
Characterization of Anti-Poliovirus Compounds Isolated from Edible Plants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Screening of Edible Plant Extracts for Anti-PV Activity
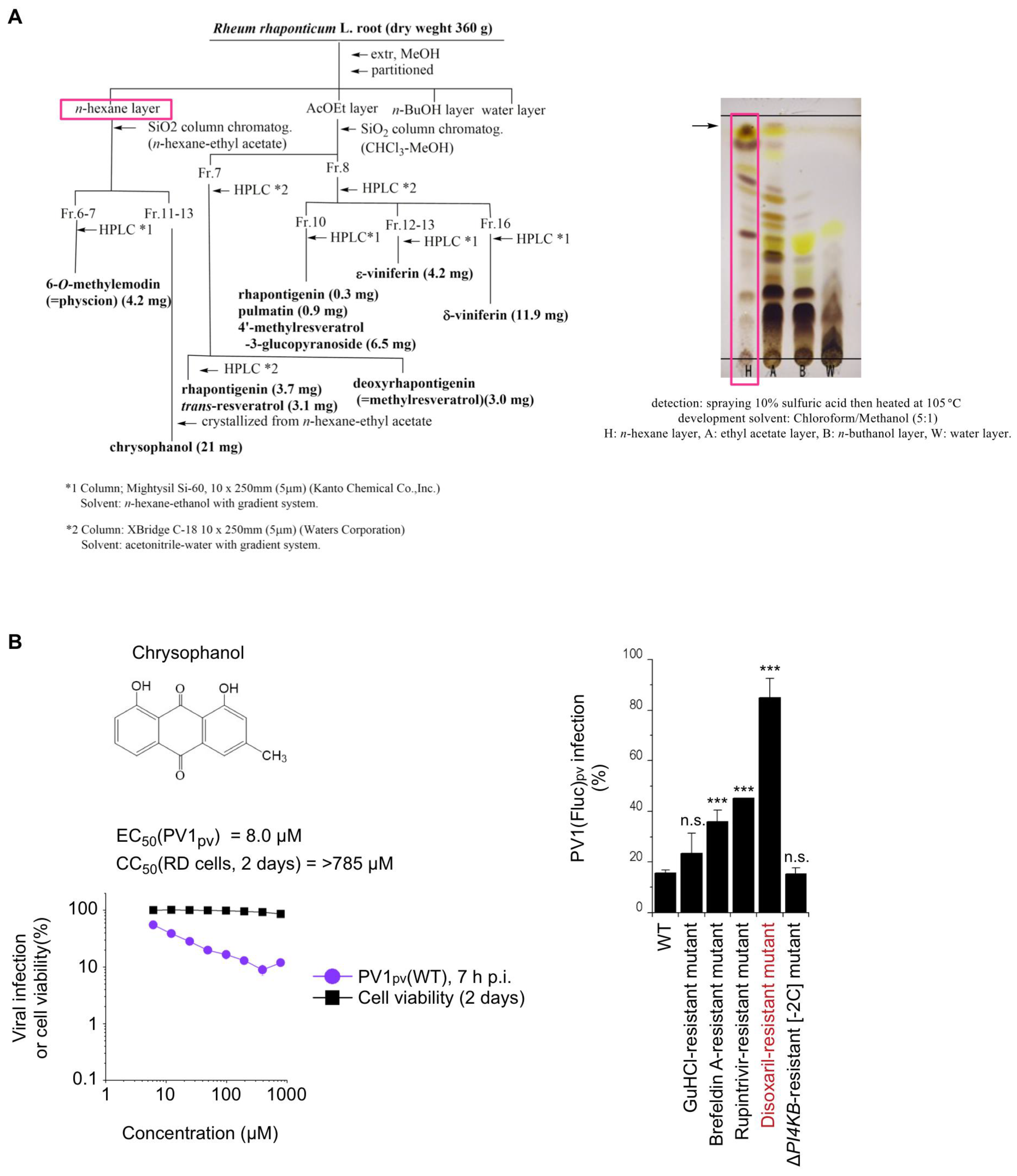
3.2. Purification and Structure Determination of Anti-PV Compound in R. Rhaponticum
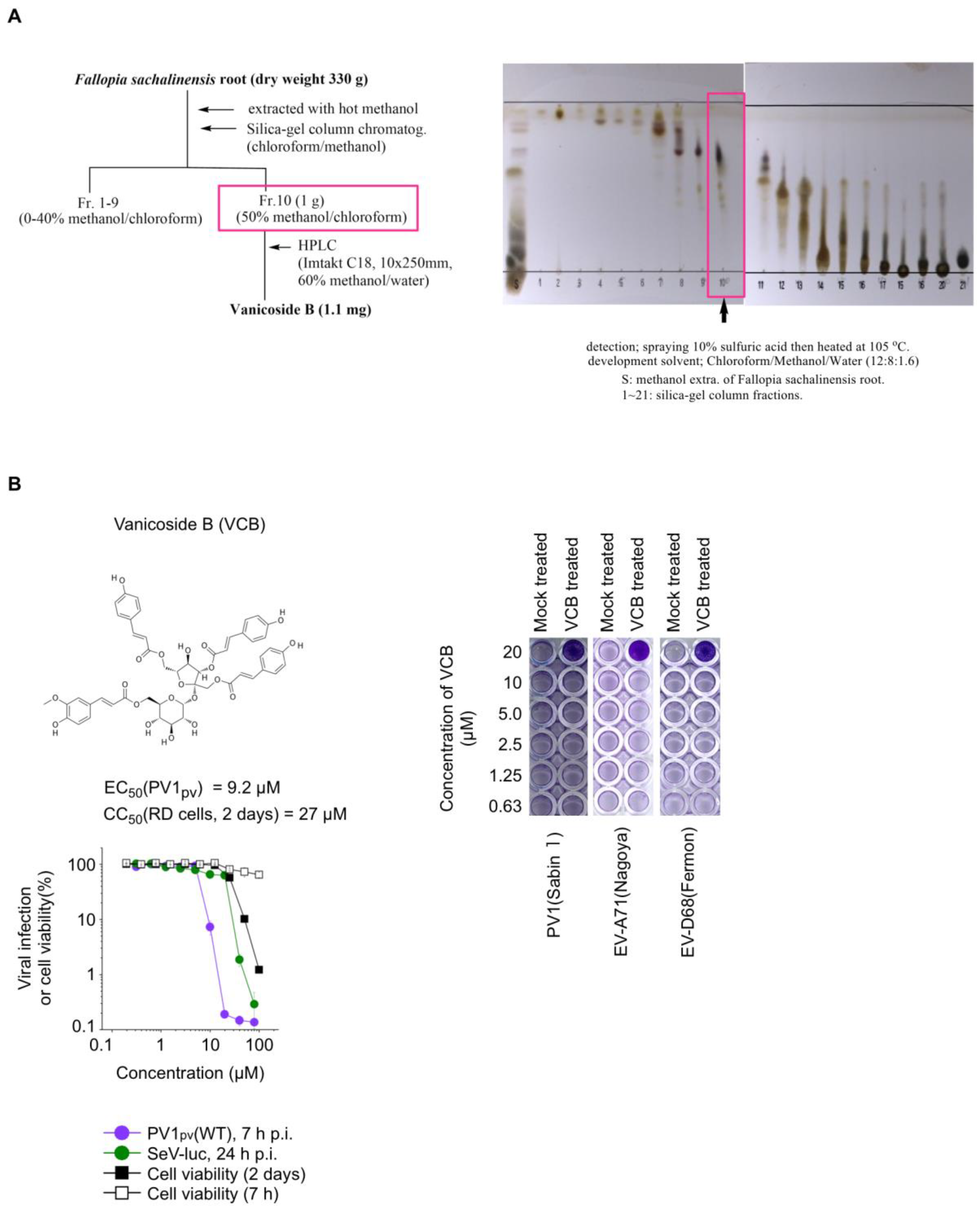
3.3. Purification and Structure Determination of Anti-PV Compound in F. Sachalinensis
3.4. Characterization of Anti-PV Activity of VCB
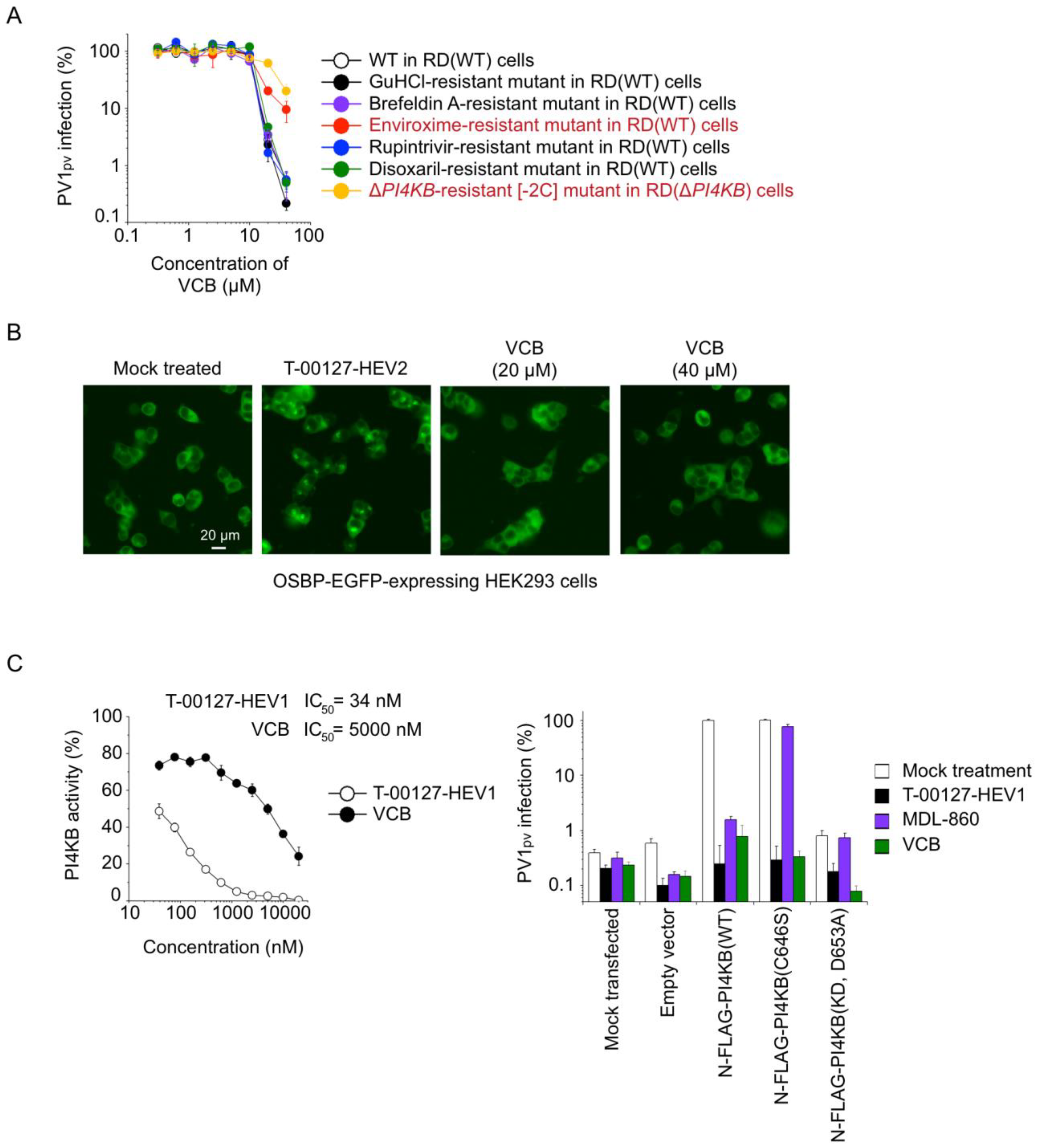
3.5. Mechanism of Anti-PV Activity of VCB
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| CCID50 | 50% cell culture infectious dose |
| cVDPV | circulating vaccine-derived poliovirus |
| EC50 | 50% effective concentration |
| EV | enterovirus |
| GuHCl | guanidine hydrochloride |
| IC50 | 50% inhibitory concentration |
| IPV | inactivated PV vaccine |
| MOI | multiplicity of infection |
| OPV | oral PV vaccine |
| OSBP | oxysterol-binding protein |
| P.i. | post-infection |
| PI4P | phosphatidylinositol 4-monophosphate |
| PV | poliovirus |
| PV1pv | type 1 PV pseudovirus |
| RO | replication organelle |
| SeV-luc | luciferase-encoding Sendai virus |
| SI | selectivity index |
| VCB | vanicoside B |
References
- Kitamura, N.; Semler, B.L.; Rothberg, P.G.; Larsen, G.R.; Adler, C.J.; Dorner, A.J.; Emini, E.A.; Hanecak, R.; Lee, J.J.; van der Werf, S.; et al. Primary structure, gene organization and polypeptide expression of poliovirus RNA. Nature 1981, 291, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Macklin, G.R.; O’Reilly, K.M.; Grassly, N.C.; Edmunds, W.J.; Mach, O.; Santhana Gopala Krishnan, R.; Voorman, A.; Vertefeuille, J.F.; Abdelwahab, J.; Gumede, N.; et al. Evolving epidemiology of poliovirus serotype 2 following withdrawal of the serotype 2 oral poliovirus vaccine. Science 2020, 368, 401–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Available online: https://polioeradication.org/polio-today/polio-now/this-week/circulating-vaccine-derived-poliovirus/ (accessed on 27 February 2023).
- Link-Gelles, R.; Lutterloh, E.; Schnabel Ruppert, P.; Backenson, P.B.; St George, K.; Rosenberg, E.S.; Anderson, B.J.; Fuschino, M.; Popowich, M.; Punjabi, C.; et al. Public Health Response to a Case of Paralytic Poliomyelitis in an Unvaccinated Person and Detection of Poliovirus in Wastewater—New York, June-August 2022. MMWR Morb. Mortal Wkly. Rep. 2022, 71, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Klapsa, D.; Wilton, T.; Zealand, A.; Bujaki, E.; Saxentoff, E.; Troman, C.; Shaw, A.G.; Tedcastle, A.; Majumdar, M.; Mate, R.; et al. Sustained detection of type 2 poliovirus in London sewage between February and July, 2022, by enhanced environmental surveillance. Lancet 2022, 400, 1531–1538. [Google Scholar] [CrossRef]
- Anis, E.; Kopel, E.; Singer, S.R.; Kaliner, E.; Moerman, L.; Moran-Gilad, J.; Sofer, D.; Manor, Y.; Shulman, L.M.; Mendelson, E.; et al. Insidious reintroduction of wild poliovirus into Israel, 2013. Eurosurveillance 2013, 18, 20586. [Google Scholar] [CrossRef] [Green Version]
- Yeh, M.T.; Bujaki, E.; Dolan, P.T.; Smith, M.; Wahid, R.; Konz, J.; Weiner, A.J.; Bandyopadhyay, A.S.; Van Damme, P.; De Coster, I.; et al. Engineering the Live-Attenuated Polio Vaccine to Prevent Reversion to Virulence. Cell Host Microbe 2020, 27, 736–751.e8. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.S.; Zipursky, S. A novel tool to eradicate an ancient scourge: The novel oral polio vaccine type 2 story. Lancet Infect. Dis. 2023, 23, e67–e71. [Google Scholar] [CrossRef]
- Collett, M.S.; Neyts, J.; Modlin, J.F. A case for developing antiviral drugs against polio. Antiviral Res. 2008, 79, 179–187. [Google Scholar] [CrossRef]
- Committee on Development of a Polio Antiviral and Its Potential Role in Global Poliomyelitis Eradication, National Research Council. Exploring the Role of Antiviral Drugs in the Eradication of Polio: Workshop Report; The National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- De Palma, A.M.; Vliegen, I.; De Clercq, E.; Neyts, J. Selective inhibitors of picornavirus replication. Med. Res. Rev. 2008, 28, 823–884. [Google Scholar] [CrossRef]
- Maynell, L.A.; Kirkegaard, K.; Klymkowsky, M.W. Inhibition of poliovirus RNA synthesis by brefeldin A. J. Virol. 1992, 66, 1985–1994. [Google Scholar] [CrossRef] [Green Version]
- Irurzun, A.; Perez, L.; Carrasco, L. Involvement of membrane traffic in the replication of poliovirus genomes: Effects of brefeldin A. Virology 1992, 191, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Bordeleau, M.E.; Mori, A.; Oberer, M.; Lindqvist, L.; Chard, L.S.; Higa, T.; Belsham, G.J.; Wagner, G.; Tanaka, J.; Pelletier, J. Functional characterization of IRESes by an inhibitor of the RNA helicase eIF4A. Nat. Chem. Biol. 2006, 2, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Geller, R.; Vignuzzi, M.; Andino, R.; Frydman, J. Evolutionary constraints on chaperone-mediated folding provide an antiviral approach refractory to development of drug resistance. Genes Dev. 2007, 21, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, H.H.; Kunz, A.; Simon, V.A.; Palese, P.; Shaw, M.L. Broad-spectrum antiviral that interferes with de novo pyrimidine biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, 5777–5782. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, D.E.; Jang, K.S.; Kim, S.J.; Cho, S.; Kim, C. Gemcitabine, a broad-spectrum antiviral drug, suppresses enterovirus infections through innate immunity induced by the inhibition of pyrimidine biosynthesis and nucleotide depletion. Oncotarget 2017, 8, 115315–115325. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, E.; Hu, C.; Cheng, H.; Chen, C.Y.; Huang, D.; Wang, R.; Zhao, Y.; Rong, L.; Vignuzzi, M.; et al. Cell-Based High-Throughput Screening Assay Identifies 2′,2′-Difluoro-2′-deoxycytidine Gemcitabine as a Potential Antipoliovirus Agent. ACS Infect. Dis. 2017, 3, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Grollman, A.P. Inhibitors of protein biosynthesis. V. Effects of emetine on protein and nucleic acid biosynthesis in HeLa cells. J. Biol. Chem. 1968, 243, 4089–4094. [Google Scholar] [CrossRef] [PubMed]
- Wikel, J.H.; Paget, C.J.; DeLong, D.C.; Nelson, J.D.; Wu, C.Y.; Paschal, J.W.; Dinner, A.; Templeton, R.J.; Chaney, M.O.; Jones, N.D.; et al. Synthesis of syn and anti isomers of 6-[[(hydroxyimino)phenyl]methyl]-1-[(1-methylethyl)sulfonyl]-1H-benzimidaz ol-2-amine. Inhibitors of rhinovirus multiplication. J. Med. Chem. 1980, 23, 368–372. [Google Scholar] [CrossRef]
- Heinz, B.A.; Vance, L.M. The antiviral compound enviroxime targets the 3A coding region of rhinovirus and poliovirus. J. Virol. 1995, 69, 4189–4197. [Google Scholar] [CrossRef] [Green Version]
- Hsu, N.Y.; Ilnytska, O.; Belov, G.; Santiana, M.; Chen, Y.H.; Takvorian, P.M.; Pau, C.; van der Schaar, H.; Kaushik-Basu, N.; Balla, T.; et al. Viral reorganization of the secretory pathway generates distinct organelles for RNA replication. Cell 2010, 141, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Kojima, H.; Nagano, T.; Okabe, T.; Wakita, T.; Shimizu, H. Phosphatidylinositol 4-kinase III beta is a target of enviroxime-like compounds for antipoliovirus activity. J. Virol. 2011, 85, 2364–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delang, L.; Paeshuyse, J.; Neyts, J. The role of phosphatidylinositol 4-kinases and phosphatidylinositol 4-phosphate during viral replication. Biochem. Pharmacol. 2012, 84, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.M.; Mitchell, D.R.; Palmer, N.J.; Van de Poel, H.; Conrath, K.; Andrews, M.; Leyssen, P.; Neyts, J. Identification of a series of compounds with potent antiviral activity for the treatment of enterovirus infections. ACS Med. Chem. Lett. 2013, 4, 585–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejdrova, I.; Chalupska, D.; Kogler, M.; Sala, M.; Plackova, P.; Baumlova, A.; Hrebabecky, H.; Prochazkova, E.; Dejmek, M.; Guillon, R.; et al. Highly Selective Phosphatidylinositol 4-Kinase IIIbeta Inhibitors and Structural Insight into Their Mode of Action. J. Med. Chem. 2015, 58, 3767–3793. [Google Scholar] [CrossRef]
- Arita, M.; Dobrikov, G.; Purstinger, G.; Galabov, A.S. Allosteric Regulation of Phosphatidylinositol 4-Kinase III Beta by an Antipicornavirus Compound MDL-860. ACS Infect. Dis. 2017, 3, 585–594. [Google Scholar] [CrossRef]
- De Palma, A.M.; Thibaut, H.J.; van der Linden, L.; Lanke, K.; Heggermont, W.; Ireland, S.; Andrews, R.; Arimilli, M.; Altel, T.; De Clercq, E.; et al. Mutations in the non-structural protein 3A confer resistance to the novel enterovirus replication inhibitor TTP-8307. Antimicrob. Agents Chemother. 2009, 53, 1850–1857. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Takebe, Y.; Wakita, T.; Shimizu, H. A bifunctional anti-enterovirus compound that inhibits replication and early stage of enterovirus 71 infection. J. Gen. Virol. 2010, 91, 2734–2744. [Google Scholar] [CrossRef]
- Arita, M.; Kojima, H.; Nagano, T.; Okabe, T.; Wakita, T.; Shimizu, H. Oxysterol-binding protein family I is the target of minor enviroxime-like compounds. J. Virol. 2013, 87, 4252–4260. [Google Scholar] [CrossRef] [Green Version]
- Strating, J.R.; van der Linden, L.; Albulescu, L.; Bigay, J.; Arita, M.; Delang, L.; Leyssen, P.; van der Schaar, H.M.; Lanke, K.H.; Thibaut, H.J.; et al. Itraconazole Inhibits Enterovirus Replication by Targeting the Oxysterol-Binding Protein. Cell Rep. 2015, 10, 600–615. [Google Scholar] [CrossRef] [Green Version]
- Albulescu, L.; Bigay, J.; Biswas, B.; Weber-Boyvat, M.; Dorobantu, C.M.; Delang, L.; van der Schaar, H.M.; Jung, Y.S.; Neyts, J.; Olkkonen, V.M.; et al. Uncovering oxysterol-binding protein (OSBP) as a target of the anti-enteroviral compound TTP-8307. Antiviral Res. 2017, 140, 37–44. [Google Scholar] [CrossRef]
- Ho, M.; Chen, E.R.; Hsu, K.H.; Twu, S.J.; Chen, K.T.; Tsai, S.F.; Wang, J.R.; Shih, S.R. An epidemic of enterovirus 71 infection in Taiwan. N. Engl. J. Med. 1999, 341, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Mirand, A.; Josset, L.; Henquell, C.; Hecquet, D.; Pilorge, L.; Petitjean-Lecherbonnier, J.; Manoha, C.; Legoff, J.; Deback, C.; et al. Epidemiological and clinical characteristics of patients infected with enterovirus D68, France, July to December 2014. Eurosurveillance 2016, 21, 30226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arita, M.; Fuchino, H.; Kawakami, H.; Ezaki, M.; Kawahara, N. Characterization of a New Antienterovirus D68 Compound Purified from Avocado. ACS Infect. Dis. 2020, 6, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Arita, M.; Bigay, J. Poliovirus Evolution toward Independence from the Phosphatidylinositol-4 Kinase III beta/Oxysterol-Binding Protein Family I Pathway. ACS Infect. Dis. 2019, 5, 962–973. [Google Scholar] [CrossRef]
- Arita, M.; Nagata, N.; Sata, T.; Miyamura, T.; Shimizu, H. Quantitative analysis of poliomyelitis-like paralysis in mice induced by a poliovirus replicon. J. Gen. Virol. 2006, 87 Pt 11, 3317–3327. [Google Scholar] [CrossRef]
- Baltera, R.F., Jr.; Tershak, D.R. Guanidine-resistant mutants of poliovirus have distinct mutations in peptide 2C. J. Virol. 1989, 63, 4441–4444. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S.; Saleh, M.C.; Gitlin, L.; Beske, O.; Andino, R. The poliovirus replication machinery can escape inhibition by an antiviral drug that targets a host cell protein. J. Virol. 2004, 78, 3378–3386. [Google Scholar] [CrossRef] [Green Version]
- Tanner, E.J.; Liu, H.M.; Oberste, M.S.; Pallansch, M.; Collett, M.S.; Kirkegaard, K. Dominant drug targets suppress the emergence of antiviral resistance. eLife 2014, 3, e03830. [Google Scholar] [CrossRef]
- Mosser, A.G.; Sgro, J.Y.; Rueckert, R.R. Distribution of drug resistance mutations in type 3 poliovirus identifies three regions involved in uncoating functions. J. Virol. 1994, 68, 8193–8201. [Google Scholar] [CrossRef] [Green Version]
- Purstinger, G.; De Palma, A.M.; Zimmerhofer, G.; Huber, S.; Ladurner, S.; Neyts, J. Synthesis and anti-CVB 3 evaluation of substituted 5-nitro-2-phenoxybenzonitriles. Bioorg. Med. Chem. Lett. 2008, 18, 5123–5125. [Google Scholar] [CrossRef]
- Arita, M.; Wakita, T.; Shimizu, H. Characterization of pharmacologically active compounds that inhibit poliovirus and enterovirus 71 infectivity. J. Gen. Virol. 2008, 89 Pt 10, 2518–2530. [Google Scholar] [CrossRef] [PubMed]
- Meselhy, M.R. Inhibition of LPS-induced NO production by the oleogum resin of Commiphora wightii and its constituents. Phytochemistry 2003, 62, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.A.; Urban, S. Phytochemical investigation of the Australian lichens Ramalina glaucescens and Xanthoria parietina. Nat. Prod. Commun. 2009, 4, 959–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Lee, B.W.; Kim, M.R.; Jun, J.G. Syntheses of Resveratrol and its Hydroxylated Derivatives as Radical Scavenger and Tyrosinase Inhibitor. Bull. Korean Chem. Soc. 2010, 31, 971–975. [Google Scholar] [CrossRef] [Green Version]
- Commodari, F.; Khiat, A.; Ibrahimi, S.; Brizius, A.R.; Kalkstein, N. Comparison of the phytoestrogen trans-resveratrol (3,4′,5-trihydroxystilbene) structures from x-ray diffraction and solution NMR. Magn. Reson. Chem. 2005, 43, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Murai, Y.; Soediro, I.; Soetarno, S.; Sastrodihardjo, S. Cytotoxic anthraquinones from Rheum pulmatum. Phytochemistry 1992, 31, 1063–1065. [Google Scholar] [CrossRef]
- Dai, L.-M.; Tang, J.; Li, H.-L.; Shen, Y.-H.; Peng, C.-Y.; Zhang, W.-D. A new stilbene glycoside from the n-butanol fraction of Veratrum dahuricum. Chem. Nat. Compd. 2009, 45, 325–329. [Google Scholar] [CrossRef]
- Ngoc, T.M.; Hung, T.M.; Thuong, P.T.; Na, M.; Kim, H.; Ha, D.T.; Min, B.-S.; Minh, P.T.H.; Bae, K. Inhibition of Human Low Density Lipoprotein and High Density Lipoprotein Oxidation by Oligostilbenes from Rhubarb. Biol. Pharm. Bull. 2008, 31, 1809–1812. [Google Scholar] [CrossRef] [Green Version]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, K.; Viret, O. Delta-viniferin, a resveratrol dehydrodimer: One of the major stilbenes synthesized by stressed grapevine leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef]
- Semple, S.J.; Pyke, S.M.; Reynolds, G.D.; Flower, R.L. In vitro antiviral activity of the anthraquinone chrysophanic acid against poliovirus. Antiviral Res. 2001, 49, 169–178. [Google Scholar] [CrossRef]
- Kiem, P.V.; Nhiem, N.X.; Cuong, N.X.; Hoa, T.Q.; Huong, H.T.; Huong, L.M.; Minh, C.V.; Kim, Y.H. New phenylpropanoid esters of sucrose from Polygonum hydropiper and their antioxidant activity. Arch. Pharm. Res. 2008, 31, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, N.D.; Dawson, P.A.; Ho, Y.K.; Brown, M.S.; Goldstein, J.L. Translocation of oxysterol binding protein to Golgi apparatus triggered by ligand binding. J. Cell Biol. 1992, 116, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, J.; Arita, M.; Sakai, S.; Kojima, H.; Senda, M.; Senda, T.; Hanada, K.; Kato, R. Ligand Recognition by the Lipid Transfer Domain of Human OSBP Is Important for Enterovirus Replication. ACS Infect. Dis. 2022, 8, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Torney, H.L.; Dulworth, J.K.; Steward, D.L. Antiviral activity and mechanism of action of 2-(3,4-dichlorophenoxy)-5-nitrobenzonitrile (MDL-860). Antimicrob. Agents Chemother. 1982, 22, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Ishitsuka, H.; Ohsawa, C.; Ohiwa, T.; Umeda, I.; Suhara, Y. Antipicornavirus flavone Ro 09-0179. Antimicrob. Agents Chemother. 1982, 22, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Galabov, A.S.; Nikolaeva, L.; Philipov, S. Aporphinoid alkaloid glaucinone: A selective inhibitor of poliovirus replication. Antiviral. Res. 1995, 26 (Suppl. 1), A347. [Google Scholar] [CrossRef]
- Nikolaeva-Glomb, L.; Philipov, S.; Galabov, A.S. Oxoglaucin—Enterovirus replication inhibitor: Study on the antiviral spectrum, mode of action and development of resistance (in Bulgarian). J. Bulg. Acad. Sci. 2007, 120, 22–26. [Google Scholar]
- Wang, J.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Anti-enterovirus 71 effects of chrysin and its phosphate ester. PLoS ONE 2014, 9, e89668. [Google Scholar] [CrossRef] [Green Version]
- Gunaseelan, S.; Wong, K.Z.; Min, N.; Sun, J.; Ismail, N.; Tan, Y.J.; Lee, R.C.H.; Chu, J.J.H. Prunin suppresses viral IRES activity and is a potential candidate for treating enterovirus A71 infection. Sci. Transl. Med. 2019, 11, eaar5759. [Google Scholar] [CrossRef]
- Zhang, H.M.; Wang, F.; Qiu, Y.; Ye, X.; Hanson, P.; Shen, H.; Yang, D. Emodin inhibits coxsackievirus B3 replication via multiple signalling cascades leading to suppression of translation. Biochem. J. 2016, 473, 473–485. [Google Scholar] [CrossRef]
- Zhong, T.; Zhang, L.Y.; Wang, Z.Y.; Wang, Y.; Song, F.M.; Zhang, Y.H.; Yu, J.H. Rheum emodin inhibits enterovirus 71 viral replication and affects the host cell cycle environment. Acta Pharmacol. Sin. 2017, 38, 392–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugent, C.I.; Johnson, K.L.; Sarnow, P.; Kirkegaard, K. Functional coupling between replication and packaging of poliovirus replicon RNA. J. Virol. 1999, 73, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arita, M. High-Order Epistasis and Functional Coupling of Infection Steps Drive Virus Evolution toward Independence from a Host Pathway. Microbiol. Spectr. 2021, 9, e0080021. [Google Scholar] [CrossRef] [PubMed]
- Buontempo, P.J.; Cox, S.; Wright-Minogue, J.; DeMartino, J.L.; Skelton, A.M.; Ferrari, E.; Albin, R.; Rozhon, E.J.; Girijavallabhan, V.; Modlin, J.F.; et al. SCH 48973: A potent, broad-spectrum, antienterovirus compound. Antimicrob. Agents Chemother. 1997, 41, 1220–1225. [Google Scholar] [CrossRef] [Green Version]
- Oberste, M.S.; Moore, D.; Anderson, B.; Pallansch, M.A.; Pevear, D.C.; Collett, M.S. In vitro antiviral activity of V-073 against polioviruses. Antimicrob. Agents Chemother. 2009, 53, 4501–4503. [Google Scholar] [CrossRef] [Green Version]
- Patick, A.K.; Brothers, M.A.; Maldonado, F.; Binford, S.; Maldonado, O.; Fuhrman, S.; Petersen, A.; Smith, G.J., 3rd; Zalman, L.S.; Burns-Naas, L.A.; et al. In vitro antiviral activity and single-dose pharmacokinetics in humans of a novel, orally bioavailable inhibitor of human rhinovirus 3C protease. Antimicrob. Agents Chemother. 2005, 49, 2267–2275. [Google Scholar] [CrossRef] [Green Version]
- Rhoden, E.; Liu, H.M.; Wang-Chern, S.W.; Oberste, M.S. Anti-poliovirus activity of protease inhibitor AG-7404, and assessment of in vitro activity in combination with antiviral capsid inhibitor compounds. Antiviral Res. 2013, 98, 186–191. [Google Scholar] [CrossRef]
- Kankam, M.K.; Burns, J.M.; Collett, M.S.; Corrado, M.L.; Hincks, J.R. A Phase 1 Study of the Safety, Tolerability, and Pharmacokinetics of Single and Multiple Oral Doses of V-7404 in Healthy Adult Volunteers. Antimicrob. Agents Chemother. 2021, 65, e0102921. [Google Scholar] [CrossRef]
- McKinlay, M.A.; Collett, M.S.; Hincks, J.R.; Oberste, M.S.; Pallansch, M.A.; Okayasu, H.; Sutter, R.W.; Modlin, J.F.; Dowdle, W.R. Progress in the development of poliovirus antiviral agents and their essential role in reducing risks that threaten eradication. J. Infect. Dis. 2014, 210 (Suppl. 1), S447–S453. [Google Scholar] [CrossRef] [Green Version]
- WHO. Available online: https://polioeradication.org/tools-and-library/current-research-areas/antivirals/ (accessed on 27 February 2023).
- Zimmermann, M.L.; Sneden, A.T. Vanicosides A and B, protein kinase C inhibitors from Polygonum pensylvanicum. J. Nat. Prod. 1994, 57, 236–242. [Google Scholar] [CrossRef]
- Ahmad, R.; Sahidin, I.; Taher, M.; Low, C.; Noor, N.M.; Sillapachaiyaporn, C.; Chuchawankul, S.; Sarachana, T.; Tencomnao, T.; Iskandar, F.; et al. Polygonumins A, a newly isolated compound from the stem of Polygonum minus Huds with potential medicinal activities. Sci. Rep. 2018, 8, 4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrot-Hadzik, I.; Zmudzinski, M.; Matkowski, A.; Preissner, R.; Kesik-Brodacka, M.; Hadzik, J.; Drag, M.; Abel, R. Reynoutria Rhizomes as a Natural Source of SARS-CoV-2 Mpro Inhibitors-Molecular Docking and In Vitro Study. Pharmaceuticals 2021, 14, 742. [Google Scholar] [CrossRef] [PubMed]
- Arita, M. Phosphatidylinositol-4 kinase III beta and oxysterol-binding protein accumulate unesterified cholesterol on poliovirus-induced membrane structure. Microbiol. Immunol. 2014, 58, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Arita, M. Mechanism of Poliovirus Resistance to Host Phosphatidylinositol-4 Kinase III β Inhibitor. ACS Infect. Dis. 2016, 2, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, H.; Dorobantu, C.M.; van der Schaar, H.M.; van Kuppeveld, F.J.M. Modulation of proteolytic polyprotein processing by coxsackievirus mutants resistant to inhibitors targeting phosphatidylinositol-4-kinase IIIbeta or oxysterol binding protein. Antiviral Res. 2017, 147, 86–90. [Google Scholar] [CrossRef]
- Melia, C.E.; van der Schaar, H.M.; Lyoo, H.; Limpens, R.; Feng, Q.; Wahedi, M.; Overheul, G.J.; van Rij, R.P.; Snijder, E.J.; Koster, A.J.; et al. Escaping Host Factor PI4KB Inhibition: Enterovirus Genomic RNA Replication in the Absence of Replication Organelles. Cell Rep. 2017, 21, 587–599. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, M.J.; Borawski, J.; Bose, A.; Capacci-Daniel, C.; Colvin, R.; Dennehy, M.; Ding, J.; Dobler, M.; Drumm, J.; Gaither, L.A.; et al. Anti-hepatitis C virus activity and toxicity of type III phosphatidylinositol-4-kinase beta inhibitors. Antimicrob. Agents Chemother. 2012, 56, 5149–5156. [Google Scholar] [CrossRef] [Green Version]
- Spickler, C.; Lippens, J.; Laberge, M.K.; Desmeules, S.; Bellavance, E.; Garneau, M.; Guo, T.; Hucke, O.; Leyssen, P.; Neyts, J.; et al. Phosphatidylinositol 4-Kinase III Beta Is Essential for Replication of Human Rhinovirus and Its Inhibition Causes a Lethal Phenotype In Vivo. Antimicrob. Agents Chemother. 2013, 57, 3358–3368. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Fujita, M.; Ishibashi, Y.; Nomanbhoy, T.; Rosenblum, J.S.; Nagasawa, M. 134. KRP-A218, an Orally Active and Selective PI4KB Inhibitor with Broad-Spectrum Anti-Rhinovirus Activity, Has Potent Therapeutic Antiviral Activity In Vivo. Open Forum Infect. Dis. 2021, 8 (Suppl. 1), S82. [Google Scholar] [CrossRef]
- Knight, Z.A.; Gonzalez, B.; Feldman, M.E.; Zunder, E.R.; Goldenberg, D.D.; Williams, O.; Loewith, R.; Stokoe, D.; Balla, A.; Toth, B.; et al. A pharmacological map of the PI3-K family defines a role for p110alpha in insulin signaling. Cell 2006, 125, 733–747. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arita, M.; Fuchino, H. Characterization of Anti-Poliovirus Compounds Isolated from Edible Plants. Viruses 2023, 15, 903. https://doi.org/10.3390/v15040903
Arita M, Fuchino H. Characterization of Anti-Poliovirus Compounds Isolated from Edible Plants. Viruses. 2023; 15(4):903. https://doi.org/10.3390/v15040903
Chicago/Turabian StyleArita, Minetaro, and Hiroyuki Fuchino. 2023. "Characterization of Anti-Poliovirus Compounds Isolated from Edible Plants" Viruses 15, no. 4: 903. https://doi.org/10.3390/v15040903