | ||
| 2261 | C | T | 749 | Gly/Ser | ||
| 2271 | G | A | ||||
| 2690 | C | T | ||||
| 2930 | C | T | ||||
| 3455 | T | C | ||||
| 4304 | C | T | ||||
| 5205 | C | T | 4 | Ser/Phe | ||
| 6351 | C | T | ||||
| 6594 | A | G | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Mendoza, M.V.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Maeda, K.; Li, T. Low Replication Efficiency of a Japanese Rabbit Hepatitis E Virus Strain in the Human Hepatocarcinoma Cell Line PLC/PRF/5. Viruses 2023, 15, 1322. https://doi.org/10.3390/v15061322
Zhang W, Mendoza MV, Ami Y, Suzaki Y, Doan YH, Maeda K, Li T. Low Replication Efficiency of a Japanese Rabbit Hepatitis E Virus Strain in the Human Hepatocarcinoma Cell Line PLC/PRF/5. Viruses. 2023; 15(6):1322. https://doi.org/10.3390/v15061322
Chicago/Turabian StyleZhang, Wenjing, Milagros Virhuez Mendoza, Yasushi Ami, Yuriko Suzaki, Yen Hai Doan, Ken Maeda, and Tiancheng Li. 2023. "Low Replication Efficiency of a Japanese Rabbit Hepatitis E Virus Strain in the Human Hepatocarcinoma Cell Line PLC/PRF/5" Viruses 15, no. 6: 1322. https://doi.org/10.3390/v15061322