
Differential Effects of RNA-Dependent RNA Polymerase 6 (RDR6) Silencing on New and Old World Begomoviruses in Nicotiana benthamiana
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Virus Inoculation
2.2. Symptom Analysis
2.3. Total Nucleic Acid (TNA) Extraction and Southern Blot Analysis
2.4. In Situ Hybridization (ISH)
2.5. Viral siRNA Detection by Northern Blot Analysis
2.6. Statistical Analysis
3. Results and Discussion
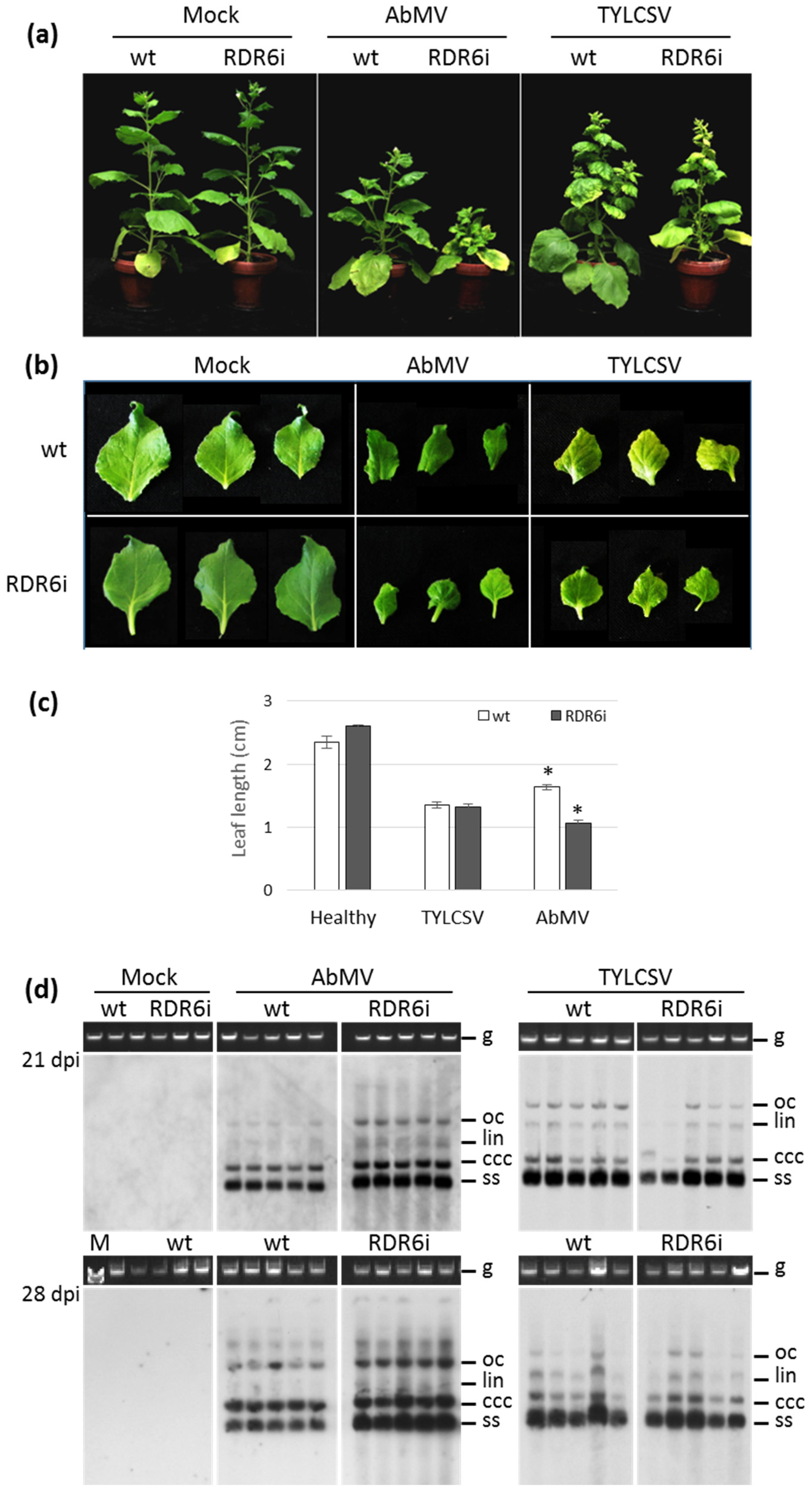
3.1. Silencing of NbRDR6 Has a Differential Effect on the Symptoms Induced by AbMV and TYLCSV and on Their Genome Accumulation

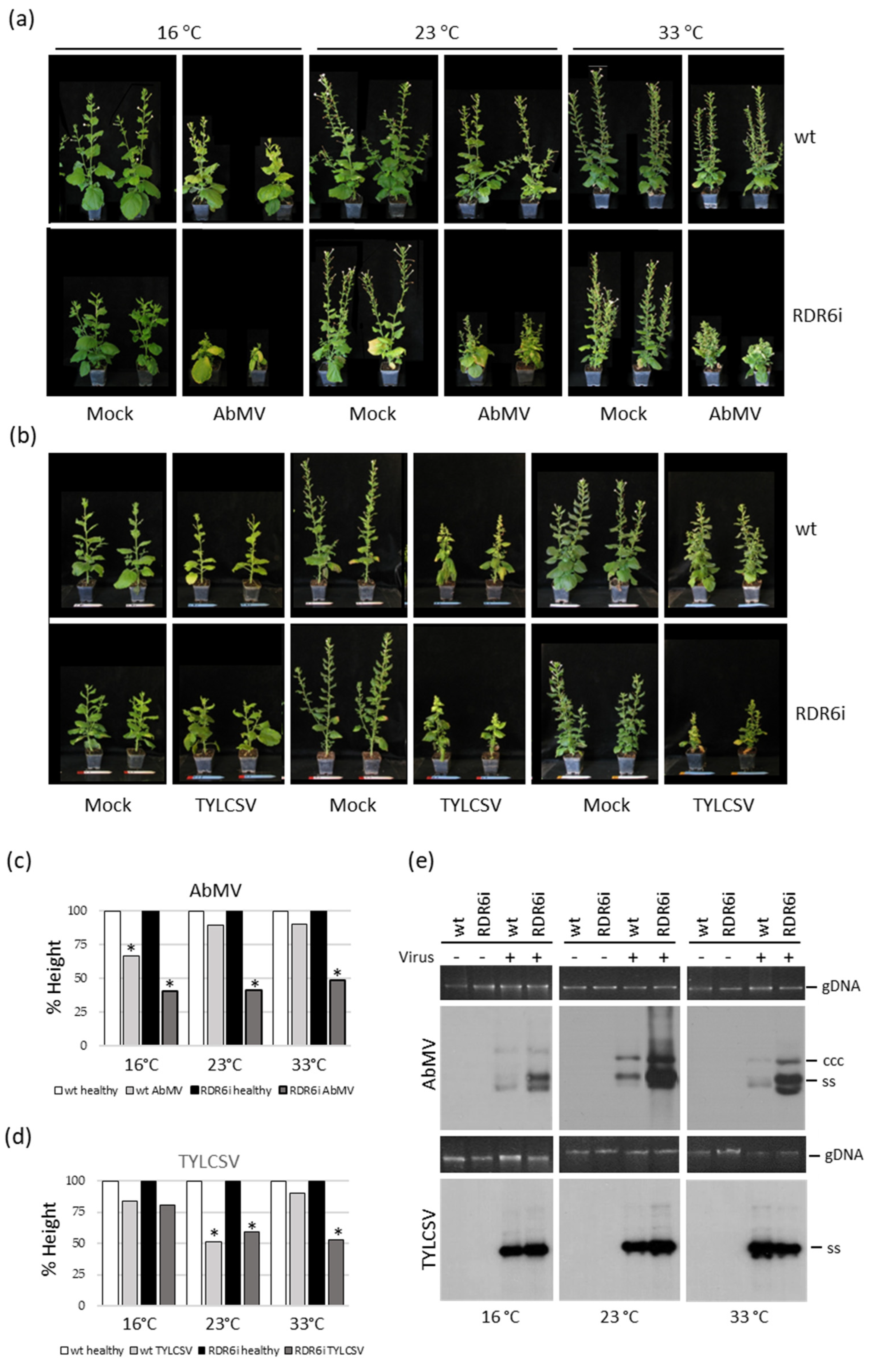
3.2. Plant Growth Temperature Influences the Response of RDR6i Plants to AbMV but Not to TYLCSV Infection

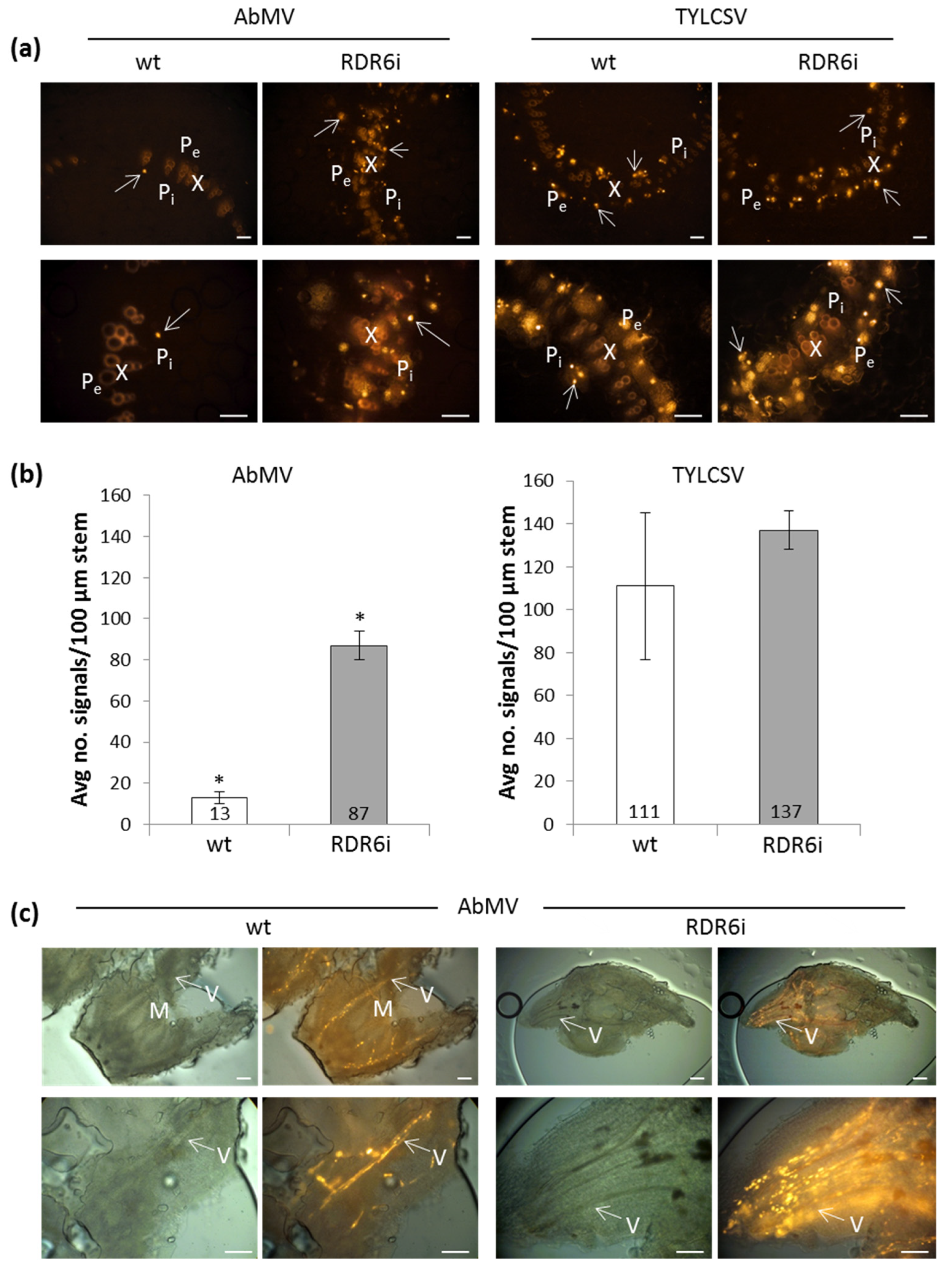
3.3. In Situ Hybridization (ISH) Reveals Increased Numbers of Infected Nuclei in RDR6i Plants with AbMV but Not TYLCSV
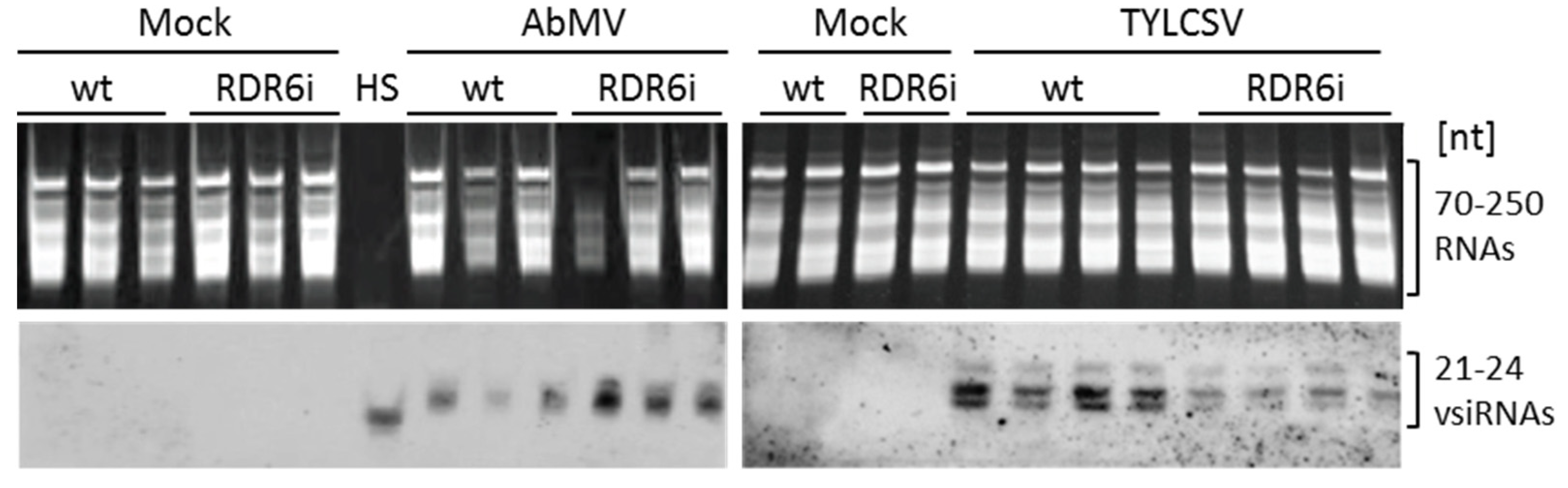
3.4. Depletion of NbRDR6 Gene Expression Modulates the Amount of Virus-Derived siRNAs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Rivera Bustamante, R.F.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef]
- Roumagnac, P.; Lett, J.M.; Fiallo-Olivé, E.; Navas-Castillo, J.; Zerbini, F.M.; Martin, D.P.; Varsani, A. Establishment of five new genera in the family Geminiviridae: Citlodavirus, Maldovirus, Mulcrilevirus, Opunvirus, and Topilevirus. Arch. Virol. 2022, 167, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV virus taxonomy profile. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef]
- Nigam, D. Genomic variation and diversification in Begomovirus genome in implication to host and vector adaptation. Plants 2021, 10, 1706. [Google Scholar] [CrossRef]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef]
- Verdel, A.; Vavasseur, A.; Le Gorrec, M.; Touat-Todeschini, L. Common themes in siRNA-mediated epigenetic silencing pathways. Int. J. Dev. Biol. 2009, 53, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Gomollon, S.; Baulcombe, D.C. Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems. Nat. Rev. Mol. Cell Biol. 2022, 23, 645–662. [Google Scholar] [CrossRef]
- Teixeira, R.M.; Ferreira, M.A.; Raimundo, G.A.S.; Fontes, E.P.B. Geminiviral triggers and suppressors of plant antiviral immunity. Microorganisms 2021, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Akbergenov, R.; Si-Ammour, A.; Blevins, T.; Amin, I.; Kutter, C.; Vanderschuren, H.; Zhang, P.; Gruissem, W.; Meins, F.; Hohn, T.; et al. Molecular characterization of geminivirus-derived small RNAs in different plant species. Nucleic Acids Res. 2006, 34, 462–471. [Google Scholar] [CrossRef]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.S.; Vazquez, F.; Robertson, D.; Meins, F.; Hohn, T.; et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing. Nucleic Acids Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef] [Green Version]
- Blevins, T.; Rajeswaran, R.; Aregger, M.; Borah, B.K.; Schepetilnikov, M.; Baerlocher, L.; Farinelli, L.; Meins, F.; Hohn, T.; Pooggin, M.M. Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense. Nucleic Acids Res. 2011, 39, 5003–5014. [Google Scholar] [CrossRef] [Green Version]
- Miozzi, L.; Pantaleo, V.; Burgyán, J.; Accotto, G.P.; Noris, E. Analysis of small RNAs derived from tomato yellow leaf curl Sardinia virus reveals a cross reaction between the major viral hotspot and the plant host genome. Virus Res. 2013, 178, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Aregger, M.; Borah, B.K.; Seguin, J.; Rajeswaran, R.; Gubaeva, E.G.; Zvereva, A.S.; Windels, D.; Vazquez, F.; Blevins, T.; Farinelli, L.; et al. Primary and secondary siRNAs in geminivirus-induced gene silencing. PLoS Pathog. 2012, 8, e1002941. [Google Scholar] [CrossRef]
- Piedra-Aguilera, Á.; Jiao, C.; Luna, A.P.; Villanueva, F.; Dabad, M.; Esteve-Codina, A.; Díaz-Pendón, J.A.; Fei, Z.; Bejarano, E.R.; Castillo, A.G. Integrated single-base resolution maps of transcriptome, sRNAome and methylome of Tomato yellow leaf curl virus (TYLCV) in tomato. Sci. Rep. 2019, 9, 2863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaistij, F.E.; Jones, L.; Baulcombe, D.C. Spreading of RNA targeting and DNA methylation in RNA silencing requires transcription of the target gene and a putative RNA-dependent RNA polymerase. Plant Cell 2002, 14, 857–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwach, F.; Vaistij, F.E.; Jones, L.; Baulcombe, D.C. An RNA-dependent RNA polymerase prevents meristem invasion by potato virus X and is required for the activity but not the production of a systemic silencing signal. Plant Physiol. 2005, 138, 1842–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Wassenegger, M.; Krczal, G. Nomenclature and functions of RNA-directed RNA polymerases. Trends Plant Sci. 2006, 11, 142–151. [Google Scholar] [CrossRef]
- Dunoyer, P.; Voinnet, O. Movement of RNA silencing between plant cells: Is the question now behind us? Trends Plant Sci. 2009, 14, 643–644. [Google Scholar] [CrossRef]
- Matzke, M.A.; Kanno, T.; Matzke, A.J. RNA-Directed DNA Methylation: The evolution of a complex epigenetic pathway in flowering plants. Annu. Rev. Plant Biol. 2015, 66, 243–267. [Google Scholar] [CrossRef]
- Hua, X.; Berkowitz, N.D.; Willmann, M.R.; Yu, X.; Lyons, E.; Gregory, B.D. Global Analysis of RNA-Dependent RNA Polymerase-Dependent Small RNAs Reveals New Substrates and Functions for These Proteins and SGS3 in Arabidopsis. Noncoding RNA 2021, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Donaire, L.; Barajas, D.; Martínez-García, B.; Martínez-Priego, L.; Pagán, I.; Llave, C. Structural and genetic requirements for the biogenesis of tobacco rattle virus-derived small interfering RNAs. J. Virol. 2008, 82, 5167–5177. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Fan, B.; MacFarlane, S.A.; Chen, Z. Analysis of the involvement of an inducible Arabidopsis RNA-dependent RNA polymerase in antiviral defense. Mol. Plant Microbe Interact. 2003, 16, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Rakhshandehroo, F.; Takeshita, M.; Squires, J.; Palukaitis, P. The influence of RNA-dependent RNA polymerase 1 on potato virus Y infection and on other antiviral response genes. Mol. Plant Microbe Interact. 2009, 22, 1312–1318. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.W.; Liu, Y.R.; Liang, J.Y.; Wang, W.P.; Zhou, J.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q.; Shi, K. The relationship between the plant-encoded RNA-dependent RNA polymerase 1 and alternative oxidase in tomato basal defense against Tobacco mosaic virus. Planta 2015, 241, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Leibman, D.; Pashkovsky, E.; Shnaider, Y.; Shtarkman, M.; Gaba, V.; Gal-On, A. Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) gene family in melon. Plants 2022, 11, 1795. [Google Scholar] [CrossRef]
- Xie, Z.; Fan, B.; Chen, C.; Chen, Z. An important role of an inducible RNA-dependent RNA polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.J.; Carter, S.A.; Cole, A.B.; Cheng, N.H.; Nelson, R.S. A natural variant of a host RNA-dependent RNA polymerase is associated with increased susceptibility to viruses by Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2004, 101, 6297–6302. [Google Scholar] [CrossRef] [Green Version]
- Mourrain, P.; Béclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Gómez, G.; Martínez, G.; Pallás, V. Viroid-induced symptoms in Nicotiana benthamiana plants are dependent on RDR6 activity. Plant Physiol. 2008, 148, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Di Serio, F.; Martínez de Alba, A.E.; Navarro, B.; Gisel, A.; Flores, R. RNA-dependent RNA polymerase 6 delays accumulation and precludes meristem invasion of a viroid that replicates in the nucleus. J. Virol. 2010, 84, 2477–2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoi, T.; Kitabayashi, S.; Kasai, A.; Sugawara, K.; Adkar-Purushothama, C.R.; Senda, M.; Hataya, T.; Sano, T. Suppression of RNA-dependent RNA polymerase 6 in tomatoes allows potato spindle tuber viroid to invade basal part but not apical part including pluripotent stem cells of shoot apical meristem. PLoS ONE 2020, 15, e0236481. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Hou, G.; Sato, S.; Clemente, T.E.; Morris, T.J. RDR6 has a broad-spectrum but temperature-dependent antiviral defense role in Nicotiana benthamiana. J. Virol. 2005, 79, 15209–15217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Muangsan, N.; Beclin, C.; Vaucheret, H.; Robertson, D. Geminivirus VIGS of endogenous genes requires SGS2/SDE1 and SGS3 and defines a new branch in the genetic pathway for silencing in plants. Plant J. 2004, 38, 1004–1014. [Google Scholar] [CrossRef]
- Li, F.; Wang, Y.; Zhou, X. SGS3 Cooperates with RDR6 in Triggering Geminivirus-Induced Gene Silencing and in Suppressing Geminivirus Infection in Nicotiana benthamiana. Viruses 2017, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Mishra, S.K.; Rahman, J.; Taneja, J.; Sundaresan, G.; Mishra, N.S.; Mukherjee, S.K. Mungbean yellow mosaic Indian virus encoded AC2 protein suppresses RNA silencing by inhibiting Arabidopsis RDR6 and AGO1 activities. Virology 2015, 486, 158–172. [Google Scholar] [CrossRef] [Green Version]
- Szittya, G.; Silhavy, D.; Molnár, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Bánfalvi, Z.; Burgyán, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. Embo J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.; Sharma, N.; Hari-Gowthem, G.; Muthamilarasan, M.; Prasad, M. Tomato yellow leaf curl virus: Impact, challenges, and management. Trends Plant Sci. 2020, 25, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Monci, F.; Sánchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Morilla, G.; Krenz, B.; Jeske, H.; Bejarano, E.R.; Wege, C. Tête à tête of tomato yellow leaf curl virus and tomato yellow leaf curl sardinia virus in single nuclei. J. Virol. 2004, 78, 10715–10723. [Google Scholar] [CrossRef] [Green Version]
- Davino, S.; Napoli, C.; Dellacroce, C.; Miozzi, L.; Noris, E.; Davino, M.; Accotto, G.P. Two new natural begomovirus recombinants associated with the tomato yellow leaf curl disease co-exist with parental viruses in tomato epidemics in Italy. Virus Res. 2009, 143, 15–23. [Google Scholar] [CrossRef]
- Davino, S.; Miozzi, L.; Panno, S.; Rubio, L.; Davino, M.; Accotto, G.P. Recombination profiles between Tomato yellow leaf curl virus and Tomato yellow leaf curl Sardinia virus in laboratory and field conditions: Evolutionary and taxonomic implications. J. Gen. Virol. 2012, 93, 2712–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jammes, M.; Urbino, C.; Diouf, M.B.; Peterschmitt, M. Refining the emergence scenario of the invasive recombinant Tomato yellow leaf curl virus-IS76. Virology 2023, 578, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Wege, C.; Gotthardt, R.D.; Frischmuth, T.; Jeske, H. Fulfilling Koch’s postulates for Abutilon mosaic virus. Arch. Virol. 2000, 145, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Strohmeier, S.; Krenz, B.; Jeske, H. Evolutionary liberties of the Abutilon mosaic virus cluster. Virus Genes 2015, 50, 63–70. [Google Scholar] [CrossRef]
- Nelson, S.C. Abutilon Mosaic. Cooperative Extension Service/College of Tropical Agriculture and Human Resources (CTAHR), University of Hawai‘i at Mānoa, Honolulu, Hawai‘i, in Cooperation with the U.S. Department of Agriculture Plant Dis. Available online: http://www.ctahr.hawaii.edu/freepubs (accessed on 26 February 2023).
- Krenz, B.; Wege, C.; Jeske, H. Cell-free construction of disarmed Abutilon mosaic virus-based gene silencing vectors. J. Virol. Methods 2010, 169, 129–137. [Google Scholar] [CrossRef]
- Wege, C.; Siegmund, D. Synergism of a DNA and an RNA virus: Enhanced tissue infiltration of the begomovirus Abutilon mosaic virus (AbMV) mediated by Cucumber mosaic virus (CMV). Virology 2007, 357, 10–28. [Google Scholar] [CrossRef] [Green Version]
- Sardo, L.; Wege, C.; Kober, S.; Kocher, C.; Accotto, G.P.; Noris, E. RNA viruses and their silencing suppressors boost Abutilon mosaic virus, but not the Old World Tomato yellow leaf curl Sardinia virus. Virus Res. 2011, 161, 170–180. [Google Scholar] [CrossRef]
- Morra, M.R.; Petty, I.T. Tissue specificity of geminivirus infection is genetically determined. Plant Cell 2000, 12, 2259–2270. [Google Scholar] [CrossRef] [Green Version]
- Jeske, H. Geminiviruses. Curr. Top. Microbiol. Immunol. 2009, 331, 185–226. [Google Scholar] [CrossRef]
- Wege, C.; Pohl, D. Abutilon mosaic virus DNA B component supports mechanical virus transmission, but does not counteract begomoviral phloem limitation in transgenic plants. Virology 2007, 365, 173–186. [Google Scholar] [CrossRef] [Green Version]
- Frischmuth, T.; Roberts, S.; von Arnim, A.; Stanley, J. Specificity of bipartite geminivirus movement proteins. Virology 1993, 196, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Kheyr-Pour, A.; Bendahmane, M.; Matzeit, V.; Accotto, G.P.; Crespi, S.; Gronenborn, B. Tomato yellow leaf curl virus from Sardinia is a whitefly-transmitted monopartite geminivirus. Nucleic Acids Res. 1991, 19, 6763–6769. [Google Scholar] [CrossRef]
- Ghanim, M.; Brumin, M.; Popovski, S. A simple, rapid and inexpensive method for localization of Tomato yellow leaf curl virus and Potato leafroll virus in plant and insect vectors. J. Virol. Methods 2009, 159, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Blevins, T. Northern blotting techniques for small RNAs. Methods Mol. Biol. 2010, 631, 87–107. [Google Scholar] [CrossRef]
- Pall, G.S.; Hamilton, A.J. Improved northern blot method for enhanced detection of small RNA. Nat. Protoc. 2008, 3, 1077–1084. [Google Scholar] [CrossRef]
- Goto, K.; Kobori, T.; Kosaka, Y.; Natsuaki, T.; Masuta, C. Characterization of silencing suppressor 2b of cucumber mosaic virus based on examination of its small RNA-binding abilities. Plant Cell Physiol. 2007, 48, 1050–1060. [Google Scholar] [CrossRef] [Green Version]
- Ludman, M.; Fátyol, K. The virological model plant, Nicotiana benthamiana expresses a single functional RDR6 homeolog. Virology 2019, 537, 143–148. [Google Scholar] [CrossRef]
- Matsuo, K.; Atsumi, G. CRISPR/Cas9-mediated knockout of the RDR6 gene in Nicotiana benthamiana for efficient transient expression of recombinant proteins. Planta 2019, 250, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Weigel, K.; Pohl, J.O.; Wege, C.; Jeske, H. A population genetics perspective on geminivirus infection. J. Virol. 2015, 89, 11926–11934. [Google Scholar] [CrossRef] [Green Version]
- Horns, T.; Jeske, H. Localization of abutilon mosaic virus (AbMV) DNA within leaf tissue by in situ hybridization. Virology 1991, 181, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Wege, C.; Saunders, K.; Stanley, J.; Jeske, H. Comparative analysis of tissue tropism of bipartite geminiviruses. J. Phytopathol. 2001, 149, 359–368. [Google Scholar] [CrossRef]
- Adkar-Purushothama, C.R.; Perreault, J.P. Suppression of RNA-Dependent RNA polymerase 6 favors the accumulation of potato spindle tuber viroid in Nicotiana benthamiana. Viruses 2019, 11, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479-480, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.Y.; Park, J.M. Cross-Talk in Viral Defense Signaling in Plants. Front. Microbiol. 2016, 7, 2068. [Google Scholar] [CrossRef] [Green Version]
- Pontier, D.; Picart, C.; Roudier, F.; Garcia, D.; Lahmy, S.; Azevedo, J.; Alart, E.; Laudié, M.; Karlowski, W.M.; Cooke, R.; et al. NERD, a plant-specific GW protein, defines an additional RNAi-dependent chromatin-based pathway in Arabidopsis. Mol. Cell 2012, 48, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuthikattu, S.; McCue, A.D.; Panda, K.; Fultz, D.; DeFraia, C.; Thomas, E.N.; Slotkin, R.K. The initiation of epigenetic silencing of active transposable elements is triggered by RDR6 and 21-22 nucleotide small interfering RNAs. Plant Physiol. 2013, 162, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Panda, K.; Slotkin, R.K. Proposed mechanism for the initiation of transposable element silencing by the RDR6-directed DNA methylation pathway. Plant Signal Behav. 2013, 8, e25206. [Google Scholar] [CrossRef] [Green Version]
- Loriato, V.A.P.; Martins, L.G.C.; Euclydes, N.C.; Reis, P.A.B.; Duarte, C.E.M.; Fontes, E.P.B. Engineering resistance against geminiviruses: A review of suppressed natural defenses and the use of RNAi and the CRISPR/Cas system. Plant Sci. 2020, 292, 110410. [Google Scholar] [CrossRef]
- Rodríguez-Negrete, E.; Lozano-Durán, R.; Piedra-Aguilera, A.; Cruzado, L.; Bejarano, E.R.; Castillo, A.G. Geminivirus Rep protein interferes with the plant DNA methylation machinery and suppresses transcriptional gene silencing. New Phytol. 2013, 199, 464–475. [Google Scholar] [CrossRef] [Green Version]
- Glick, E.; Zrachya, A.; Levy, Y.; Mett, A.; Gidoni, D.; Belausov, E.; Citovsky, V.; Gafni, Y. Interaction with host SGS3 is required for suppression of RNA silencing by tomato yellow leaf curl virus V2 protein. Proc. Natl. Acad. Sci. USA 2008, 105, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Yang, X.; Wang, Y.; Xie, Y.; Zhou, X. Tomato Yellow Leaf Curl Virus V2 Interacts with Host Histone Deacetylase 6 To Suppress Methylation-Mediated Transcriptional Gene Silencing in Plants. J. Virol. 2018, 92, e00036-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Ding, Y.; He, L.; Zhang, G.; Zhu, J.K.; Lozano-Duran, R. A virus-encoded protein suppresses methylation of the viral genome through its interaction with AGO4 in the Cajal body. eLife 2020, 9, e55542. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA silencing against geminivirus: Complementary action of posttranscriptional gene silencing and transcriptional gene silencing in host recovery. J. Virol. 2009, 83, 1332–1340. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Inoculum | Days Post-Inoculation (dpi) | ||
|---|---|---|---|
| 19 | 31 | 42 | |
| AbMV | 0% | 0–25% | 50–75% |
| TYLCSV | 0% | 0% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noris, E.; Pegoraro, M.; Palzhoff, S.; Urrejola, C.; Wochner, N.; Kober, S.; Ruoff, K.; Matić, S.; Schnepf, V.; Weisshaar, N.; et al. Differential Effects of RNA-Dependent RNA Polymerase 6 (RDR6) Silencing on New and Old World Begomoviruses in Nicotiana benthamiana. Viruses 2023, 15, 919. https://doi.org/10.3390/v15040919
Noris E, Pegoraro M, Palzhoff S, Urrejola C, Wochner N, Kober S, Ruoff K, Matić S, Schnepf V, Weisshaar N, et al. Differential Effects of RNA-Dependent RNA Polymerase 6 (RDR6) Silencing on New and Old World Begomoviruses in Nicotiana benthamiana. Viruses. 2023; 15(4):919. https://doi.org/10.3390/v15040919
Chicago/Turabian StyleNoris, Emanuela, Mattia Pegoraro, Sandra Palzhoff, Catalina Urrejola, Nicolai Wochner, Sigi Kober, Kerstin Ruoff, Slavica Matić, Vera Schnepf, Nina Weisshaar, and et al. 2023. "Differential Effects of RNA-Dependent RNA Polymerase 6 (RDR6) Silencing on New and Old World Begomoviruses in Nicotiana benthamiana" Viruses 15, no. 4: 919. https://doi.org/10.3390/v15040919