
Human SCARB2 Acts as a Cellular Associator for Helping Coxsackieviruses A10 Infection
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells and Viruses
2.3. Antibodies and Western Blotting
2.4. Co-Immunoprecipitation
2.5. Reduction of Cellular Gene Expression by siRNA
2.6. CVA10 Infection in hSCARB2-Tg Mice
2.7. Real-Time RT-PCR
2.8. ELISA
2.9. Virus Neutralization Assay
2.10. Enzyme-Linked Immunosorbent Spot Assay
2.11. Statistical Analysis
3. Results
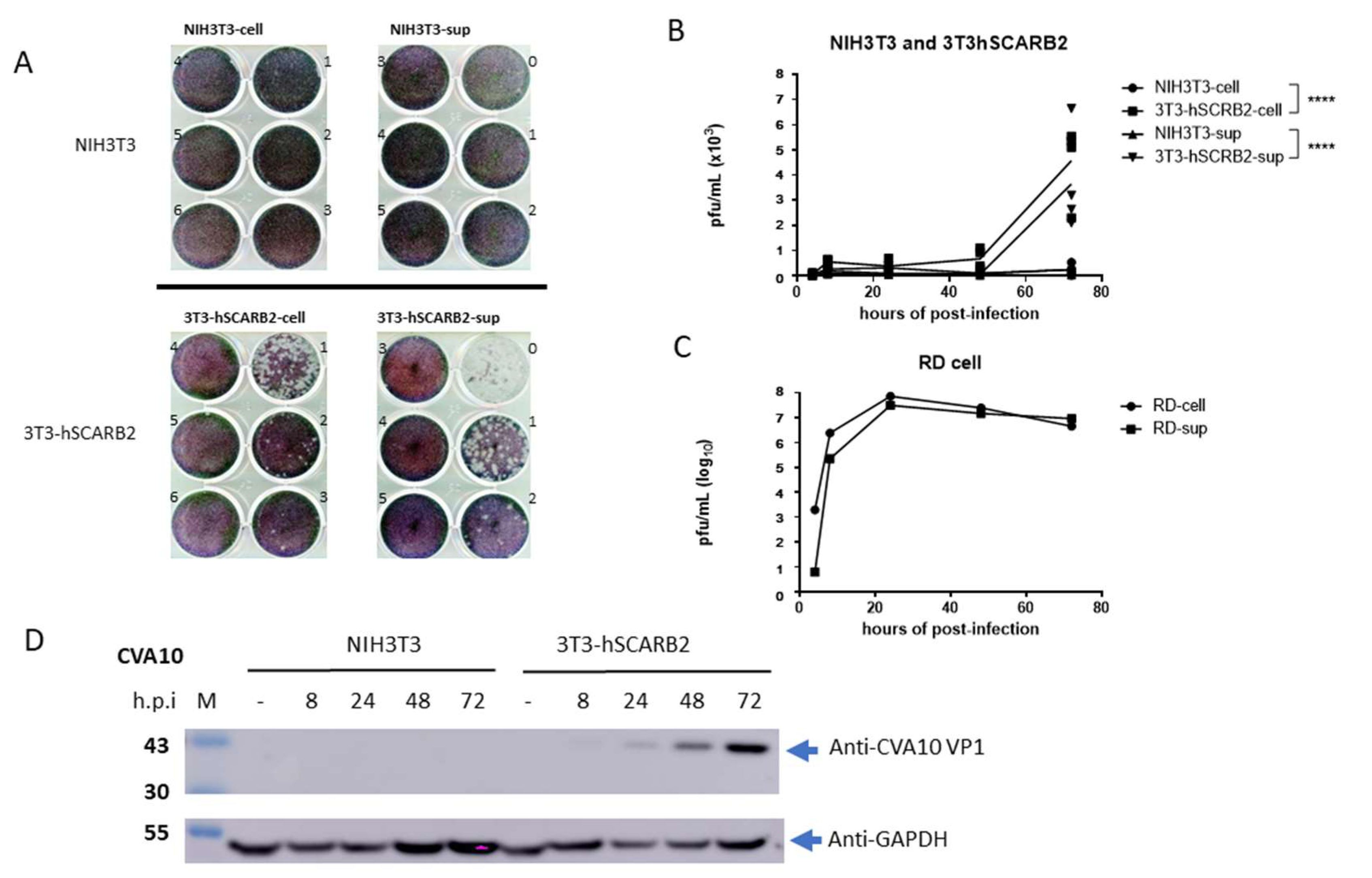
3.1. hSCARB2 Expression in 3T3-SCARB2 Facilitates CVA10 Infection
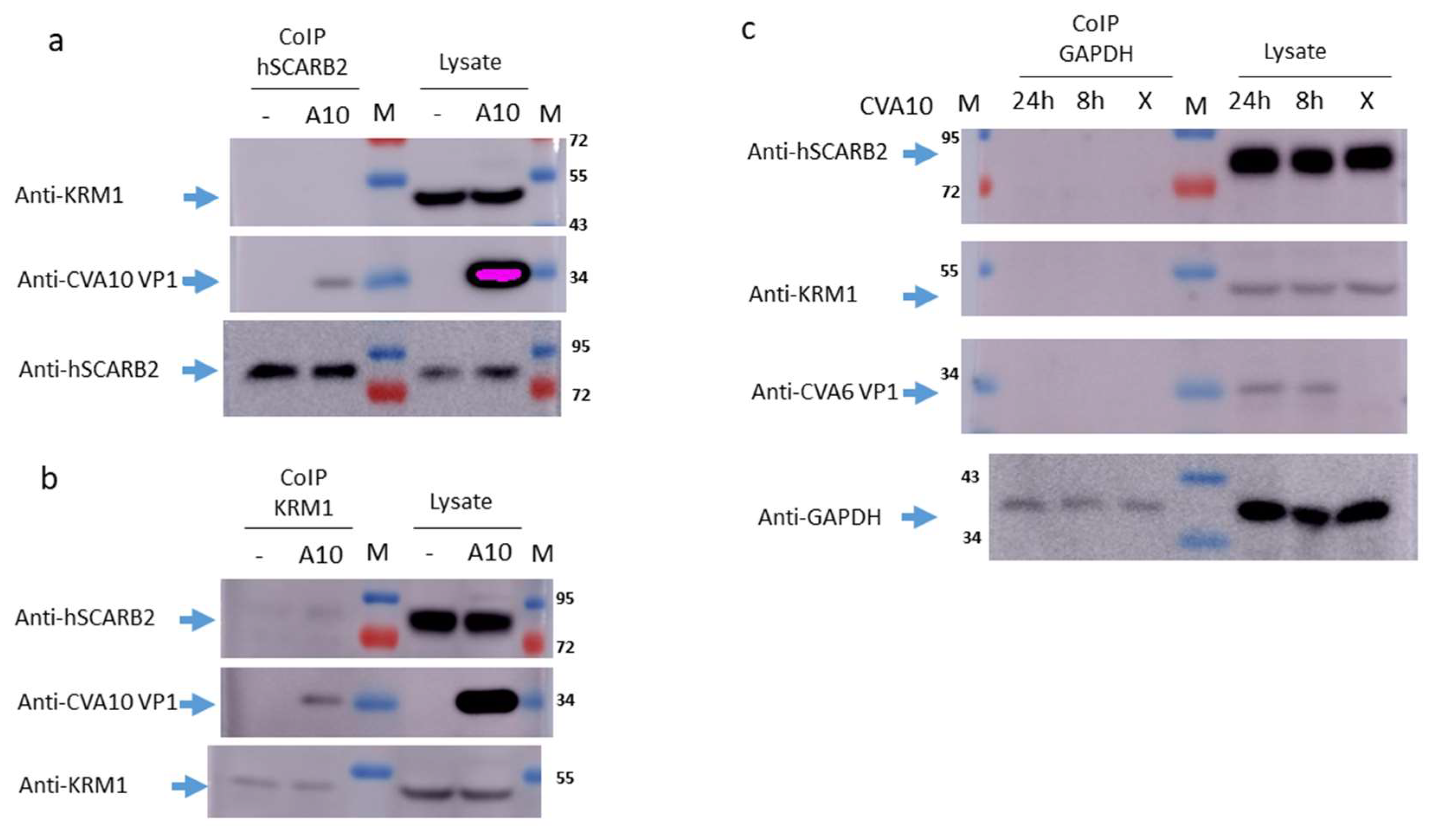
3.2. Interaction of CVA10 with hSCARB2 and KRM1
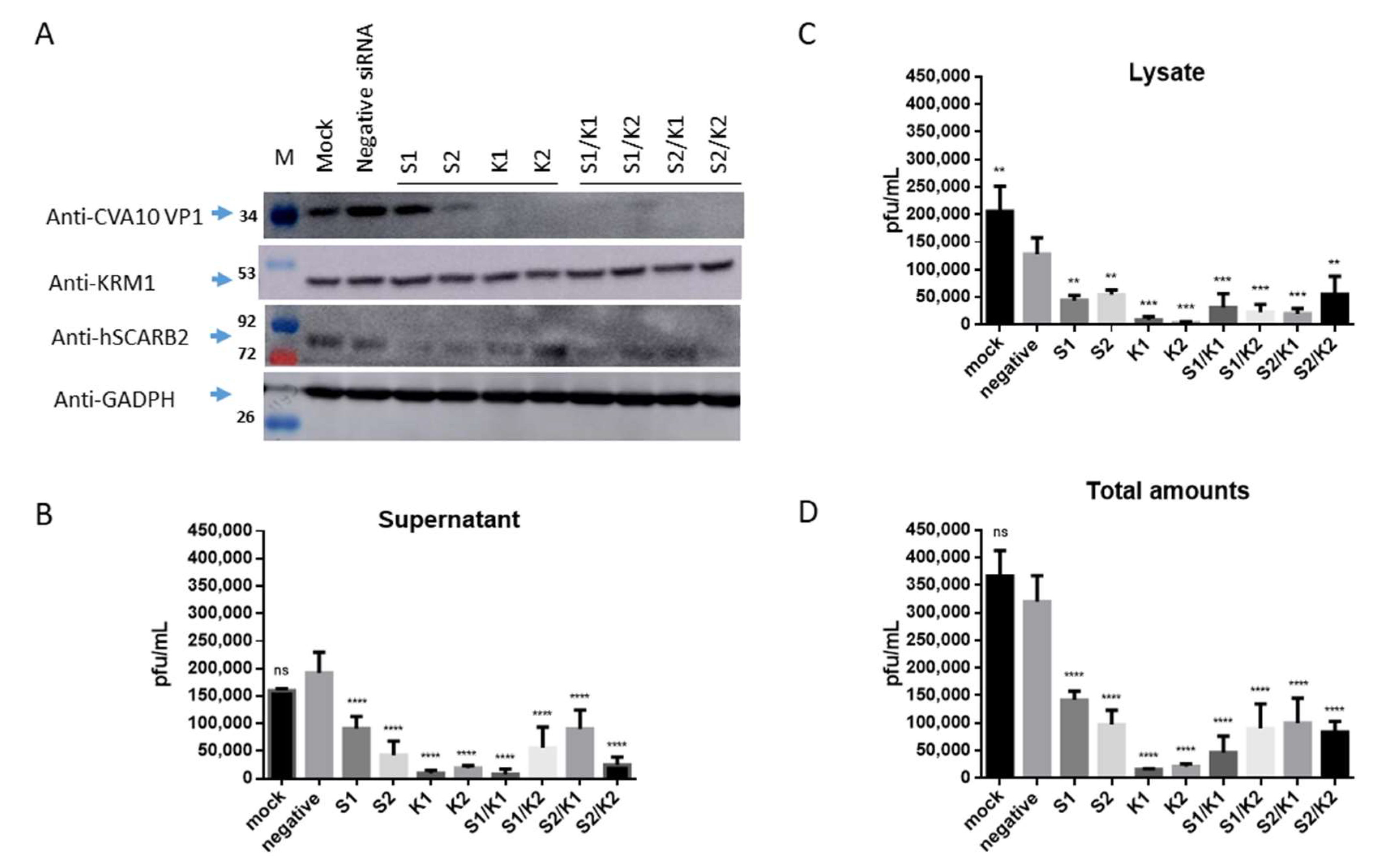
3.3. Inhibition of CVA10 Infection by Knocking down hSCARB2 Expression
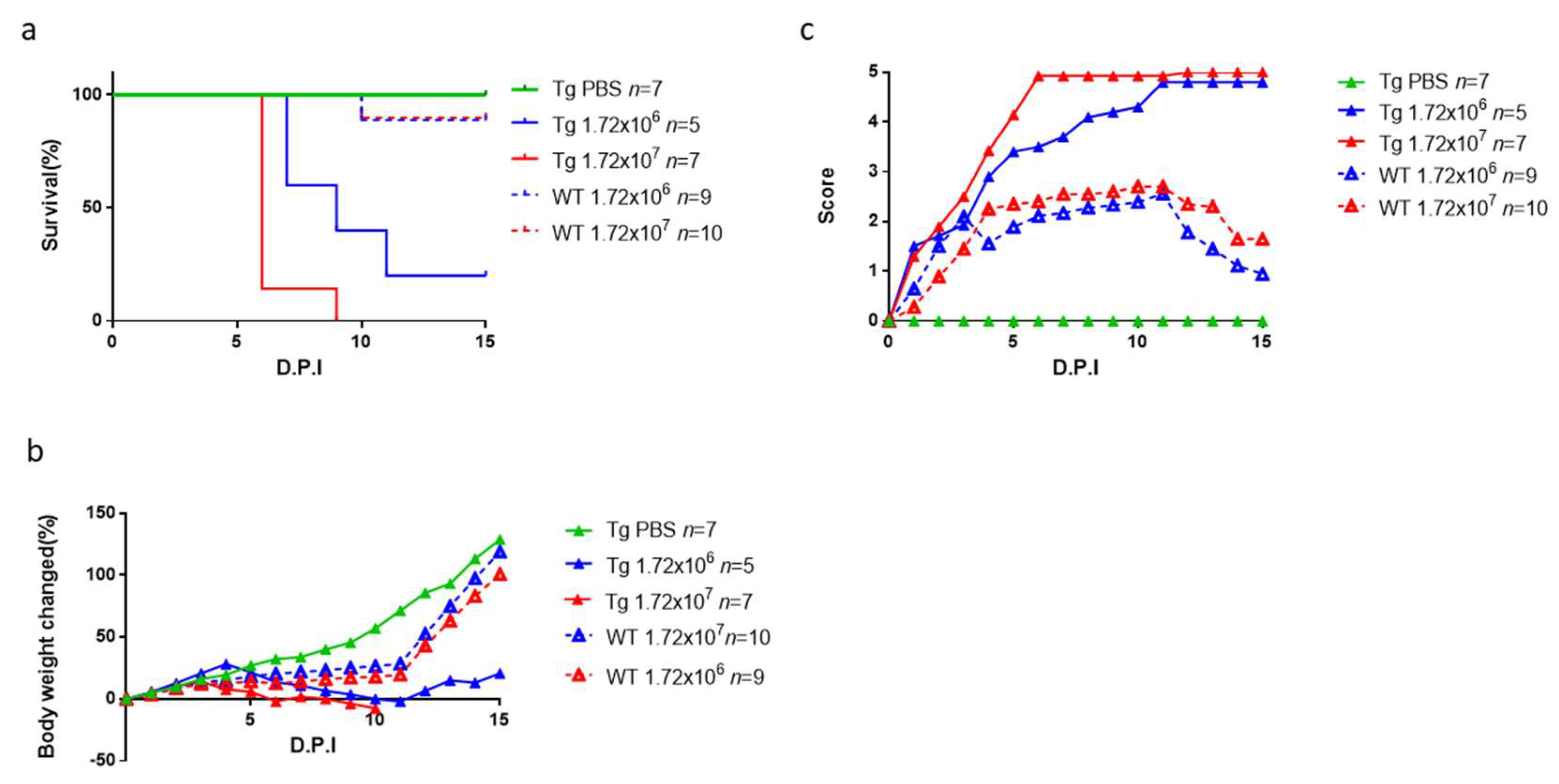
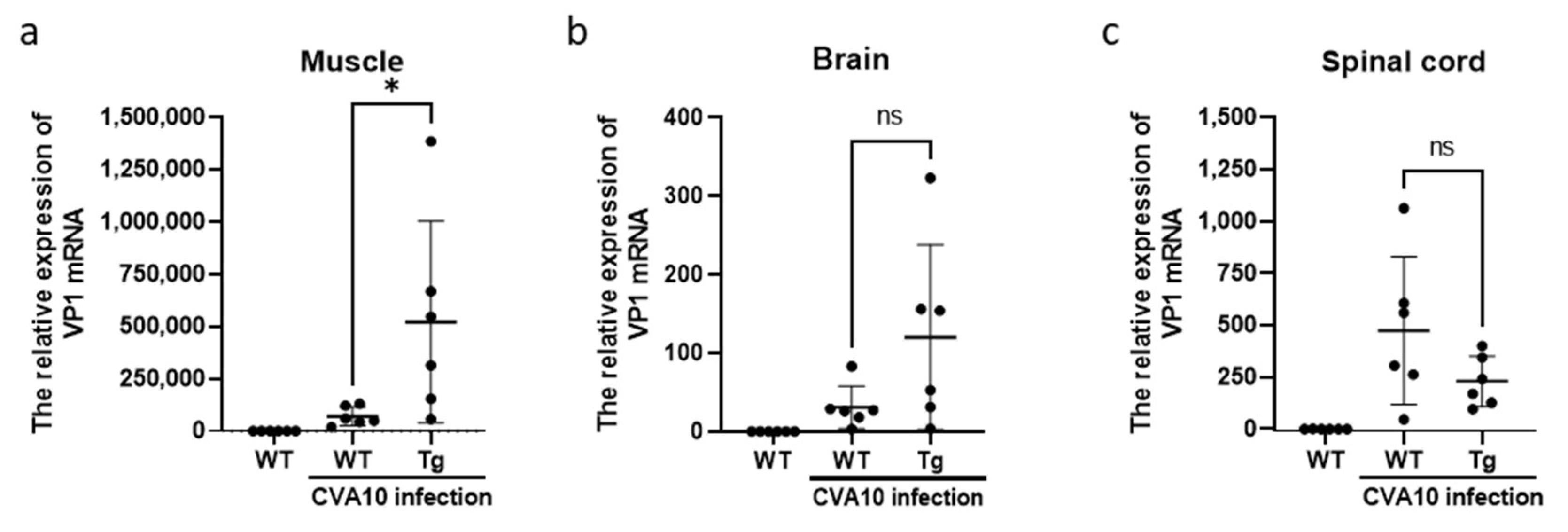
3.4. CVA10 Induces a Severe Neurological Disease in hSCARB2-Tg Mice
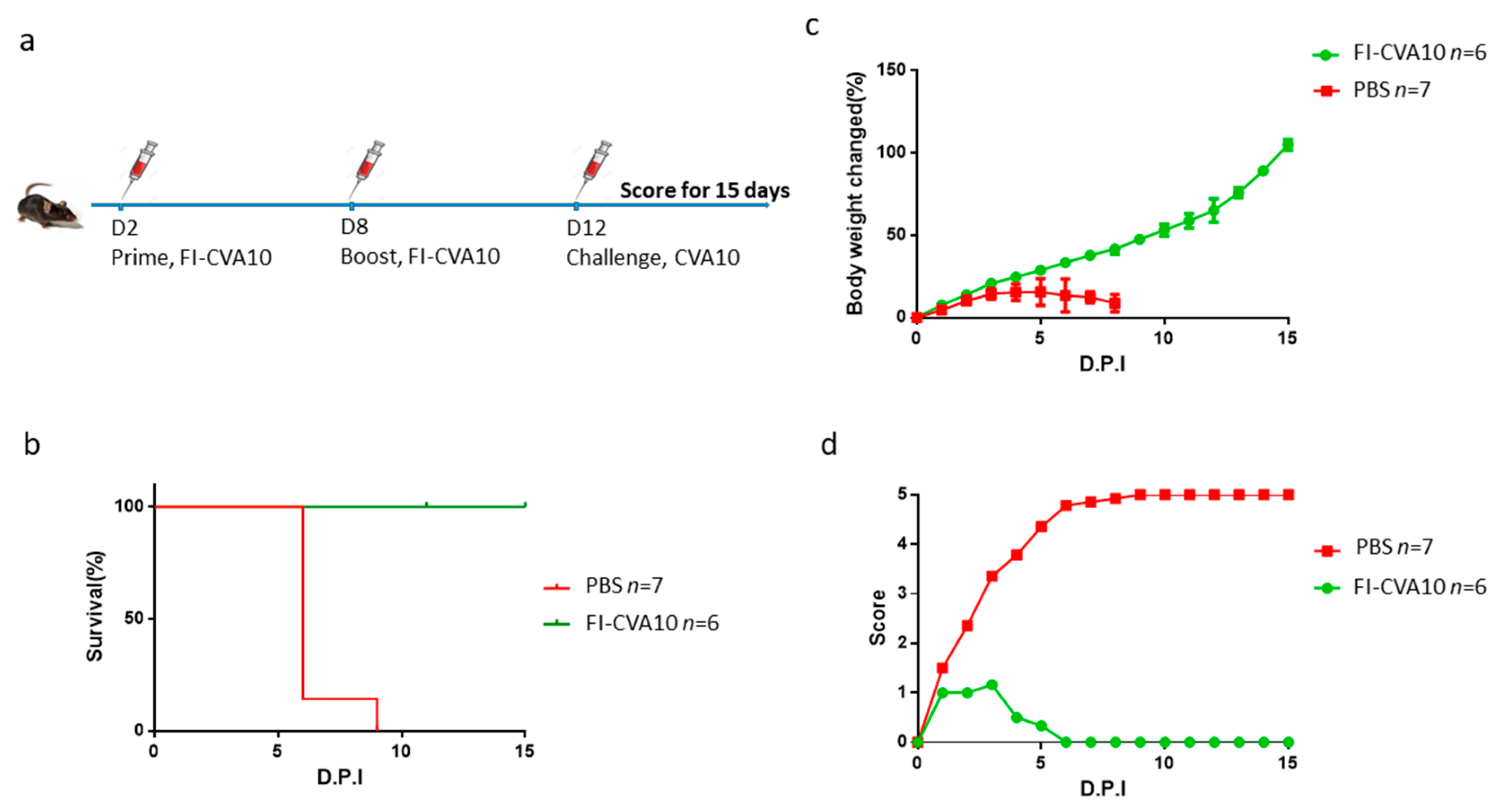
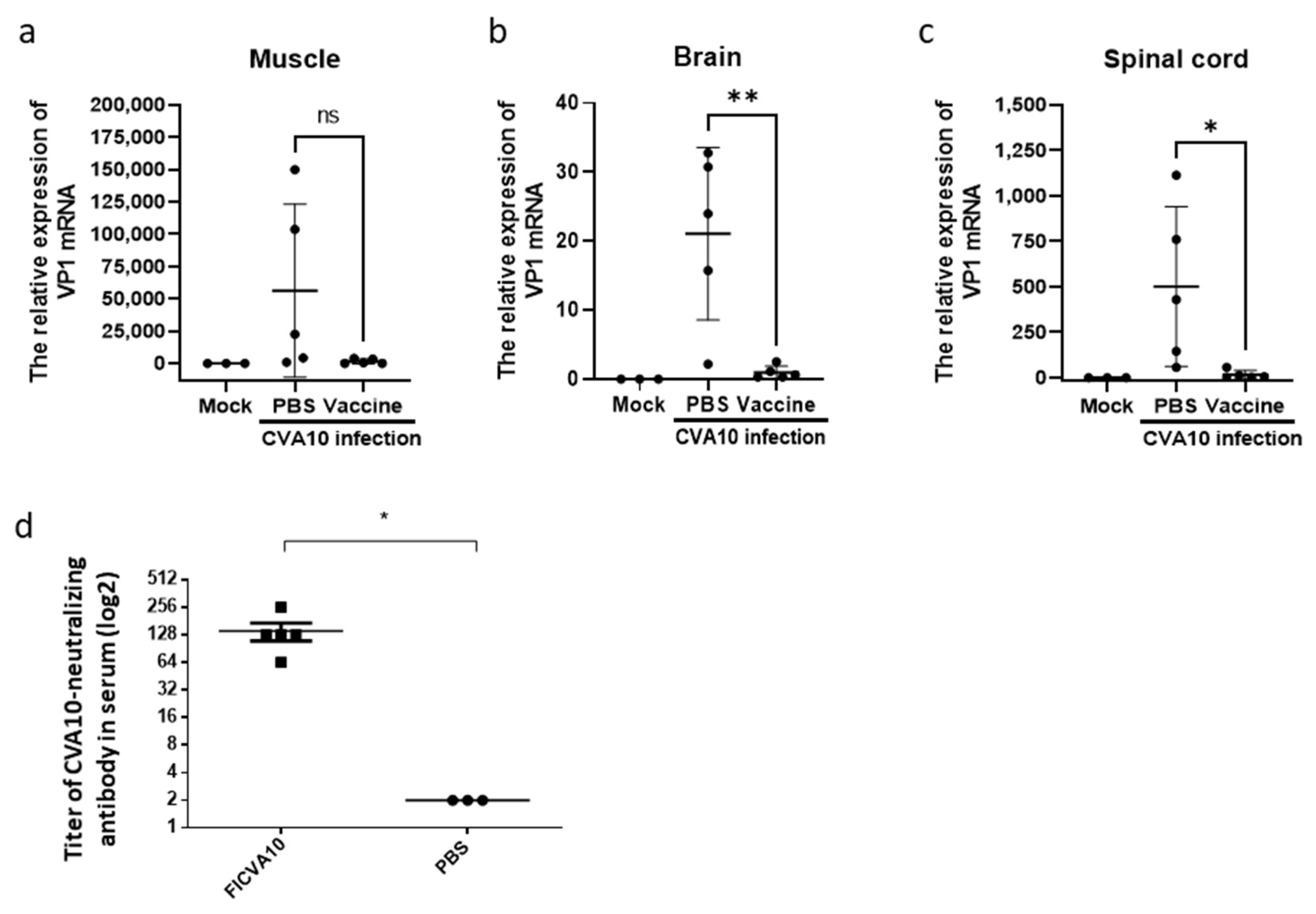
3.5. Immunization of Anti-CVA10 Vaccine Protects Mice from Live CVA10 Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Itagaki, A.; Kamahora, T.; Kurimura, T. Isolation and characterization of a cold-sensitive strain of coxsackievirus A10. J. Gen. Virol. 1987, 68 Pt 4, 1191–1194. [Google Scholar] [CrossRef]
- Hoa-Tran, T.N.; Nguyen, A.T.; Dao, A.T.H.; Kataoka, C.; Ta, H.T.T.; Nguyen, H.T.V.; Takemura, T.; Nguyen, T.T.T.; Vu, H.M.; Nguyen, T.T.H.; et al. Genetic characterization of VP1 of coxsackieviruses A2, A4, and A10 associated with hand, foot, and mouth disease in Vietnam in 2012-2017: Endemic circulation and emergence of new HFMD-causing lineages. Arch. Virol. 2020, 165, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Mirand, A.; Henquell, C.; Archimbaud, C.; Ughetto, S.; Antona, D.; Bailly, J.L.; Peigue-Lafeuille, H. Outbreak of hand, foot and mouth disease/herpangina associated with coxsackievirus A6 and A10 infections in 2010, France: A large citywide, prospective observational study. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2012, 18, E110–E118. [Google Scholar]
- Lizasoain, A.; Mir, D.; Martinez, N.; Colina, R. Coxsackievirus A10 causing hand-foot-and-mouth disease in Uruguay. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2020, 94, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; He, S.; Yan, Q.; Xu, X.; Wu, W.; Ge, S.; Zhang, S.; Chen, M.; Xia, N. Severe hand, foot and mouth disease associated with Coxsackievirus A10 infections in Xiamen, China in 2015. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2017, 93, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Gao, F.; Mao, Q.; Sun, S.; Wu, X.; Liu, S.; Yang, X.; Liang, Z. Hand, foot, and mouth disease associated with coxsackievirus A10: More serious than it seems. Expert Rev. Anti-Infect. Ther. 2019, 17, 233–242. [Google Scholar]
- Fuschino, M.E.; Lamson, D.M.; Rush, K.; Carbone, L.S.; Taff, M.L.; Hua, Z.; Landi, K.; George, K.S. Detection of coxsackievirus A10 in multiple tissues of a fatal infant sepsis case. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2012, 53, 259–261. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, D.; Yan, D.; Zhu, S.; Liu, J.; Wang, H.; Zhao, S.; Yu, D.; Nan, L.; An, J.; et al. Molecular evidence of persistent epidemic and evolution of subgenotype B1 coxsackievirus A16-associated hand, foot, and mouth disease in China. J. Clin. Microbiol. 2010, 48, 619–622. [Google Scholar] [CrossRef]
- Munivenkatappa, A.; Yadav, P.D.; Nyayanit, D.A.; Majumdar, T.D.; Sangal, L.; Jain, S.; Sinha, D.P.; Shrivastava, A.; Mourya, D.T. Molecular diversity of Coxsackievirus A10 circulating in the southern and northern region of India [2009-17]. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 66, 101–110. [Google Scholar]
- Zhang, Z.; Dong, Z.; Li, J.; Carr, M.J.; Zhuang, D.; Wang, J.; Zhang, Y.; Ding, S.; Tong, Y.; Li, D.; et al. Protective Efficacies of Formaldehyde-Inactivated Whole-Virus Vaccine and Antivirals in a Murine Model of Coxsackievirus A10 Infection. J. Virol. 2017, 91, e00333-17. [Google Scholar]
- Li, S.; Zhao, H.; Yang, L.; Hou, W.; Xu, L.; Wu, Y.; Wang, W.; Chen, C.; Wan, J.; Ye, X.; et al. A neonatal mouse model of coxsackievirus A10 infection for anti-viral evaluation. Antivir. Res. 2017, 144, 247–255. [Google Scholar]
- Gao, W.; Yue, L.; Yang, T.; Shen, D.; Li, H.; Song, X.; Xie, T.; He, X.; Xie, Z. Proliferation characteristics of coxsackievirus A10 in mice and immune protection ability of experimental inactivated vaccine. Biomed. Pharmacother. 2021, 143, 112212. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, C.; Liu, Q.; Gong, S.; Geng, L.; Huang, Z. A virus-like particle vaccine protects mice against coxsackievirus A10 lethal infection. Antivir. Res. 2018, 152, 124–130. [Google Scholar] [PubMed]
- Zhang, Z.; Dong, Z.; Wang, Q.; Carr, M.J.; Li, J.; Liu, T.; Li, D.; Shi, W. Characterization of an inactivated whole-virus bivalent vaccine that induces balanced protective immunity against coxsackievirus A6 and A10 in mice. Vaccine 2018, 36, 7095–7104. [Google Scholar]
- Chen, C.; Xia, Y.; Zhu, S.; Xu, F.; Sun, Y.; Lu, H.; Gao, M.; Yang, Z.; Mao, Z.; Ge, Q.; et al. Muscle destruction caused by coxsackievirus A10 in gerbils: Construction of a novel animal model for antiviral evaluation. Virus Res. 2020, 286, 198067. [Google Scholar] [CrossRef]
- Shen, C.; Liu, Q.; Zhou, Y.; Ku, Z.; Wang, L.; Lan, K.; Ye, X.; Huang, Z. Inactivated coxsackievirus A10 experimental vaccines protect mice against lethal viral challenge. Vaccine 2016, 34, 5005–5012. [Google Scholar] [CrossRef]
- An, H.H.; Li, M.; Liu, R.L.; Wu, J.; Meng, S.L.; Guo, J.; Wang, Z.J.; Qian, S.S.; Shen, S. Humoral and cellular immunogenicity and efficacy of a coxsackievirus A10 vaccine in mice. Emerg. Microbes Infect. 2023, 12, e2147022. [Google Scholar] [CrossRef]
- Zhu, L.; Sun, Y.; Fan, J.; Zhu, B.; Cao, L.; Gao, Q.; Zhang, Y.; Liu, H.; Rao, Z.; Wang, X. Structures of Coxsackievirus A10 unveil the molecular mechanisms of receptor binding and viral uncoating. Nat. Commun. 2018, 9, 4985. [Google Scholar] [CrossRef] [PubMed]
- Staring, J.; van den Hengel, L.G.; Raaben, M.; Blomen, V.A.; Carette, J.E.; Brummelkamp, T.R. KREMEN1 Is a Host Entry Receptor for a Major Group of Enteroviruses. Cell Host Microbe 2018, 23, 636–643.e5. [Google Scholar] [CrossRef]
- Mao, B.; Wu, W.; Davidson, G.; Marhold, J.; Li, M.; Mechler, B.M.; Delius, H.; Hoppe, D.; Stannek, P.; Walter, C.; et al. Kremen proteins are Dickkopf receptors that regulate Wnt/beta-catenin signalling. Nature 2002, 417, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Shimizu, H. Cellular receptors for human enterovirus species a. Front. Microbiol. 2012, 3, 105. [Google Scholar] [CrossRef]
- de Villiers, W.J.; Smart, E.J. Macrophage scavenger receptors and foam cell formation. J. Leukoc. Biol. 1999, 66, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, E.L.; Tanaka, Y.; Saftig, P. At the acidic edge: Emerging functions for lysosomal membrane proteins. Trends Cell Biol. 2003, 13, 137–145. [Google Scholar] [PubMed]
- Yamayoshi, S.; Yamashita, Y.; Li, J.; Hanagata, N.; Minowa, T.; Takemura, T.; Koike, S. Scavenger receptor B2 is a cellular receptor for enterovirus 71. Nat. Med. 2009, 15, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Yu, S.L.; Shao, H.Y.; Lin, H.Y.; Liu, C.C.; Hsiao, K.N.; Chitra, E.; Tsou, Y.L.; Chang, H.W.; Sia, C.; et al. Human SCARB2 Transgenic Mice as an Infectious Animal Model for Enterovirus 71. PLoS ONE 2013, 8, e57591. [Google Scholar]
- Lin, Y.W.; Lin, H.Y.; Tsou, Y.L.; Chitra, E.; Hsiao, K.N.; Shao, H.Y.; Liu, C.C.; Sia, C.; Chong, P.; Chow, Y.H. Human SCARB2-mediated entry and endocytosis of EV71. PLoS ONE 2012, 7, e30507. [Google Scholar]
- Liu, C.C.; Chou, A.H.; Lien, S.P.; Lin, H.Y.; Liu, S.J.; Chang, J.Y.; Guo, M.S.; Chow, Y.H.; Yang, W.S.; Chang, K.H.; et al. Identification and characterization of a cross-neutralization epitope of Enterovirus 71. Vaccine 2011, 29, 4362–4372. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dun, Y.; Yan, J.; Wang, M.; Wang, M.; Liu, L.; Yu, R.; Zhang, S. Rac1-dependent endocytosis and Rab5-dependent intracellular trafficking are required by Enterovirus A71 and Coxsackievirus A10 to establish infections. Biochem. Biophys. Res. Commun. 2020, 529, 97–103. [Google Scholar]
- Sanchez, E.G.; Perez-Nunez, D.; Revilla, Y. Mechanisms of Entry and Endosomal Pathway of African Swine Fever Virus. Vaccines 2017, 5, 42. [Google Scholar] [CrossRef]
- Lai, R.H.; Chow, Y.H.; Chung, N.H.; Chen, T.C.; Shie, F.S.; Juang, J.L. Neurotropic EV71 causes encephalitis by engaging intracellular TLR9 to elicit neurotoxic IL12-p40-iNOS signaling. Cell Death Dis. 2022, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Lin, Y.W.; Ho, H.M.; Lin, M.H.; Liu, C.C.; Shao, H.Y.; Chong, P.; Sia, C.; Chow, Y.H. Protective efficacy of VP1-specific neutralizing antibody associated with a reduction of viral load and pro-inflammatory cytokines in human SCARB2-transgenic mice. PLoS ONE 2013, 8, e69858. [Google Scholar] [CrossRef]
- Tsou, Y.L.; Lin, Y.W.; Shao, H.Y.; Yu, S.L.; Wu, S.R.; Lin, H.Y.; Liu, C.C.; Huang, C.; Chong, P.; Chow, Y.H. Recombinant adeno-vaccine expressing enterovirus 71-like particles against hand, foot, and mouth disease. PLoS Negl. Trop. Dis. 2015, 9, e0003692. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.; Cai, J.; Ge, Y.; Wang, C.; Qiu, Y.; Xia, A.; Zeng, M. Non-polio enterovirus infections in children with central nervous system disorders in Shanghai, 2016-2018: Serotypes and clinical characteristics. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2020, 129, 104516. [Google Scholar] [CrossRef]
- Gonzalez, G.; Carr, M.J.; Kobayashi, M.; Hanaoka, N.; Fujimoto, T. Enterovirus-Associated Hand-Foot and Mouth Disease and Neurological Complications in Japan and the Rest of the World. Int. J. Mol. Sci. 2019, 20, 5201. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Target Gene | Sequences (5′->3′) of siRNAs |
|---|---|---|
| CVA10-F | CVA10 VP1 | CGGTCCCTTTCATGTCACCA |
| CVA10-R | ACTCTCACTGCAAAGGTGCC | |
| Probe of CVA10-P | CVA10 VP1 | HEX-AAACTCACTGACCCTCCTGCACAAGTCTCA-BHQ1 |
| β-actin-F | Mouse β-actin | ACCAACTGGGACGACATGGAGAAA |
| β-actin-R | TAGCACAGCCTGGATAGCAACGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-L.; Chung, N.-H.; Lin, Y.-C.; Liao, Y.-A.; Chen, Y.-C.; Chow, Y.-H. Human SCARB2 Acts as a Cellular Associator for Helping Coxsackieviruses A10 Infection. Viruses 2023, 15, 932. https://doi.org/10.3390/v15040932
Yu S-L, Chung N-H, Lin Y-C, Liao Y-A, Chen Y-C, Chow Y-H. Human SCARB2 Acts as a Cellular Associator for Helping Coxsackieviruses A10 Infection. Viruses. 2023; 15(4):932. https://doi.org/10.3390/v15040932
Chicago/Turabian StyleYu, Shu-Ling, Nai-Hsiang Chung, Yu-Ching Lin, Yi-An Liao, Ying-Chin Chen, and Yen-Hung Chow. 2023. "Human SCARB2 Acts as a Cellular Associator for Helping Coxsackieviruses A10 Infection" Viruses 15, no. 4: 932. https://doi.org/10.3390/v15040932