
Large-Scale Application of Double-Stranded RNA Shows Potential for Reduction of Sacbrood Virus Disease in Apis cerana Apiaries
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Sacbrood Virus
2.2. Extraction of Viral RNA
2.3. Construction of SBV Recombinant Plasmid
2.4. Production of Anti-SBV dsRNA
2.5. Real-Time Reverse Transcription PCR for Evaluation of SBV Inhibition by dsRNA
2.6. Efficacy of dsRNA In Vitro
2.7. Field Efficacy
2.8. Data Analysis
3. Results
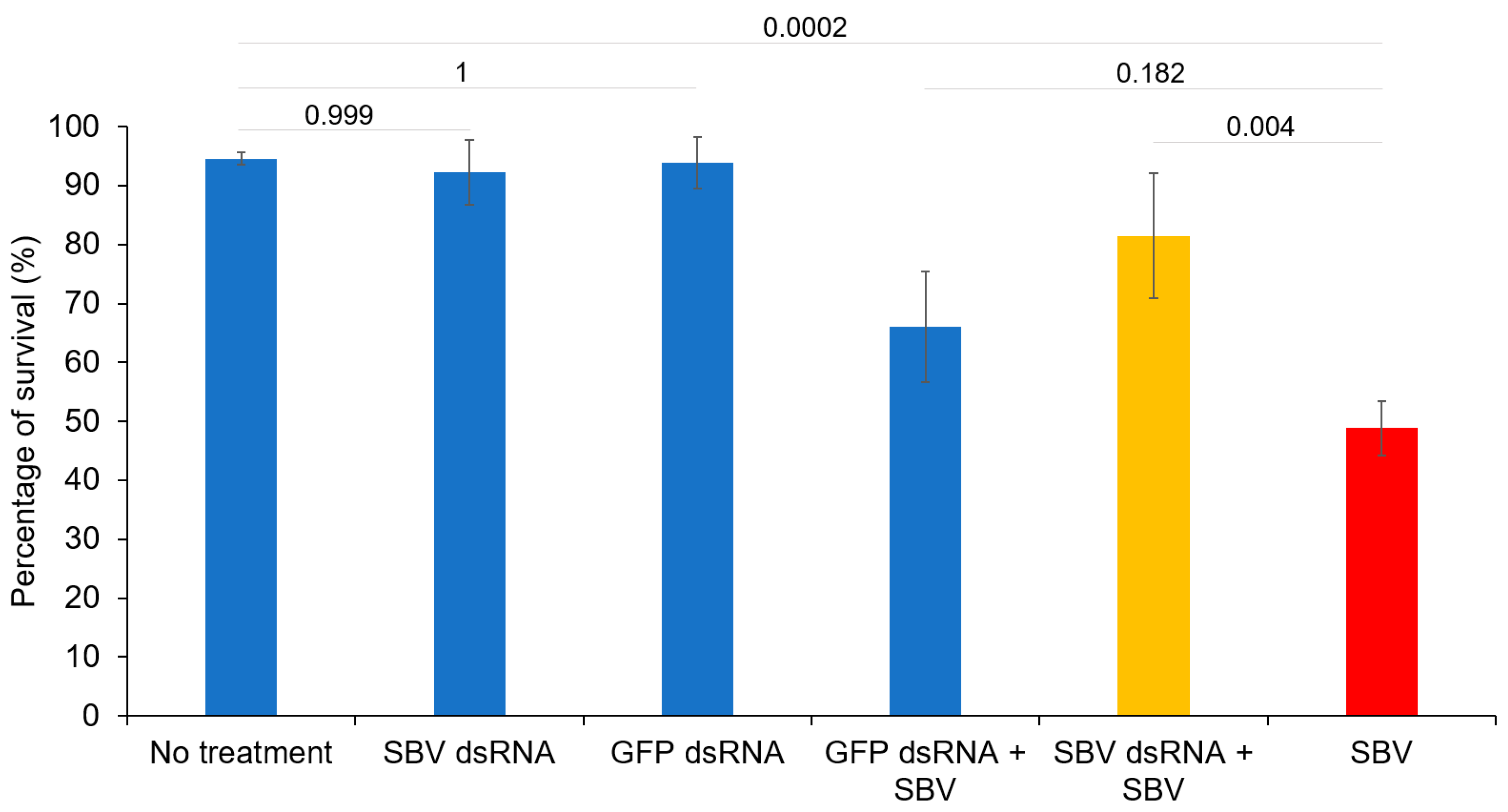
3.1. Efficacy of dsRNA In Vitro
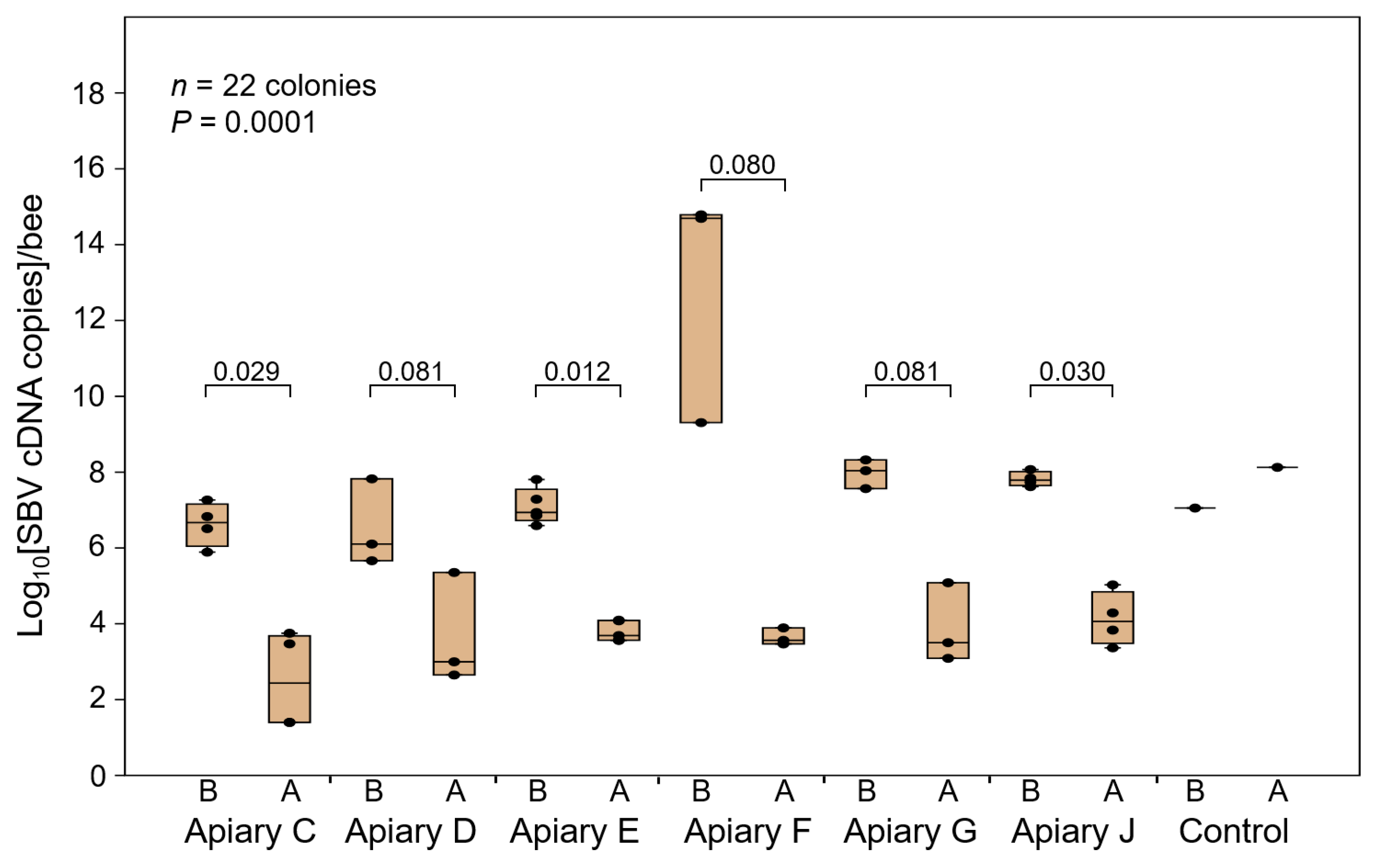
3.2. Field Application of dsRNA for the Prevention and Treatment of SBV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, R.C.; Ball, B.V.; Willcocks, M.M.; Carter, M.J. The Nucleotide Sequence of Sacbrood Virus of the Honey Bee: An Insect Picorna-like Virus. J. Gen. Virol. 1999, 80 Pt 6, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L. Recent Research on Honey Bee Viruses. Bee World 1975, 56, 55–64. [Google Scholar] [CrossRef]
- Bailey, L.; Fernando, E.F.W. Effects of Sacbrood Virus on Adult Honey-Bees. Ann. Appl. Biol. 1972, 72, 27–35. [Google Scholar] [CrossRef]
- Bailey, L.; Gibbs, A.J.; Woods, R.D. Sacbrood Virus of the Larval Honey Bee (Apis mellifera Linnaeus). Virology 1964, 23, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.L.; Gibbs, A.J. Transpuparial Transmission of Kashmir Bee Virus and Sacbrood Virus in the Honey Bee (Apis mellifera). Ann. Appl. Biol. 1989, 114, 1–7. [Google Scholar] [CrossRef]
- Anderson, D.L.; Giacon, H. Reduced Pollen Collection by Honey Bee (Hymenoptera: Apidae) Colonies Infected with Nosema apis and Sacbrood Virus. J. Econ. Entomol. 1992, 85, 47–51. [Google Scholar] [CrossRef]
- Wei, R.; Cao, L.; Feng, Y.; Chen, Y.; Chen, G.; Zheng, H. Sacbrood Virus: A Growing Threat to Honey Bees and Wild Pollinators. Viruses 2022, 14, 1871. [Google Scholar] [CrossRef]
- Ahn, A.J.; Ahn, K.S.; Suh, G.H.; Noh, J.H.; Kim, Y.H.; Yoo, M.S.; Kang, S.W.; Shin, S.S. Efficacy of Silver Ions against Sacbrood Virus Infection in the Eastern Honey Bee Apis cerana. J. Vet. Sci. 2015, 16, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.S.; Lee, M.Y.; Hong, I.P.; Kim, N.S.; Kim, H.K.; Lee, K.G.; Lee, M.L. Occurrence of Sacbrood Virus in Korean Apiaries from Apis cerana (Hymenoptera: Apidae). Korean J. Apic. 2010, 25, 187–191. [Google Scholar]
- Liu, X.; Zhang, Y.; Yan, X.; Han, R. Prevention of Chinese Sacbrood Virus Infection in Apis cerana Using RNA Interference. Curr. Microbiol. 2010, 61, 422–428. [Google Scholar] [CrossRef]
- Ma, M. New Insights of Sacbrood Virus. Virol. Sin. 2014, 29, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Liuhao, W.; Jun, G.; Yujie, T.; Yanping, C.; Jie, W.; Jilian, L. Chinese Sacbrood Virus Infection in Asian Honey Bees (Apis cerana cerana) and Host Immune Responses to the Virus Infection. J. Invertebr. Pathol. 2017, 150, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Procházková, M.; Füzik, T.; Škubník, K.; Moravcová, J.; Ubiparip, Z.; Přidal, A.; Plevka, P. Virion Structure and Genome Delivery Mechanism of Sacbrood Honeybee Virus. Proc. Natl. Acad. Sci. USA 2018, 115, 7759–7764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, A.T.; Kim, J.M.; Lim, S.J.; Yoo, M.S.; Cho, Y.S.; Yoon, B.S. Development of Ultra-Rapid PCR System for Genotyping of Sacbrood Virus. Korean J. Apic. 2018, 33, 83–98. [Google Scholar] [CrossRef]
- Charpentier, G.; Vidau, C.; Ferdy, J.B.; Tabart, J.; Vetillard, A. Lethal and Sub-Lethal Effects of Thymol on Honeybee (Apis mellifera) Larvae Reared in vitro. Pest Manag. Sci. 2014, 70, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Vung, N.N.; Kim, I.; Lee, M.Y.; Kim, H.K.; Kim, D.W.; Choi, Y.S. Controlling Sacbrood Virus Disease in Apis cerana Colonies with Biological Methods in Korea. Korean J. Apic. 2018, 33, 283–295. [Google Scholar] [CrossRef]
- Saleh, M.C.; van Rij, R.P.; Hekele, A.; Gillis, A.; Foley, E.; O’Farrell, P.H.; Andino, R. The Endocytic Pathway Mediates Cell Entry of dsRNA to Induce RNAi Silencing. Nat. Cell Biol. 2006, 8, 793–802. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Broeck, J.V. Biological Mechanisms Determining the Success of RNA Interference in Insects. Int. Rev. Cell Mol. Biol. 2014, 312, 139–167. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Van Wielendaele, P.; Vanden Broeck, J. Scavenger Receptor-mediated Endocytosis Facilitates RNA Interference in the Desert Locust, Schistocerca gregaria. Insect Mol. Biol. 2014, 23, 320–329. [Google Scholar] [CrossRef]
- Siomi, H.; Siomi, M. On the Road to Reading the RNA-interference Code. Nature 2009, 457, 396–404. [Google Scholar] [CrossRef]
- Brutscher, L.M.; Flenniken, M.L. RNAi and Antiviral Defense in the Honey Bee. J. Immunol. Res. 2015, 2015, 941897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMenamin, A.J.; Flenniken, M.L. Recently Identified Bee Viruses and Their Impact on Bee Pollinators. Curr. Opin. Insect Sci. 2018, 26, 120–129. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, A.J.; Daughenbaugh, K.F.; Parekh, F.; Pizzorno, M.C.; Flenniken, M.L. Honey Bee and Bumble Bee Antiviral Defense. Viruses 2018, 10, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Virus and dsRNA-Triggered Transcriptional Responses Reveal Key Components of Honey Bee Antiviral Defense. Sci. Rep. 2017, 7, 6448. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.E.; Bradish, H.M.; Gatehouse, J.A.; Fitches, E.C. Systemic RNAi in the Small Hive Beetle Aethina tumida Murray (Coleoptera: Nitidulidae), a Serious Pest of the European Honey Bee Apis mellifera. Pest Manag. Sci. 2017, 73, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Garbian, Y.; Maori, E.; Kalev, H.; Shafir, S.; Sela, I. Bidirectional Transfer of RNAi between Honey Bee and Varroa destructor: Varroa Gene Silencing Reduces Varroa Population. PLoS Pathog. 2012, 8, e1003035. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Evans, J.D. RNAi in Treating Honey Bee Diseases. Bee Cult. 2012, 140, 27–29. [Google Scholar]
- Desai, S.D.; Eu, Y.J.; Whyard, S.; Currie, R.W. Reduction in Deformed Wing Virus Infection in Larval and Adult Honey Bees (Apis mellifera L.) by Double-stranded RNA Ingestion. Insect Mol. Biol. 2012, 21, 446–455. [Google Scholar] [CrossRef]
- Hunter, W.; Ellis, J.; van Engelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-scale Field Application of RNAi Technology Reducing Israeli Acute Paralysis Virus Disease in Honey Bees (Apis mellifera, Hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a Bee-affecting Virus Associated with Colony Collapse Disorder Can Be Silenced by dsRNA Ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Han, R. The High-throughput Production of dsRNA against Sacbrood Virus for Use in the Honey Bee Apis cerana (Hymenoptera: Apidae). Virus Genes 2016, 52, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Truong, A.-T.; Yoo, M.S.; Seo, S.K.; Hwang, T.J.; Yoon, S.S.; Cho, Y.S. Prevalence of Honey Bee Pathogens and Parasites in South Korea: A Five-year Surveillance Study from 2017 to 2021. Heliyon 2023, 9, e13494. [Google Scholar] [CrossRef] [PubMed]
- Truong, A.T.; Yoo, M.S.; Yun, B.R.; Kang, J.E.; Noh, J.; Hwang, T.J.; Seo, S.K.; Yoon, S.S.; Cho, Y.S. Prevalence and Pathogen Detection of Varroa and Tropilaelaps Mites in Apis mellifera (Hymenoptera, Apidae) Apiaries in South Korea. J. Apic. Res. 2022, 1–9. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Tukey’s Honestly Significant Difference (HSD) Test. Encycl. Res. Des. 2010, 3, 1–5. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Paul, D.R. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ullah, A.; Gajger, I.T.; Majoros, A.; Dar, S.A.; Khan, S.; Kalimullah; Shah, A.H.; Khabir, M.N.; Hussain, R.; Khan, H.U.; et al. Viral Impacts on Honey Bee Populations: A Review. Saudi J. Biol. Sci. 2021, 28, 523–530. [Google Scholar] [CrossRef]
- Remnant, E.J.; Shi, M.; Buchmann, G.; Blacquière, T.; Holmes, E.C.; Beekman, M.; Ashe, A. A Diverse Range of Novel RNA Viruses in Geographically Distinct Honey Bee Populations. J. Virol. 2017, 91, e00158-17. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y. Viruses and Viral Diseases of the Honey Bee, Apis mellifera. In Recent Advances in Entomological Research; Liu, T., Kang, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 105–120. ISBN 9783642178146. [Google Scholar]
- Bailey, L. The Multiplication and Spread of Sacbrood Virus of Bees. Ann. Appl. Biol. 1969, 63, 483–491. [Google Scholar] [CrossRef]
- Gauthier, L.; Tentcheva, D.; Tournaire, M.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Viral Load Estimation in Asymptomatic Honey Bee Colonies Using the Quantitative RT-PCR Technique. Apidologie 2007, 38, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Maori, E.; Garbian, Y.; Kunik, V.; Mozes-Koch, R.; Malka, O.; Kalev, H.; Sabath, N.; Sela, I.; Shafir, S. A Transmissible RNA Pathway in Honey Bees. Cell Rep. 2019, 27, 1949–1959.e1946. [Google Scholar] [CrossRef] [Green Version]
- Chejanovsky, N.; Ophir, R.; Schwager, M.S.; Slabezki, Y.; Grossman, S.; Cox-Foster, D. Characterization of Viral siRNA Populations in Honey Bee Colony Collapse Disorder. Virology 2014, 454–455, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Groups | Application Routes (Concentration) | Apiary No. | Colony No. | No. of Colonies with SBV Symptoms (%) | SBV Presence 1 | Surviving Colonies 2 |
|---|---|---|---|---|---|---|
| dsRNA treatment | Spray (10 mg) | 5 | 9 | 0 (0) | + | 9 |
| Oral (10 mg) | 5 | 9 | 0 (0) | + | 9 | |
| Oral (20 mg) | 7 | 60 | 0 (0) | + | 60 | |
| Subtotal | 17 | 78 | 0 (0) | + | 78 | |
| Negative control | No treatment | 4 | 7 | 3 (42) | + | 4 |
| Groups | Before Administration | Apiaries (Hives) | Administration Interval (Week(s)) | After Administration | Results | Efficacy |
|---|---|---|---|---|---|---|
| Prevention | Infected; no disease symptoms | 1 (33) | 4 | Healthy | Healthy and split | Effective |
| 16 (134) | 2 | Healthy | Healthy and split | Effective | ||
| Treatment | Disease symptoms | 4 (18) | 4 | Pulled out larvae | Colony loss within 2 months (100%) | Not effective |
| 11 (78) | 2 | Pulled out larvae | Survival for more than 2 months but finally loss (100%) | Not effective | ||
| 1 (6) | 1 | Less pulled out larvae | Keeping colony until 8 months; finally loss (100%) | Partially effective | ||
| Total | 33 (269) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, M.-S.; Truong, A.-T.; Jeong, H.; Hahn, D.-H.; Lee, J.-S.; Yoon, S.-S.; Youn, S.-Y.; Cho, Y.-S. Large-Scale Application of Double-Stranded RNA Shows Potential for Reduction of Sacbrood Virus Disease in Apis cerana Apiaries. Viruses 2023, 15, 897. https://doi.org/10.3390/v15040897
Yoo M-S, Truong A-T, Jeong H, Hahn D-H, Lee J-S, Yoon S-S, Youn S-Y, Cho Y-S. Large-Scale Application of Double-Stranded RNA Shows Potential for Reduction of Sacbrood Virus Disease in Apis cerana Apiaries. Viruses. 2023; 15(4):897. https://doi.org/10.3390/v15040897
Chicago/Turabian StyleYoo, Mi-Sun, A-Tai Truong, Hana Jeong, Do-Hyun Hahn, Ju-Seong Lee, Soon-Seek Yoon, So-Youn Youn, and Yun-Sang Cho. 2023. "Large-Scale Application of Double-Stranded RNA Shows Potential for Reduction of Sacbrood Virus Disease in Apis cerana Apiaries" Viruses 15, no. 4: 897. https://doi.org/10.3390/v15040897