
Virulence Profiles of Wild-Type, P.1 and Delta SARS-CoV-2 Variants in K18-hACE2 Transgenic Mice

 , ,
, ,  ,
,  , and
, and 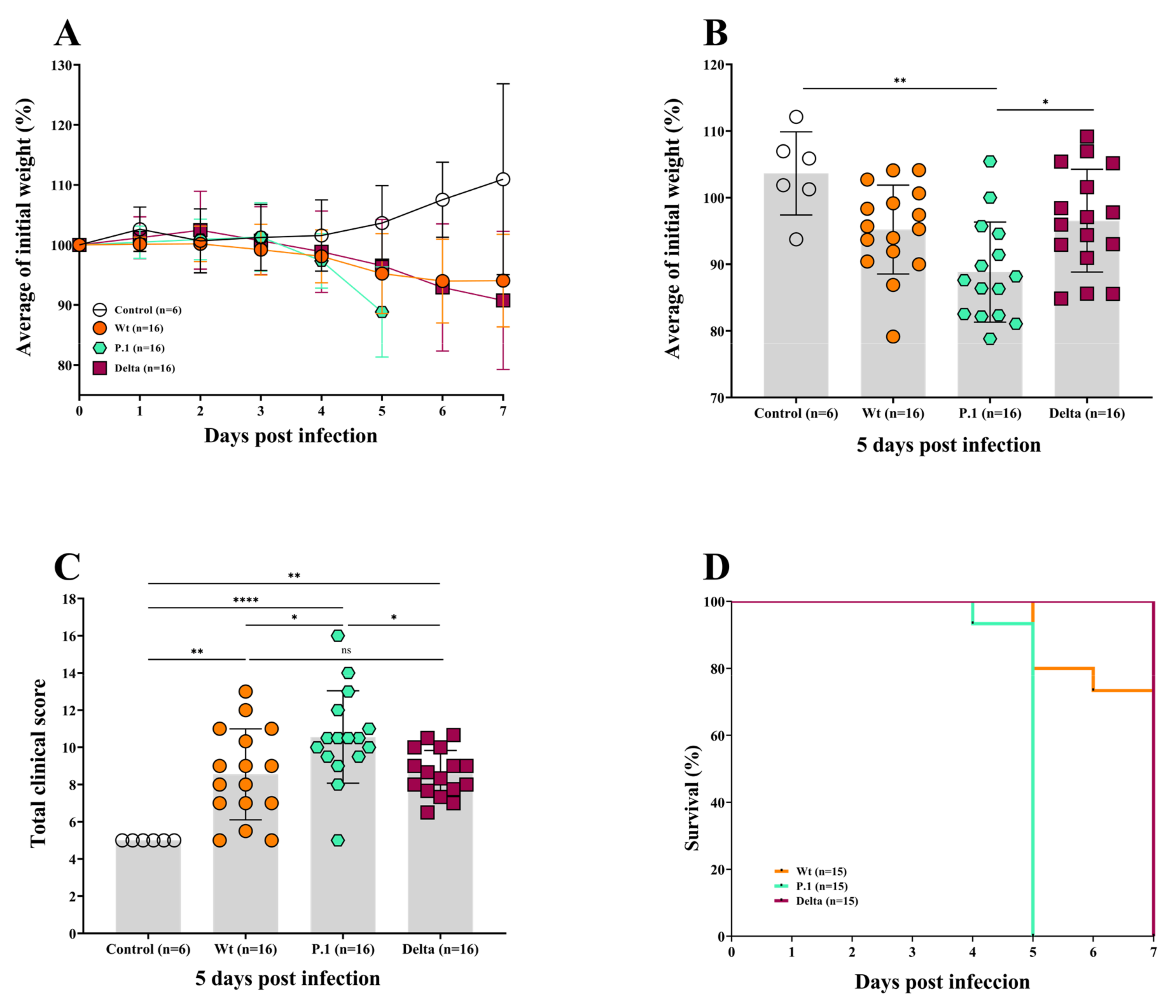{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Infection with the Wt SARS-CoV-2, P.1, and Delta variants and the Experimental Groups
2.3. The Clinical Evaluation of the SARS-CoV-2 Infection
2.4. Whole-Body Plethysmography Evaluation
2.5. Quantification of the Viral Load by RT-qPCR
2.6. Histopathology Analysis
2.7. Statistical Analysis
3. Results
3.1. The P.1 Variant Induces a More Severe Disease Than the Wt Strain and Delta Variant in ACE2-Transgenic Mice
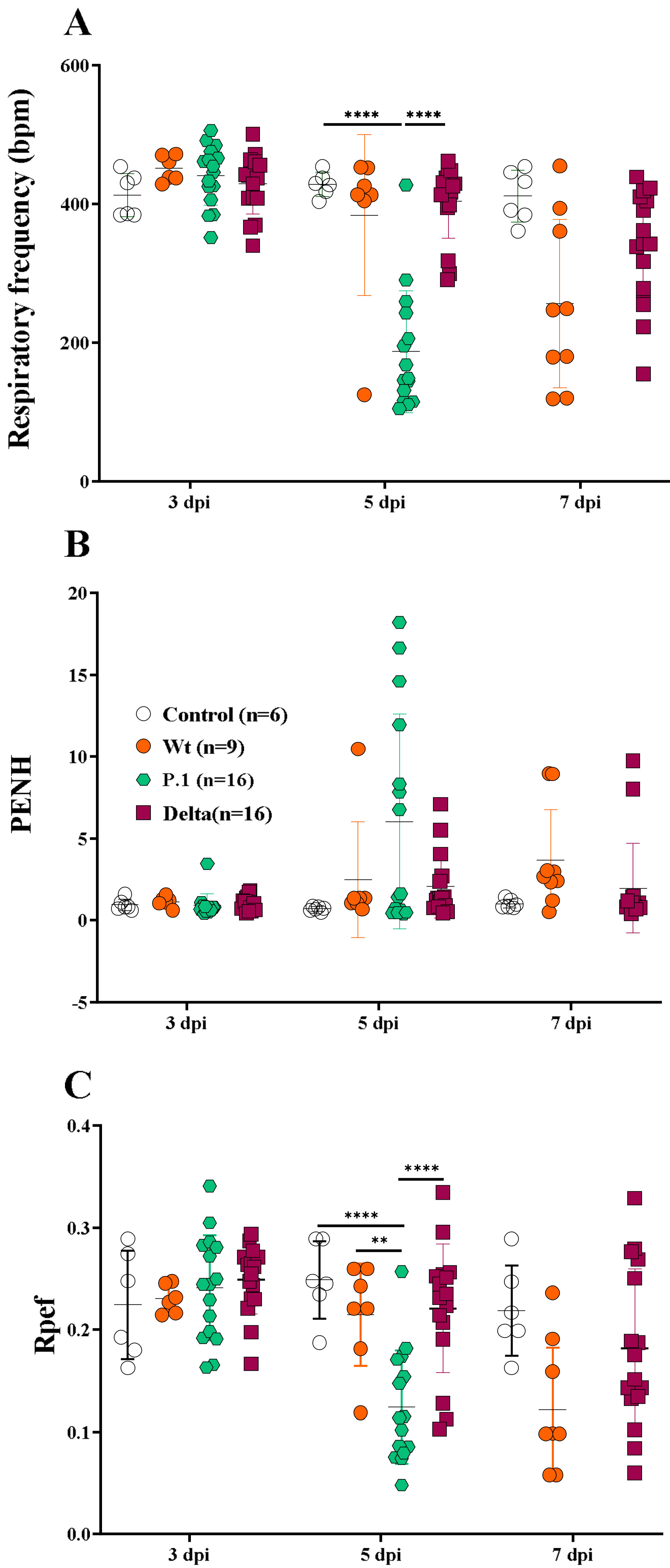
3.2. The Whole-Body Plethysmography Demonstrates That the SARS-CoV-2 P.1 Variant Induces More Severe Lung Dysfunction
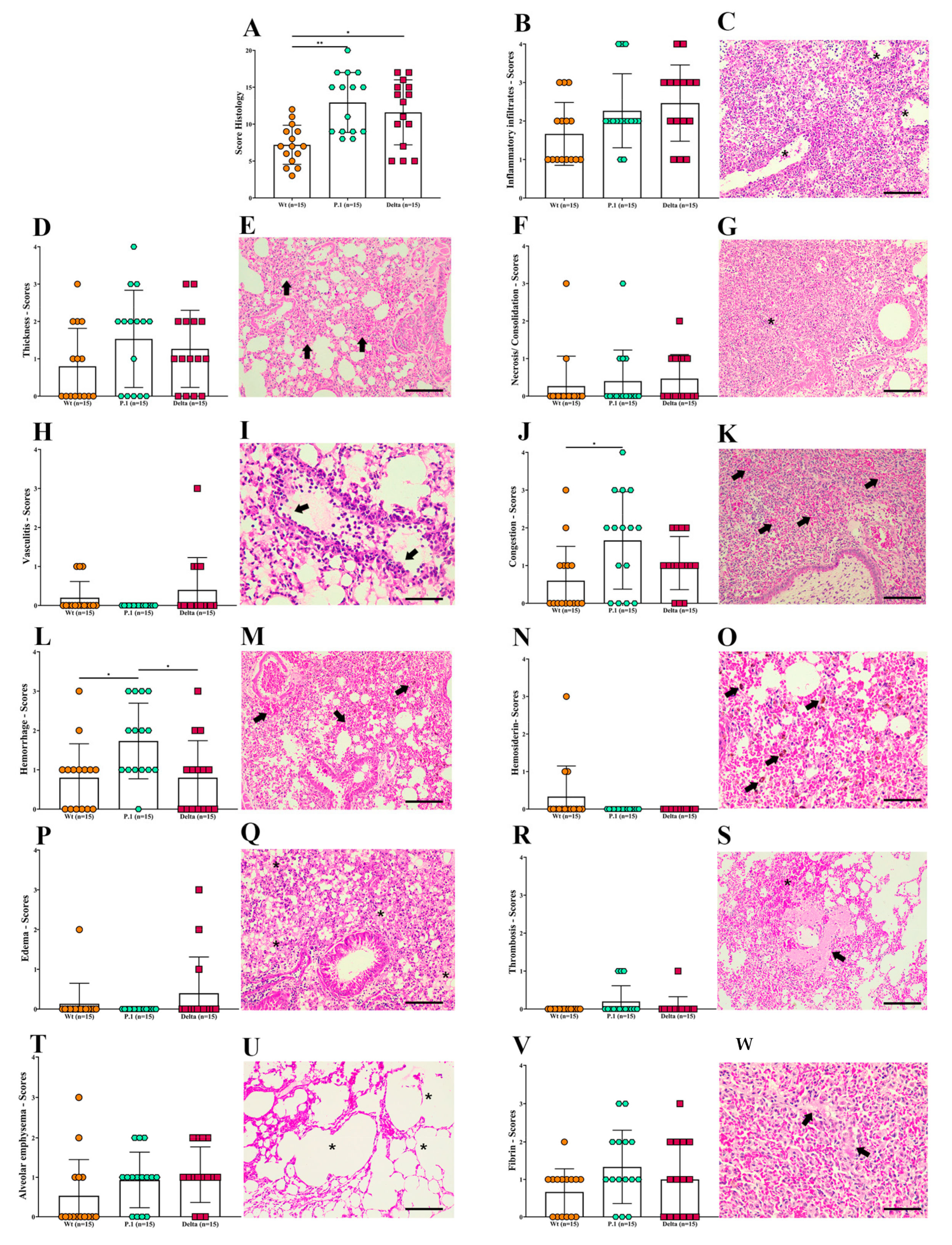
3.3. Histopathological Changes in the Lungs of K18-hACE2 Mice after Infection with the Wt Strain or the P.1 and Delta Variants
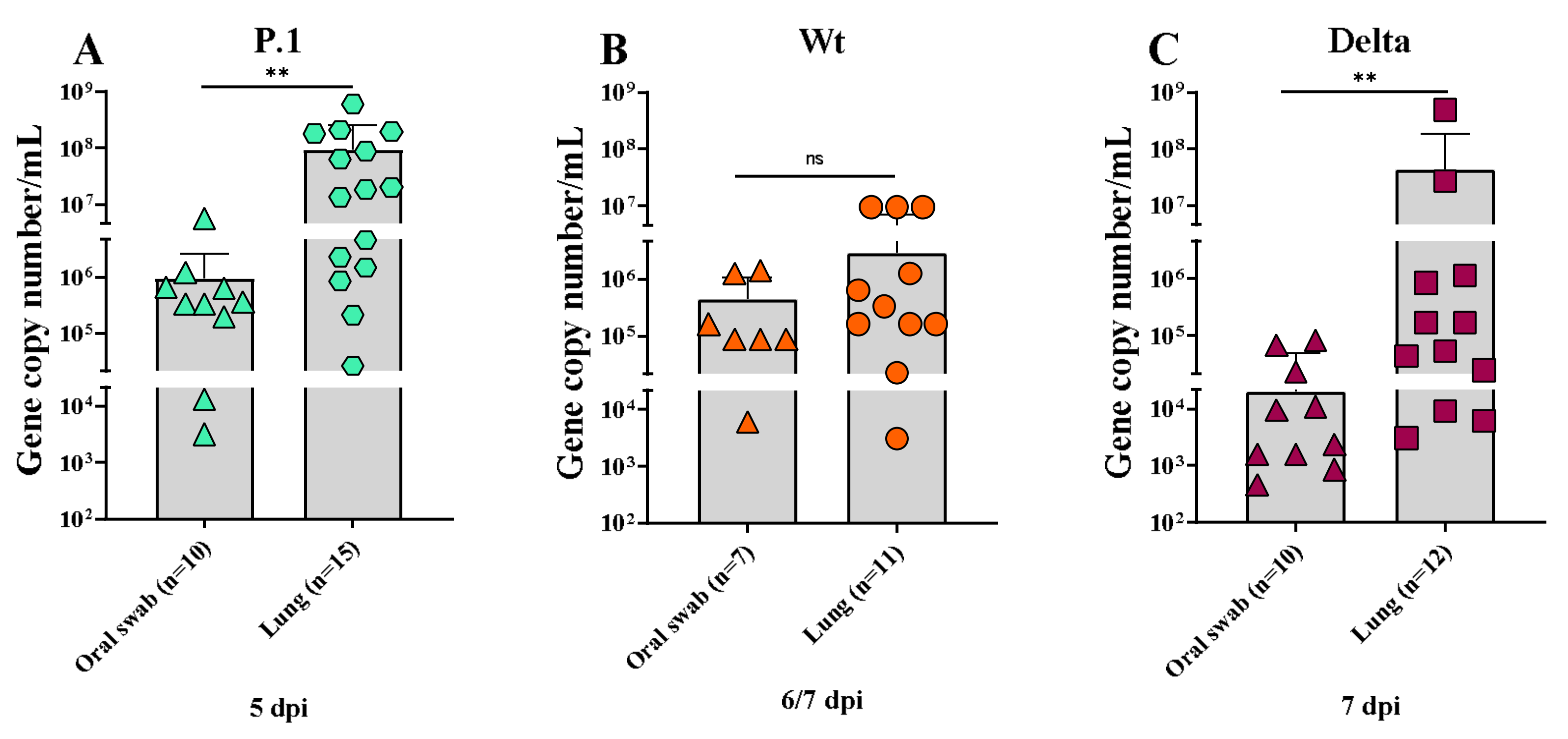
3.4. Quantification of SARS-CoV-2 in the Lungs and Oral Swabs of K18-hACE2 Mice Infected with the SARS-CoV-2 Wt, P.1, and Delta Lineages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johns Hopkins University & Medicine sCOVID-19 Dashboard John Hopkins. Available online: https://coronavirus.jhu.edu/map.html (accessed on 27 March 2023).
- Caramaschi, S.; Kapp, M.E.; Miller, S.E.; Eisenberg, R.; Johnson, J.; Epperly, G.; Maiorana, A.; Silvestri, G.; Giannico, G.A. Histopathological Findings and Clinicopathologic Correlation in COVID-19: A Systematic Review. Mod. Pathol. 2021, 34, 1614–1633. [Google Scholar] [CrossRef] [PubMed]
- Umakanthan, S.; Sahu, P.; Ranade, A.V.; Bukelo, M.M.; Rao, J.S.; Abrahao-Machado, L.F.; Dahal, S.; Kumar, H.; Kv, D. Origin, Transmission, Diagnosis and Management of Coronavirus Disease 2019 (COVID-19). Postgrad. Med. J. 2020, 96, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Merdji, H.; Mayeur, S.; Schenck, M.; Oulehri, W.; Clere-Jehl, R.; Cunat, S.; Herbrecht, J.-E.; Janssen-Langenstein, R.; Nicolae, A.; Helms, J.; et al. Histopathological Features in Fatal COVID-19 Acute Respiratory Distress Syndrome. Med. Intensiv. (Engl. Ed.) 2021, 45, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Angeles Montero-Fernandez, M.; Pardo-Garcia, R. Histopathology Features of the Lung in COVID-19 Patients. Diagn. Histopathol. 2021, 27, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Oladunni, F.S.; Park, J.G.; Pino, P.A.; Gonzalez, O.; Akhter, A.; Allué-Guardia, A.; Olmo-Fontánez, A.; Gautam, S.; Garcia-Vilanova, A.; Ye, C.; et al. Lethality of SARS-CoV-2 Infection in K18 Human Angiotensin-Converting Enzyme 2 Transgenic Mice. Nat. Commun. 2020, 11, 6122. [Google Scholar] [CrossRef]
- Deshmukh, V.; Motwani, R.; Kumar, A.; Kumari, C.; Raza, K. Histopathological Observations in COVID-19: A Systematic Review. J. Clin. Pathol. 2021, 74, 76–83. [Google Scholar] [CrossRef]
- Tarrés-Freixas, F.; Trinité, B.; Pons-Grífols, A.; Romero-Durana, M.; Riveira-Muñoz, E.; Ávila-Nieto, C.; Pérez, M.; Garcia-Vidal, E.; Pérez-Zsolt, D.; Muñoz-Basagoiti, J.; et al. Heterogeneous Infectivity and Pathogenesis of SARS-CoV-2 Variants Beta, Delta and Omicron in Transgenic K18-hACE2 Mice and Wildtype Mice. Front. Microbiol. 2022, 13, 840757. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Flight, K.E.; Higham, S.L.; Wang, Z.; Pierce, B.F. Progress of the COVID-19 Vaccine Effort: Viruses, Vaccines and Variants versus Efficacy, Effectiveness and Escape. Nat. Rev. Immunol. 2021, 21, 626–636. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The Proximal Origin of SARS-CoV-2. Nat. Med. 2020, 46, 450–452. [Google Scholar] [CrossRef]
- Lin, L.; Liu, Y.; Tang, X.; He, D. The Disease Severity and Clinical Outcomes of the SARS-CoV-2 Variants of Concern. Front. Public Health 2021, 9, 775224. [Google Scholar] [CrossRef]
- Buchan, S.A.; Chung, H.; Brown, K.A.; Austin, P.C.; Fell, D.B.; Gubbay, J.B.; Nasreen, S.; Schwartz, K.L.; Sundaram, M.E.; Tadrous, M.; et al. Estimated Effectiveness of COVID-19 Vaccines against Omicron or Delta Symptomatic Infection and Severe Outcomes. Jama Network Open 2022, 5, e2232760. [Google Scholar] [CrossRef]
- Cromer, D.; Juno, J.A.; Khoury, D.; Reynaldi, A.; Wheatley, A.K.; Kent, S.J.; Davenport, M.P. Prospects for Durable Immune Control of SARS-CoV-2 and Prevention of Reinfection. Nat. Rev. Immunol. 2021, 21, 395–404. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Johnson, B.A.; Xia, H.; Ku, Z.; Schindewolf, C.; Widen, S.G.; An, Z.; Weaver, S.C.; Menachery, V.D.; et al. Delta Spike P681R Mutation Enhances SARS-CoV-2 Fitness over Alpha Variant. bioRxiv 2021, 1, 1–29. [Google Scholar] [CrossRef]
- UK Health Security Agency (UKHSA). SARS-CoV-2 Variants of Concern and Variants under Investigation in England- Technical Briefing 31; Sage: London, UK, 2021. [Google Scholar]
- Burki, T. The Origin of SARS-CoV-2 Variants of Concern. Lancet Infect. Dis. 2022, 22, 174–175. [Google Scholar] [CrossRef]
- Ferguson, N.; Ghani, A.; Cori, A.; Hogan, A.; Hinsley, W.; Volz, E. Report 49: Growth, Population Distribution and Immune Escape of Omicron in England; Imperial College London: London, UK, 2021; pp. 1–10. [Google Scholar]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The Biological and Clinical Significance of Emerging SARS-CoV-2 Variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated Transmissibility and Impact of SARS-CoV-2 Lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Ramundo, M.S.; De Jesus, J.G.; Andrade, P.S.; Coletti, T.M. Genomics and Epidemiology of the P.1 SARS-CoV-2 Lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar]
- Tosta, S.; Giovanetti, M.; Brandão Nardy, V.; Reboredo de Oliveira da Silva, L.; Kelly Astete Gómez, M.; Gomes Lima, J.; Cardoso, C.W.; Silva, T.O.; de Souza, M.S.P.L.; Dia, P.H.P.; et al. Short Report: Early Genomic Detection of Sars-Cov-2 p.1 Variant in Northeast Brazil. PLoS Negl. Trop. Dis. 2021, 15, e0011024. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 Variant of Concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Prete, C.A.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U.G.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite High Seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Malani, P.N. Variants of SARS-CoV-2. JAMA 2021, 326, 880. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, J.G.; Barateiro, A.; Peixoto, E.P.M.; Barros, A.B.C.D.S.; de Souza, R.M.; Clark, T.G.; Campino, S.; Wrenger, C.; Wunderlich, G.; Palmisano, G.; et al. Adverse Pregnancy Outcomes Are Associated with Plasmodium Vivax Malaria in a Prospective Cohort of Women from the Brazilian Amazon. PLoS Negl. Trop. Dis. 2021, 15, e0009390. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and Epidemiology of a Novel SARS-CoV-2 Lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Kistler, K.; Huddleston, J.; Bedford, T. Rapid and Parallel Adaptive Mutations in Spike S1 Drive Clade Success in SARS-CoV-2. Cell Host Microbe. 2022, 30, 545–555. [Google Scholar] [CrossRef]
- Imai, M.; Halfmann, P.J.; Yamayoshi, S.; Iwatsuki-horimoto, K.; Chiba, S. Characterization of a New SARS-CoV-2 Variant That Emerged in Brazil. Proc. Natl. Acad. Sci. USA 2021, 118, e2106535118. [Google Scholar] [CrossRef]
- Nonaka, C.K.V.; Gräf, T.; de Lorenzo Barcia, C.A.; Costa, V.F.; de Oliveira, J.L.; da Passos, R.H.; Bastos, I.N.; de Santana, M.C.B.; Santos, I.M.; de Sousa, K.A.F.; et al. SARS-CoV-2 Variant of Concern P.1 (Gamma) Infection in Young and Middle-Aged Patients Admitted to the Intensive Care Units of a Single Hospital in Salvador, Northeast Brazil, February 2021. Int. J. Infect. Dis. 2021, 111, 47–54. [Google Scholar] [CrossRef]
- Keegan, L.T.; Truelove, S.; Lessler, J. Analysis of Vaccine Effectiveness against COVID-19 and the Emergence of Delta and Other Variants of Concern in Utah. JAMA Netw. Open 2021, 4, 12–15. [Google Scholar] [CrossRef]
- McKeigue, P.M.; McAllister, D.A.; Hutchinson, S.J.; Robertson, C.; Stockton, D.; Colhoun, H.M. Vaccine Efficacy against Severe COVID-19 in Relation to Delta Variant (B.1.617.2) and Time since Second Dose in Patients in Scotland (REACT-SCOT): A Case-Control Study. Lancet Respir. Med. 2022, 2600, 566–572. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Zhang, L.; Wan, S.; Zhang, L.; Zhou, F. SARS-CoV-2 Omicron Variant: Recent Progress and Future Perspectives. Signal Transduct. Target. Ther. 2022, 7, 141. [Google Scholar] [CrossRef]
- Plante, J.A.; Mitchell, B.M.; Plante, K.S.; Debbink, K.; Weaver, S.C.; Menachery, V.D. The Variant Gambit: COVID-19′s next Move. Cell Host Microbe 2021, 29, 508–515. [Google Scholar] [CrossRef]
- Jacobs, J.L.; Haidar, G.; Mellors, J.W. COVID-19: Challenges of Viral Variants. Annu. Rev. Med. 2023, 74, 31–53. [Google Scholar] [CrossRef]
- McCray, P.B.; Pewe, L.; Wohlford-Lenane, C.; Hickey, M.; Manzel, L.; Shi, L.; Netland, J.; Jia, H.P.; Halabi, C.; Sigmund, C.D.; et al. Lethal Infection of K18-HACE2 Mice Infected with Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2007, 81, 813–821. [Google Scholar] [CrossRef]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.d.M.; de Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; de Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 Isolation from the First Reported Patients in Brazil and Establishment of a Coordinated Task Network. Mem. Inst. Oswaldo Cruz 2020, 115, 1–8. [Google Scholar] [CrossRef]
- Yang, X.-H.; Deng, W.; Tong, Z.; Liu, Y.-X.; Zhang, L.-F.; Zhu, H.; Gao, H.; Huang, L.; Liu, Y.-L.; Ma, C.-M.; et al. Mice Transgenic for Human Angiotensin-Converting Enzyme 2 Provide a Model for SARS Coronavirus Infection. Comp. Med. 2007, 57, 450–459. [Google Scholar]
- Zheng, J.; Wong, L.-Y.R.; Li, K.; Verma, A.K.; Ortiz, M.E.; Wohlford-Lenane, C.; Leidinger, M.R.; Knudson, C.M.; Meyerholz, D.K.; McCray, P.B.; et al. COVID-19 Treatments and Pathogenesis Including Anosmia in K18-HACE2 Mice. Nature 2021, 589, 603–607. [Google Scholar] [CrossRef]
- Lomask, M. Further Exploration of the Penh Parameter. Exp. Toxicol. Pathol. 2006, 57, 13–20. [Google Scholar] [CrossRef]
- Menachery, V.D.; Gralinski, L.E.; Baric, R.S.; Ferris, M.T. New Metrics for Evaluating Viral Respiratory Pathogenesis. PLoS ONE 2015, 10, e0131451. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 Novel Coronavirus (2019-NCoV) by Real-Time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A Mouse-Adapted Model of SARS-CoV-2 to Test COVID-19 Countermeasures. Nature 2020, 586, 560–566. [Google Scholar] [CrossRef]
- Winkler, E.S.; Bailey, A.L.; Kafai, N.M.; Nair, S.; McCune, B.T.; Yu, J.; Fox, J.M.; Chen, R.E.; Earnest, J.T.; Keeler, S.P.; et al. SARS-Cov-2 Infection of Human ACE-Transgenic Mice Causes Severe Lung Inflammation and Impaired Function. Nat. Immunol. 2020, 21, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Rheumatology High-Stakes Heterogeneity in COVID-19. Lancet Rheumatol. 2020, 2, e577. [CrossRef] [PubMed]
- Lee, K.S.; Wong, T.Y.; Russ, B.P.; Horspool, A.M.; Miller, O.A.; Rader, N.A.; Givi, J.P.; Winters, M.T.; Wong, Z.Y.A.; Cyphert, H.A.; et al. SARS-CoV-2 Delta Variant Induces Enhanced Pathology and Inflammatory Responses in K18-HACE2 Mice. PLoS ONE 2022, 17, e0273430. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mostafavi, H.; Ng, W.H.; Freitas, J.R.; King, N.J.C.; Zaid, A.; Taylor, A.; Mahalingam, S. The Delta SARS-CoV-2 Variant of Concern Induces Distinct Pathogenic Patterns of Respiratory Disease in K18-HACE2 Transgenic Mice Compared to the Ancestral Strain from Wuhan. MBio 2022, 13, e00683-22. [Google Scholar] [CrossRef] [PubMed]
- Shuai, H.; Chan, J.F.W.; Hu, B.; Chai, Y.; Yuen, T.T.T.; Yin, F.; Huang, X.; Yoon, C.; Hu, J.C.; Liu, H.; et al. Attenuated Replication and Pathogenicity of SARS-CoV-2 B.1.1.529 Omicron. Nature 2022, 603, 693–699. [Google Scholar] [CrossRef]
- Yang, J.H.; Yang, M.S.; Kim, D.M.; Kim, B.; Tark, D.; Kang, S.M.; Lee, G.H. Delta (B1.617.2) Variant of SARS-CoV-2 Induces Severe Neurotropic Patterns in K18-HACE2 Mice. Sci. Rep. 2023, 13, 3303. [Google Scholar] [CrossRef]
- Stolp, B.; Stern, M.; Ambiel, I.; Hofmann, K.; Morath, K.; Gallucci, L.; Cortese, M.; Bartenschlager, R.; Ruggieri, A.; Graw, F.; et al. SARS-CoV-2 Variants of Concern Display Enhanced Intrinsic Pathogenic Properties and Expanded Organ Tropism in Mouse Models. Cell Rep. 2022, 38, 110387. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Iida, S.; Iwatsuki-Horimoto, K.; Maemura, T.; Kiso, M.; Scheaffer, S.M.; Darling, T.L.; Joshi, A.; Loeber, S.; Singh, G.; et al. SARS-CoV-2 Omicron Virus Causes Attenuated Disease in Mice and Hamsters. Nature 2022, 603, 687–692. [Google Scholar] [CrossRef]
- Thakur, V.; Bhola, S.; Thakur, P.; Patel, S.K.S.; Kulshrestha, S.; Ratho, R.K.; Kumar, P. Waves and Variants of SARS-CoV-2: Understanding the Causes and Effect of the COVID-19 Catastrophe. Infection 2021, 50, 309–325. [Google Scholar] [CrossRef]
- Chan, J.F.W.; Zhang, A.J.; Yuan, S.; Poon, V.K.M.; Chan, C.C.S.; Lee, A.C.Y.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. 2020, 2019, 2428–2446. [Google Scholar] [CrossRef]
- Port, J.; Yinda, C.K.; Avanzato, V.; Schulz, J.; Holbrook, M.; van Doremalen, N.; Shaia, C.; Fischer, R.; Munster, V. Increased Aerosol Transmission for B.1.1.7 (Alpha Variant) over Lineage A Variant of SARS-CoV-2. Nat. Microbiol. 2022, 7, 213–223. [Google Scholar] [CrossRef]
- Sun, S.H.; Chen, Q.; Gu, H.J.; Yang, G.; Wang, Y.X.; Huang, X.Y.; Liu, S.S.; Zhang, N.N.; Li, X.F.; Xiong, R.; et al. A Mouse Model of SARS-CoV-2 Infection and Pathogenesis. Cell Host Microbe 2020, 28, 124–133.e4. [Google Scholar] [CrossRef]
- Fumagalli, V.; Ravà, M.; Marotta, D.; Di Lucia, P.; Laura, C.; Sala, E.; Grillo, M.; Bono, E.; Giustini, L.; Perucchini, C.; et al. Administration of Aerosolized SARS-CoV-2 to K18-HACE2 Mice Uncouples Respiratory Infection from Fatal Neuroinvasion. Sci. Immunol. 2022, 7, eabl9929. [Google Scholar] [CrossRef]
- Freitas, A.R.R.; Beckedorff, O.A.; Cavalcanti, L.P.d.G.; Siqueira, A.M.; de Castro, D.B.; da Costa, C.F.; Lemos, D.R.Q.; Barros, E.N.C. The Emergence of Novel SARS-CoV-2 Variant P.1 in Amazonas (Brazil) Was Temporally Associated with a Change in the Age and Sex Profile of COVID-19 Mortality: A Population Based Ecological Study. Lancet Reg. Health Am. 2021, 1, 100021. [Google Scholar] [CrossRef]
- da Silva, J.C.; Félix, V.B.; Leão, S.A.B.F.; Trindade-Filho, E.M.; Scorza, F.A. New Brazilian Variant of the SARS-CoV-2 (P1/Gamma) of COVID-19 in Alagoas State. Brazilian J. Infect. Dis. 2021, 25, 19–21. [Google Scholar] [CrossRef]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 Variant: A New Chapter in the COVID-19 Pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Ramanathan, K.; Antognini, D.; Combes, A.; Paden, M.; Zakhary, B.; Ogino, M.; MacLaren, G.; Brodie, D.; Shekar, K. Planning and Provision of ECMO Services for Severe ARDS during the COVID-19 Pandemic and Other Outbreaks of Emerging Infectious Diseases. Lancet Respir. Med. 2020, 8, 518–526. [Google Scholar] [CrossRef]
- Hu, Z.; Huang, X.; Zhang, J.; Fu, S.; Ding, D.; Tao, Z. Differences in Clinical Characteristics Between Delta Variant and Wild-Type SARS-CoV-2 Infected Patients. Front. Med. 2022, 8, 792135. [Google Scholar] [CrossRef]
- Twohig, K.A.; Nyberg, T.; Zaidi, A.; Thelwall, S.; Sinnathamby, M.A.; Aliabadi, S.; Seaman, S.R.; Harris, R.J.; Hope, R.; Lopez-Bernal, J.; et al. Hospital Admission and Emergency Care Attendance Risk for SARS-CoV-2 Delta (B.1.617.2) Compared with Alpha (B.1.1.7) Variants of Concern: A Cohort Study. Lancet Infect. Dis. 2022, 22, 35–42. [Google Scholar] [CrossRef]
- Meyerowitz, E.A.; Richterman, A. SARS-CoV-2 Transmission and Prevention in the Era of the Delta Variant. Infect. Dis. Clin. North Am. 2022, 36, 267–293. [Google Scholar] [CrossRef]
- Golden, J.W.; Cline, C.R.; Zeng, X.; Garrison, A.R.; Carey, B.D.; Mucker, E.M.; White, L.E.; Shamblin, J.D.; Brocato, R.L.; Liu, J.; et al. Human Angiotensin-Converting Enzyme 2 Transgenic Mice Infected with SARS-CoV-2 Develop Severe and Fatal Respiratory Disease. JCI Insight 2020, 5, e142032. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yan, L.M.; Wan, L.; Xiang, T.X.; Le, A.; Liu, J.M.; Peiris, M.; Poon, L.L.M.; Zhang, W. Viral Dynamics in Mild and Severe Cases of COVID-19. Lancet Infect. Dis. 2020, 20, 656–657. [Google Scholar] [CrossRef]
- Spinelli, M.A.; Rutherford, G.; Gandhi, M. Lowering SARS-CoV-2 Viral Load Might Affect Transmission but Not Disease Severity in Secondary Cases—Authors’ Reply. Lancet Infect. Dis. 2021, 21, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Deftereos, S.G.; Siasos, G.; Giannopoulos, G.; Vrachatis, D.A.; Angelidis, C.; Giotaki, S.G.; Gargalianos, P.; Giamarellou, H.; Gogos, C.; Daikos, G.; et al. The Greek Study in the Effects of Colchicine in COvid-19 Complications Prevention (GRECCO-19 Study): Rationale and Study Design. Hell. J. Cardiol. 2020, 61, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Kumar, M.; Sharma, P.; Mohan, T.; Prakash, A.; Kumari, R.; Kaur, P. Effect of Double Mutation (L452R and E484Q) on the Binding Affinity of Monoclonal Antibodies (MAbs) against the RBD—A Target for Vaccine Development. Vaccines 2022, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, M.; Tripathi, M.K.; Gupta, D.; Vishwakarma, P.; Das, U.; Kaur, P. Genomic and Structural Mechanistic Insight to Reveal the Differential Infectivity of Omicron and Other Variants of Concern. Comput. Biol. Med. 2022, 150, 106129. [Google Scholar] [CrossRef]
- Nealon, J.; Cowling, B.J. Omicron Severity: Milder but Not Mild. Lancet 2022, 399, 412–413. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva Santos, Y.; Gamon, T.H.M.; de Azevedo, M.S.P.; Telezynski, B.L.; de Souza, E.E.; de Oliveira, D.B.L.; Dombrowski, J.G.; Rosa-Fernandes, L.; Palmisano, G.; de Moura Carvalho, L.J.; et al. Virulence Profiles of Wild-Type, P.1 and Delta SARS-CoV-2 Variants in K18-hACE2 Transgenic Mice. Viruses 2023, 15, 999. https://doi.org/10.3390/v15040999
da Silva Santos Y, Gamon THM, de Azevedo MSP, Telezynski BL, de Souza EE, de Oliveira DBL, Dombrowski JG, Rosa-Fernandes L, Palmisano G, de Moura Carvalho LJ, et al. Virulence Profiles of Wild-Type, P.1 and Delta SARS-CoV-2 Variants in K18-hACE2 Transgenic Mice. Viruses. 2023; 15(4):999. https://doi.org/10.3390/v15040999
Chicago/Turabian Styleda Silva Santos, Yasmin, Thais Helena Martins Gamon, Marcela Santiago Pacheco de Azevedo, Bruna Larotonda Telezynski, Edmarcia Elisa de Souza, Danielle Bruna Leal de Oliveira, Jamille Gregório Dombrowski, Livia Rosa-Fernandes, Giuseppe Palmisano, Leonardo José de Moura Carvalho, and et al. 2023. "Virulence Profiles of Wild-Type, P.1 and Delta SARS-CoV-2 Variants in K18-hACE2 Transgenic Mice" Viruses 15, no. 4: 999. https://doi.org/10.3390/v15040999