
Super-Resolution Microscopy Analysis of Hepatitis B Viral cccDNA and Host Factors
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. HBV Infection of Primary Human Hepatocyte
2.3. Knockdown of DOCK11 Using shRNA
2.4. Overexpression of DOCK11
2.5. Quantitative Real-Time PCR
2.6. Western Blotting
2.7. Chromatin Immunoprecipitation
2.8. FISH of Viral RNA, Relaxed Circular DNA, and cccDNA
2.9. Immunofluorescence
2.10. Statistical Analysis
3. Results
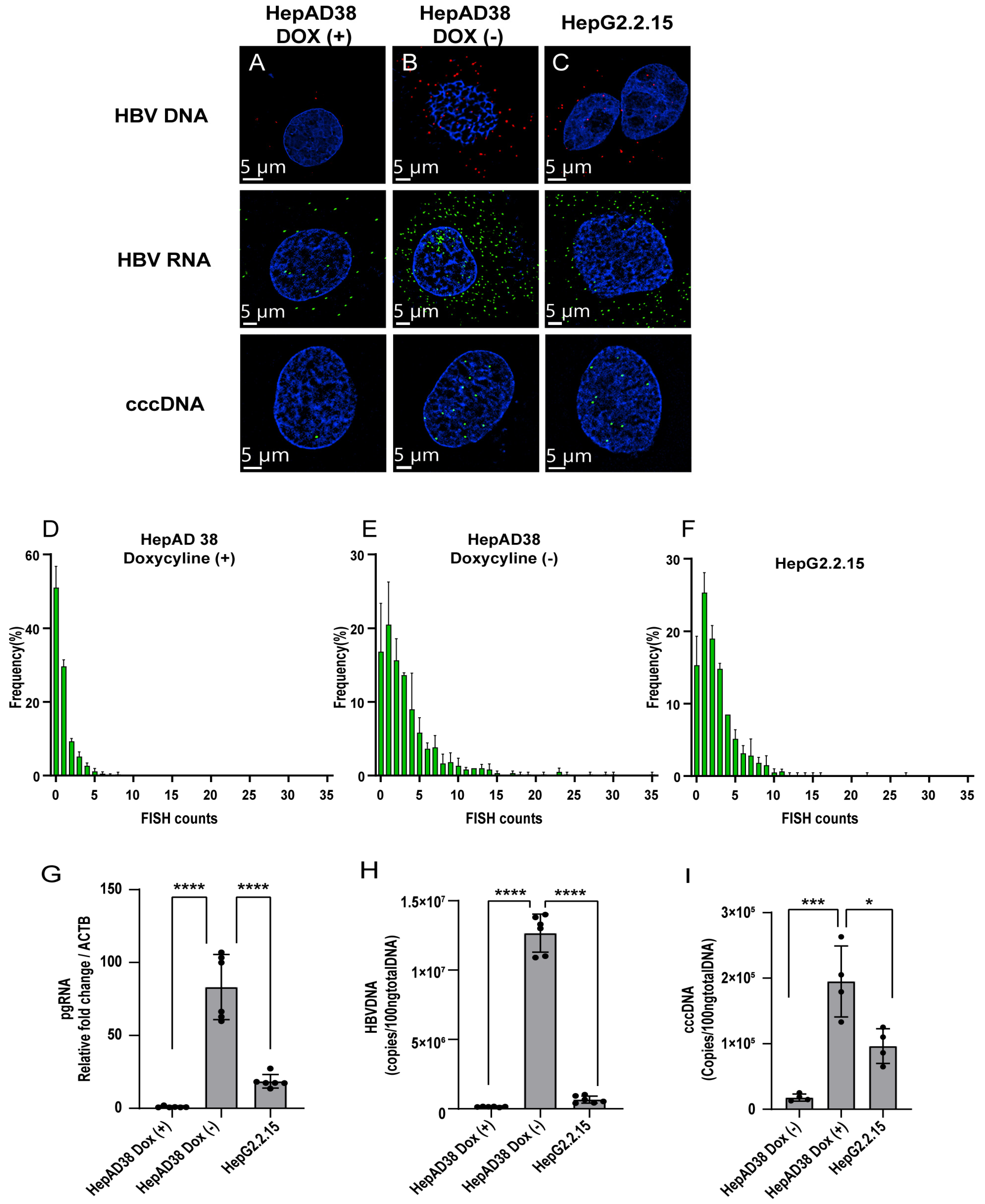
3.1. FISH with Branched DNA Amplification Facilitates the Subcellular Localization of Key HBV Nucleic Acids in HBV-Inducible Hepatocytes by Super-Resolution Confocal Microscopy
3.2. Detection of HBV Nucleic Acids in HBV-Replication Cell Lines
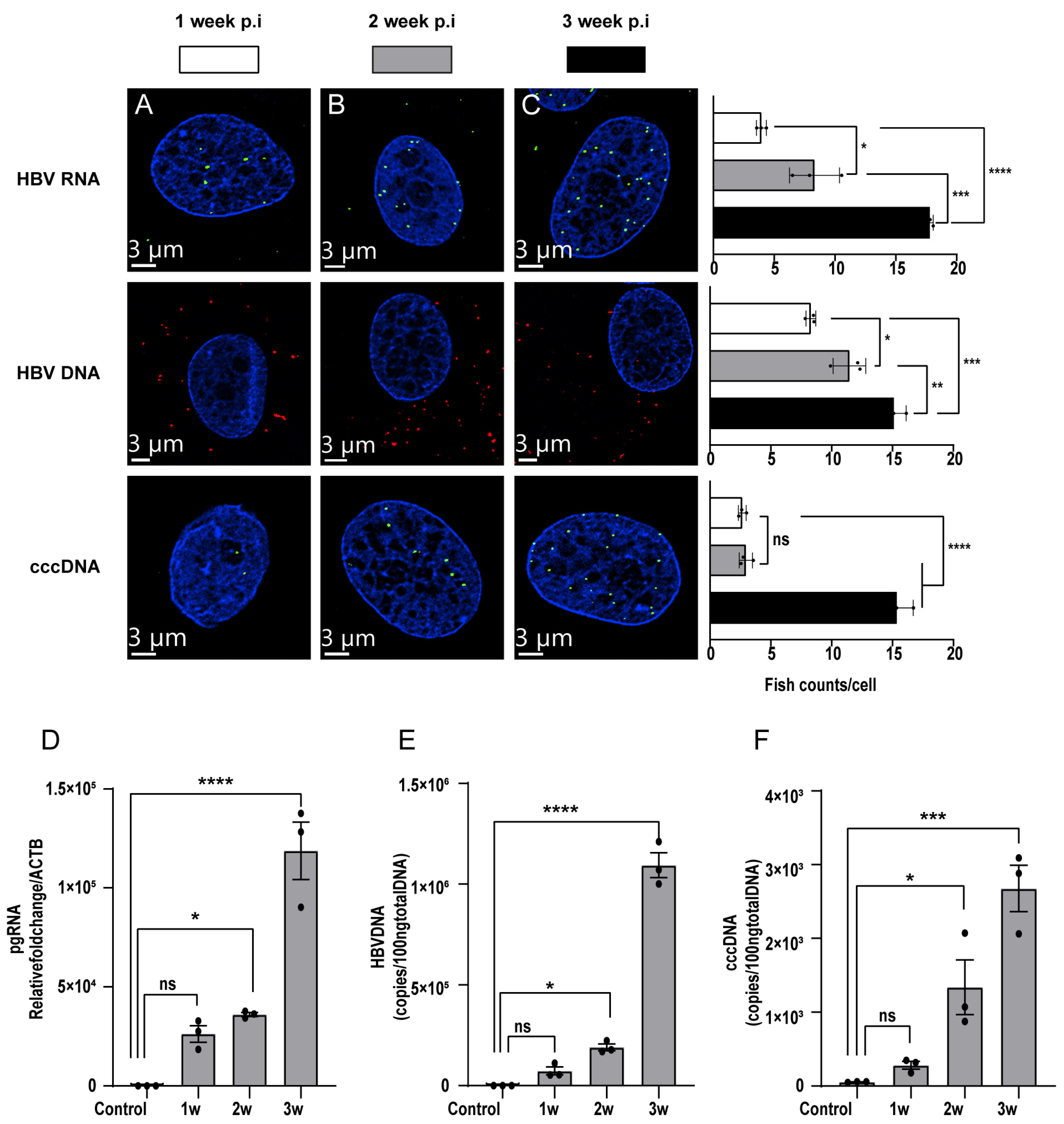
3.3. Detection of HBV Nucleic Acids in HBV-Infected PXB-Cells
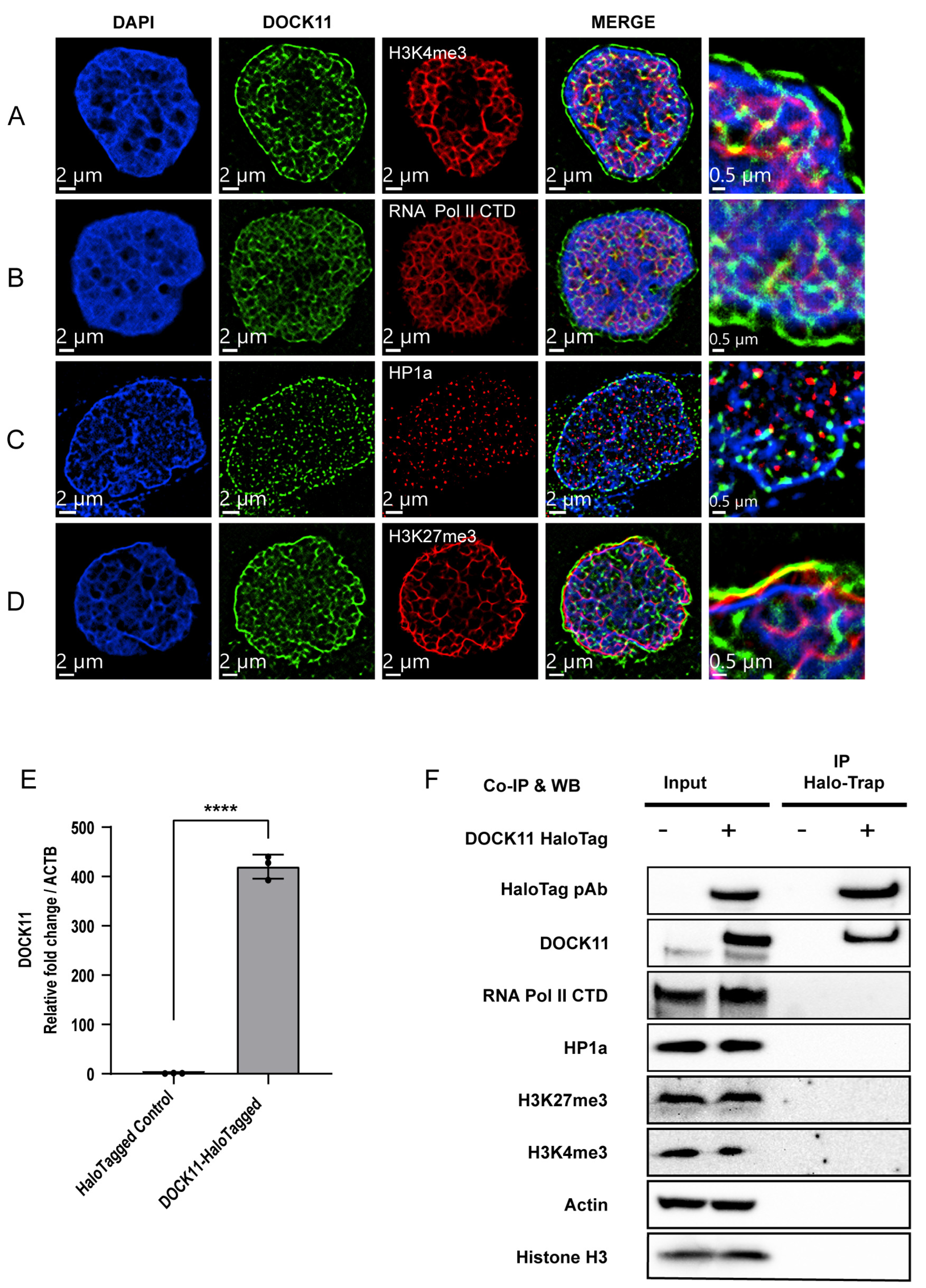
3.4. Determination of DOCK11 Partners for Promoting cccDNA Replication
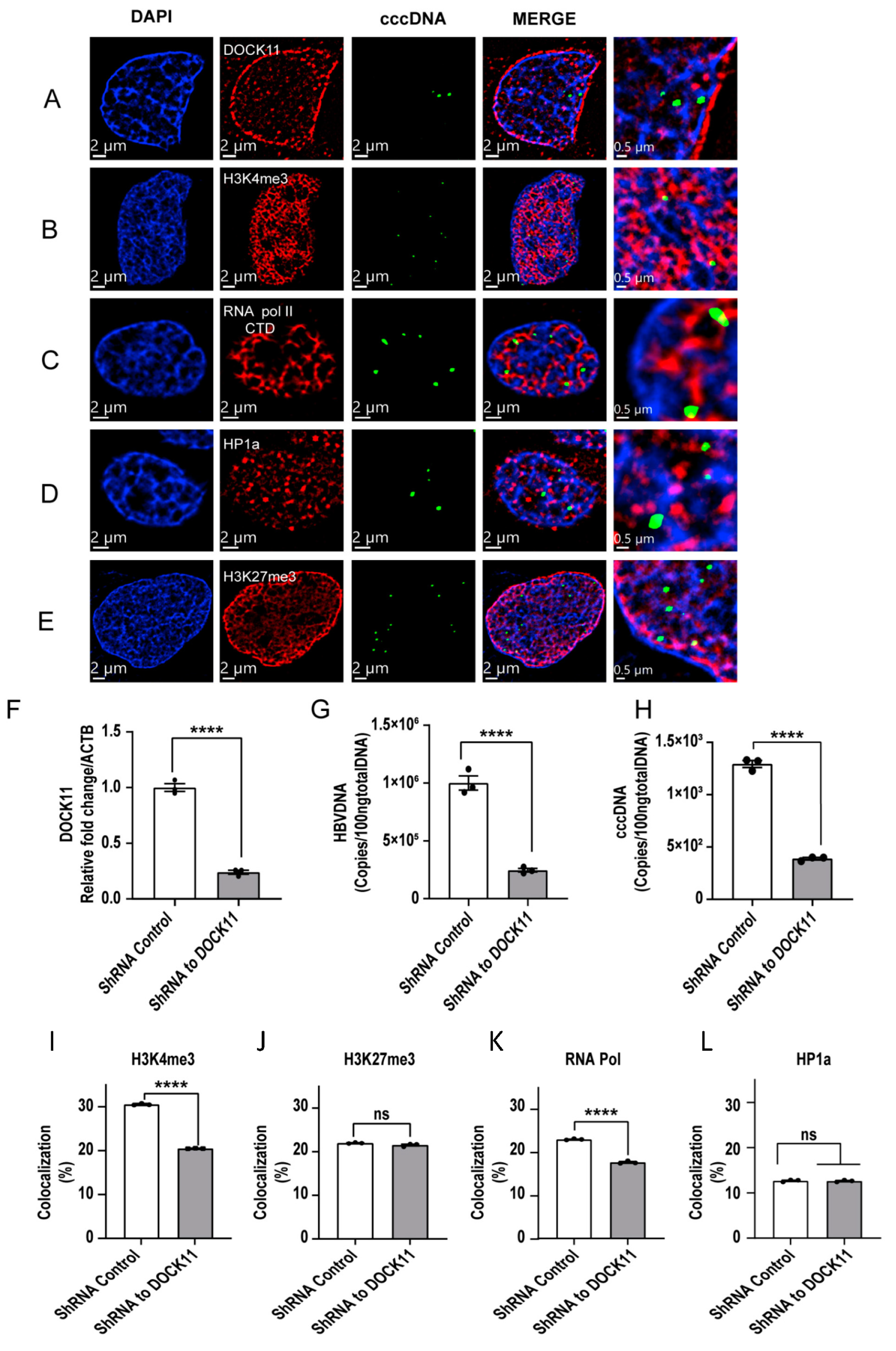
3.5. Localization of cccDNA with DOCK11 and Histone Markers Analyzed by Dragonfly Confocal Microscopy
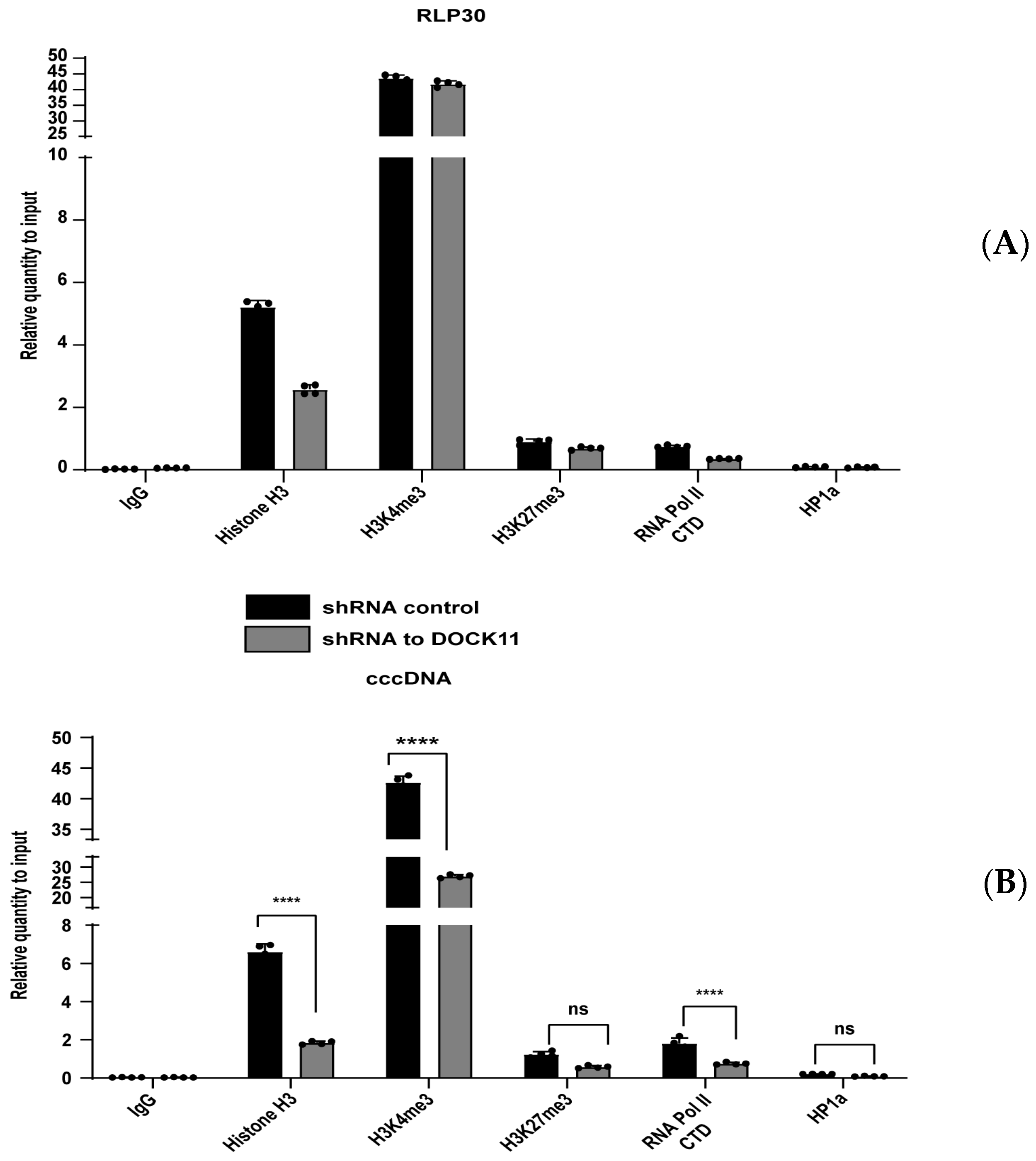
3.6. DOCK11 Knockdown Decreases cccDNA-Associated H3K4me3 and Pol II C-Terminal Domain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Hepatitis B. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 1 November 2022).
- Bock, C.T.; Schranz, P.; Schroder, C.H.; Zentgraf, H. Hepatitis B virus genome is organized into nucleosomes in the nucleus of the infected cell. Virus Genes 1994, 8, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.T.; Schwinn, S.; Locarnini, S.; Fyfe, J.; Manns, M.P.; Trautwein, C.; Zentgraf, H. Structural organization of the hepatitis B virus minichromosome. J. Mol. Biol. 2001, 307, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Li, S.; Shen, F.; Li, H.; Qian, S.; Lee, D.H.; Wu, J.Z.; Yang, W. Characterization of nucleosome positioning in hepadnaviral covalently closed circular DNA minichromosomes. J. Virol. 2012, 86, 10059–10069. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Pollicino, T.; Belloni, L.; Raffa, G.; Pediconi, N.; Squadrito, G.; Raimondo, G.; Levrero, M. Hepatitis B virus replication is regulated by the acetylation status of hepatitis B virus cccDNA-bound H3 and H4 histones. Gastroenterology 2006, 130, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Lebosse, F.; Inchauspe, A.; Locatelli, M.; Miaglia, C.; Diederichs, A.; Fresquet, J.; Chapus, F.; Hamed, K.; Testoni, B.; Zoulim, F. Quantification and epigenetic evaluation of the residual pool of hepatitis B covalently closed circular DNA in long-term nucleoside analogue-treated patients. Sci. Rep. 2020, 10, 21097. [Google Scholar] [CrossRef]
- Riviere, L.; Gerossier, L.; Ducroux, A.; Dion, S.; Deng, Q.; Michel, M.L.; Buendia, M.A.; Hantz, O.; Neuveut, C. HBx relieves chromatin-mediated transcriptional repression of hepatitis B viral cccDNA involving SETDB1 histone methyltransferase. J. Hepatol. 2015, 63, 1093–1102. [Google Scholar] [CrossRef]
- Rall, L.B.; Standring, D.N.; Laub, O.; Rutter, W.J. Transcription of hepatitis B virus by RNA polymerase II. Mol. Cell Biol. 1983, 3, 1766–1773. [Google Scholar]
- Hashimoto, S.; Shirasaki, T.; Yamashita, T.; Iwabuchi, S.; Suzuki, Y.; Takamura, Y.; Ukita, Y.; Deshimaru, S.; Okayama, T.; Ikeo, K.; et al. DOCK11 and DENND2A play pivotal roles in the maintenance of hepatitis B virus in host cells. PLoS ONE 2021, 16, e0246313. [Google Scholar] [CrossRef]
- Zhang, X.; Yue, L.; Li, C.; Yuan, Z. A Fluorescent In Situ Hybridization (FISH) Assay for Detection of HBV DNA in Cell Culture Models. In ICE-HBV. Available online: https://ice-hbv.org/protocol/a-fluorescent-in-situ-hybridization-fish-assay-for-detection-of-hbv-dna-in-cell-culture-models/?pdf=1 (accessed on 1 November 2021).
- Zhang, X.; Lu, W.; Zheng, Y.; Wang, W.; Bai, L.; Chen, L.; Feng, Y.; Zhang, Z.; Yuan, Z. In situ analysis of intrahepatic virological events in chronic hepatitis B virus infection. J. Clin. Investig. 2016, 126, 1079–1092. [Google Scholar] [CrossRef]
- Guo, J.T.; Pryce, M.; Wang, X.; Barrasa, M.I.; Hu, J.; Seeger, C. Conditional replication of duck hepatitis B virus in hepatoma cells. J. Virol. 2003, 77, 1885–1893. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Yang, W.; Baird, D.; Feng, Q.; Cerione, R.A. Identification of a DOCK180-related guanine nucleotide exchange factor that is capable of mediating a positive feedback activation of Cdc42. J. Biol. Chem. 2006, 281, 35253–35262. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, N.; Culley, S.; Ashdown, G.; Owen, D.M.; Pereira, P.M.; Henriques, R. Fast live-cell conventional fluorophore nanoscopy with ImageJ through super-resolution radial fluctuations. Nat. Commun. 2016, 7, 12471. [Google Scholar] [CrossRef]
- Cox, S.; Rosten, E.; Monypenny, J.; Jovanovic-Talisman, T.; Burnette, D.T.; Lippincott-Schwartz, J.; Jones, G.E.; Heintzmann, R. Bayesian localization microscopy reveals nanoscale podosome dynamics. Nat. Methods 2011, 9, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Tropberger, P.; Mercier, A.; Robinson, M.; Zhong, W.; Ganem, D.E.; Holdorf, M. Mapping of histone modifications in episomal HBV cccDNA uncovers an unusual chromatin organization amenable to epigenetic manipulation. Proc. Natl. Acad. Sci. USA 2015, 112, E5715–E5724. [Google Scholar] [CrossRef]
- Hong, X.; Kim, E.S.; Guo, H. Epigenetic regulation of hepatitis B virus covalently closed circular DNA: Implications for epigenetic therapy against chronic hepatitis B. Hepatology 2017, 66, 2066–2077. [Google Scholar] [CrossRef] [PubMed]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible expression of human hepatitis B virus (HBV) in stably transfected hepatoblastoma cells: A novel system for screening potential inhibitors of HBV replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [CrossRef]
- Peng, B.; Jing, Z.; Zhou, Z.; Sun, Y.; Guo, G.; Tan, Z.; Diao, Y.; Yao, Q.; Ping, Y.; Li, X.; et al. Nonproductive Hepatitis B Virus Covalently Closed Circular DNA Generates HBx-Related Transcripts from the HBx/Enhancer I Region and Acquires Reactivation by Superinfection in Single Cells. J. Virol. 2023, 97, e0171722. [Google Scholar] [CrossRef]
- Li, M.; Sohn, J.A.; Seeger, C. Distribution of Hepatitis B Virus Nuclear DNA. J. Virol. 2018, 92, E01391–E1417. [Google Scholar] [CrossRef]
- Zhang, H.; Tu, T. Approaches to quantifying hepatitis B virus covalently closed circular DNA. Clin. Mol. Hepatol. 2022, 28, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Allweiss, L.; Testoni, B.; Yu, M.; Lucifora, J.; Ko, C.; Qu, B.; Lütgehetmann, M.; Guo, H.; Urban, S.; Fletcher, S.P.; et al. Quantification of the hepatitis B virus cccDNA: Evidence-based guidelines for monitoring the key obstacle of HBV cure. Gut 2023, 72, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Zhao, H.; Wu, Y.; Peng, B.; Gao, Z.; Sun, Y.; Duan, J.; Qi, Y.; Li, Y.; Zhou, Z.; et al. Transcriptionally inactive hepatitis B virus episome DNA preferentially resides in the vicinity of chromosome 19 in 3D host genome upon infection. Cell Rep. 2021, 35, 109288. [Google Scholar] [CrossRef] [PubMed]
- Schoelz, J.M.; Riddle, N.C. Functions of HP1 proteins in transcriptional regulation. Epigenetics Chromatin 2022, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.L. Ras and Rho GTPase regulation of Pol II transcription: A shortcut model revisited. Transcription 2017, 8, 268–274. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, G.; Chen, Y.; Zhao, Y.; Gao, P.; Liu, B.; Wang, H.; Zheng, Z.L. C-terminal domain (CTD) phosphatase links Rho GTPase signaling to Pol II CTD phosphorylation in Arabidopsis and yeast. Proc. Natl. Acad. Sci. USA 2016, 113, E8197–E8206. [Google Scholar] [CrossRef]
- Chen, L.F.; Mu, Y.; Greene, W.C. Acetylation of RelA at discrete sites regulates distinct nuclear functions of NF-kappaB. EMBO J. 2002, 21, 6539–6548. [Google Scholar] [CrossRef]
- Yang, X.D.; Huang, B.; Li, M.; Lamb, A.; Kelleher, N.L.; Chen, L.F. Negative regulation of NF-kappaB action by Set9-mediated lysine methylation of the RelA subunit. EMBO J. 2009, 28, 1055–1066. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (cccDNA/cell) | HepAD38 (Dox+) | HepAD38 (Dox−) | HepG2.2.15 | |
|---|---|---|---|---|
| FISH counts | Average | 1.51 | 5.59 | 3.9 |
| Median | 4 | 11 | 11.5 | |
| Min./Max | 0/8 | 0/31 | 0/32 | |
| PCR measurement | Average | 1.07 | 11.1 | 5.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doan, P.T.B.; Nio, K.; Shimakami, T.; Kuroki, K.; Li, Y.-Y.; Sugimoto, S.; Takayama, H.; Okada, H.; Kaneko, S.; Honda, M.; et al. Super-Resolution Microscopy Analysis of Hepatitis B Viral cccDNA and Host Factors. Viruses 2023, 15, 1178. https://doi.org/10.3390/v15051178
Doan PTB, Nio K, Shimakami T, Kuroki K, Li Y-Y, Sugimoto S, Takayama H, Okada H, Kaneko S, Honda M, et al. Super-Resolution Microscopy Analysis of Hepatitis B Viral cccDNA and Host Factors. Viruses. 2023; 15(5):1178. https://doi.org/10.3390/v15051178
Chicago/Turabian StyleDoan, Phuong Thi Bich, Kouki Nio, Tetsuro Shimakami, Kazuyuki Kuroki, Ying-Yi Li, Saiho Sugimoto, Hideo Takayama, Hikari Okada, Shuichi Kaneko, Masao Honda, and et al. 2023. "Super-Resolution Microscopy Analysis of Hepatitis B Viral cccDNA and Host Factors" Viruses 15, no. 5: 1178. https://doi.org/10.3390/v15051178