
CD8+ Lymphocytes from Healthy Blood Donors Secrete Antiviral Levels of Interferon-Alpha
Abstract
:1. Introduction
2. Methods
3. Results
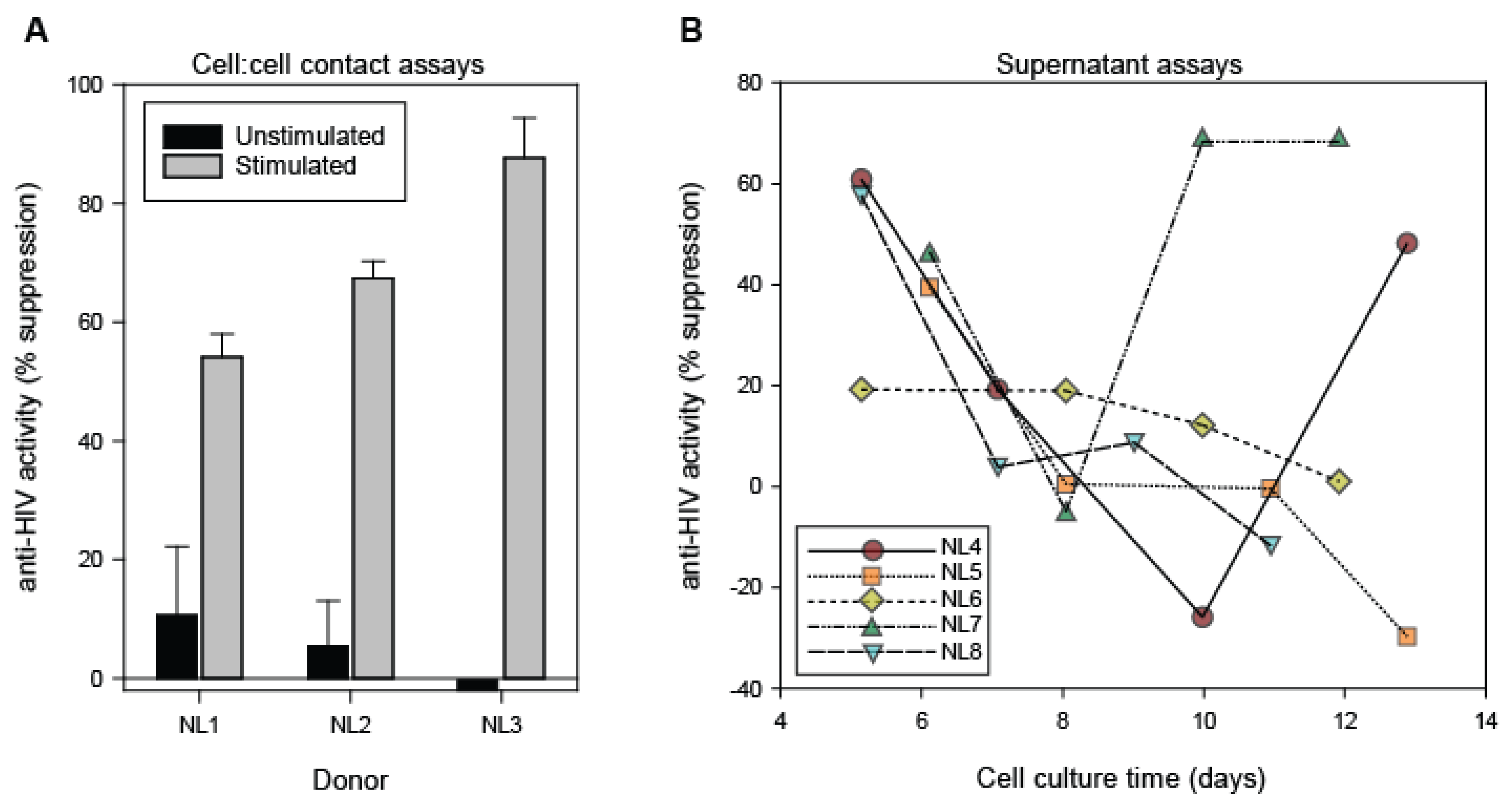
3.1. Antiviral Activity of CD8+ T Cells and CD8+ T Cell Fluids from Healthy Blood Donors
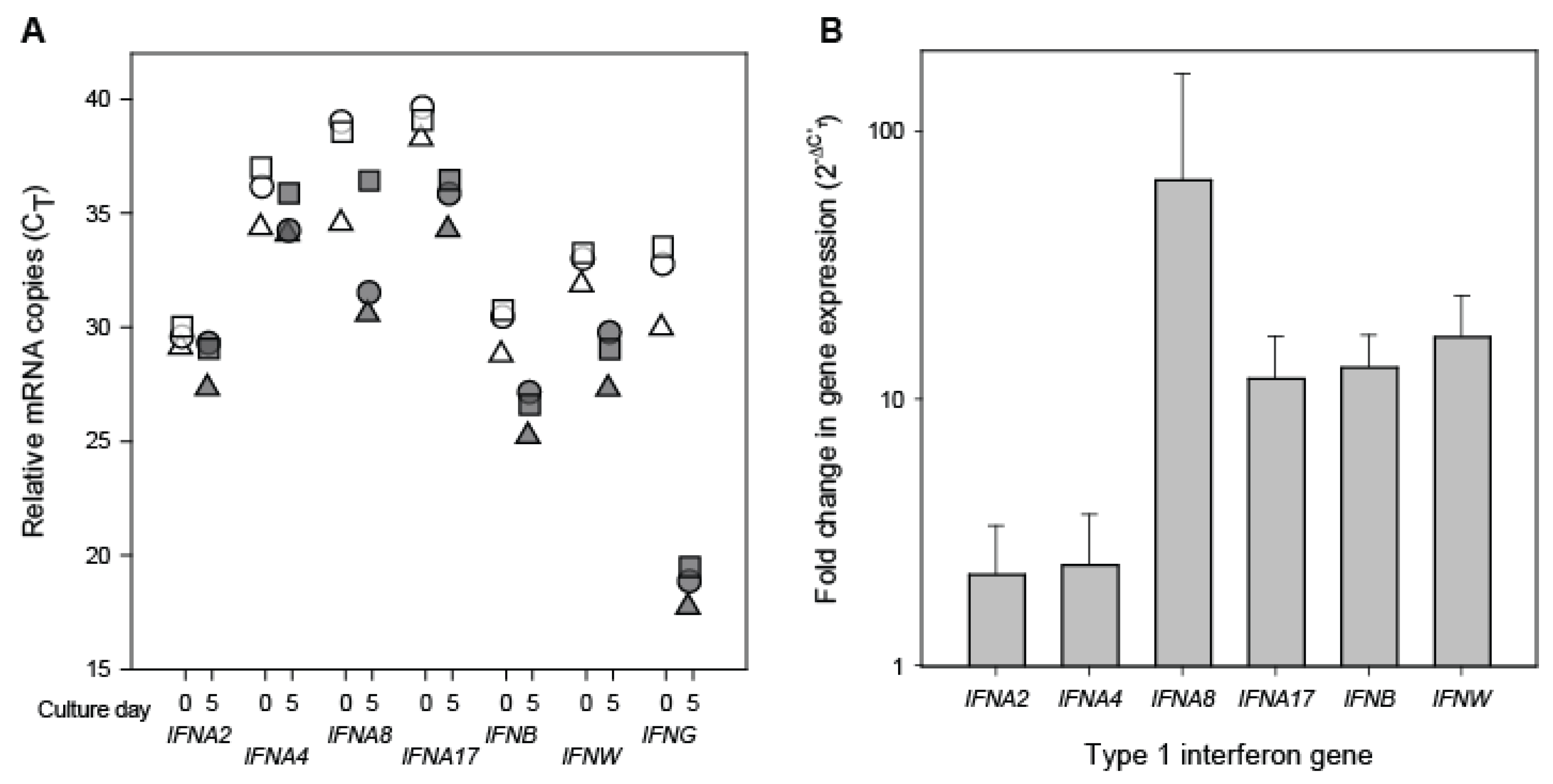
3.2. Interferon Transcript Levels in Resting and Stimulated CD8+ Cells
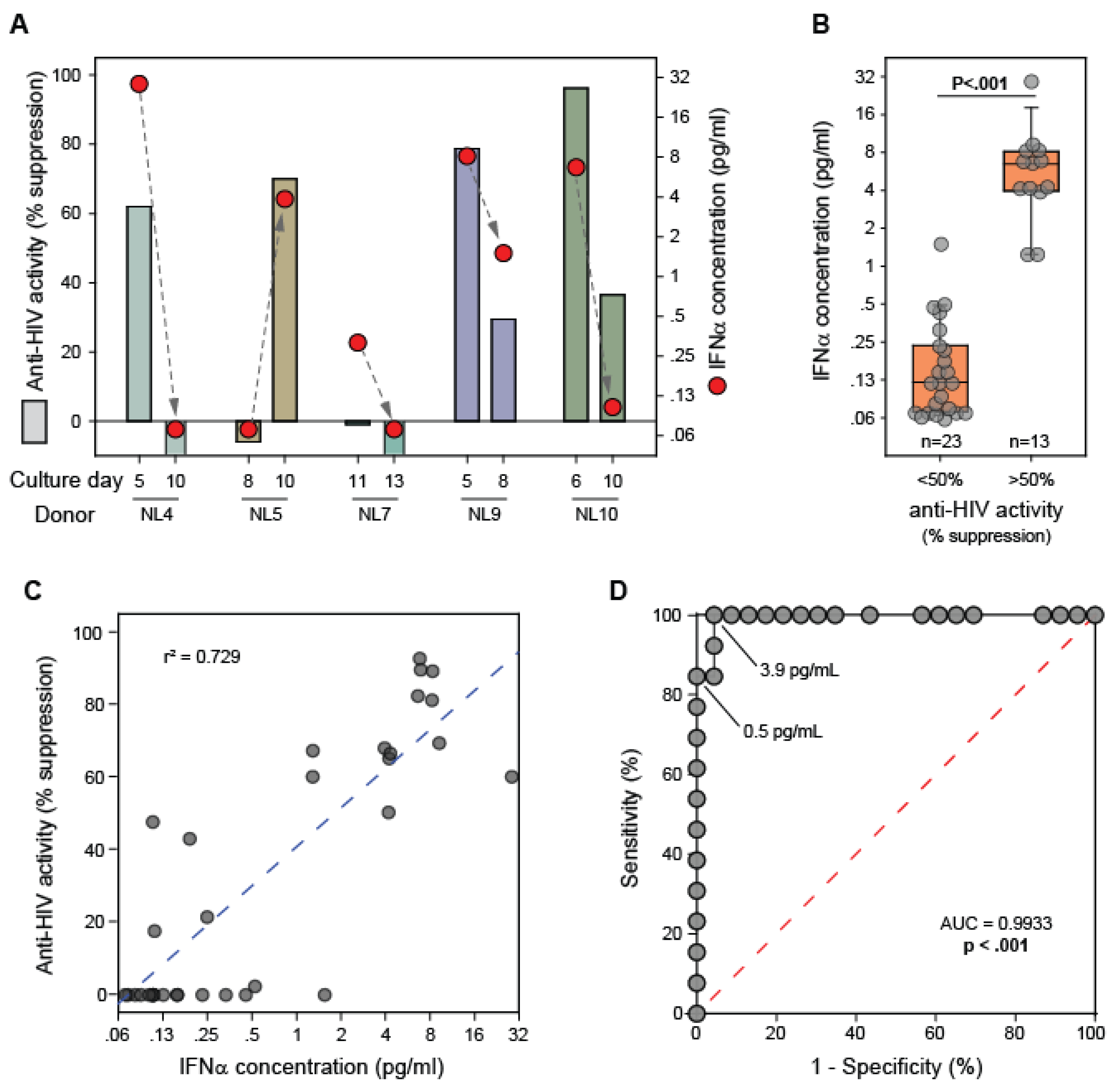
3.3. IFNα Levels in CD8+ T Cell Fluids from Healthy Blood Donors
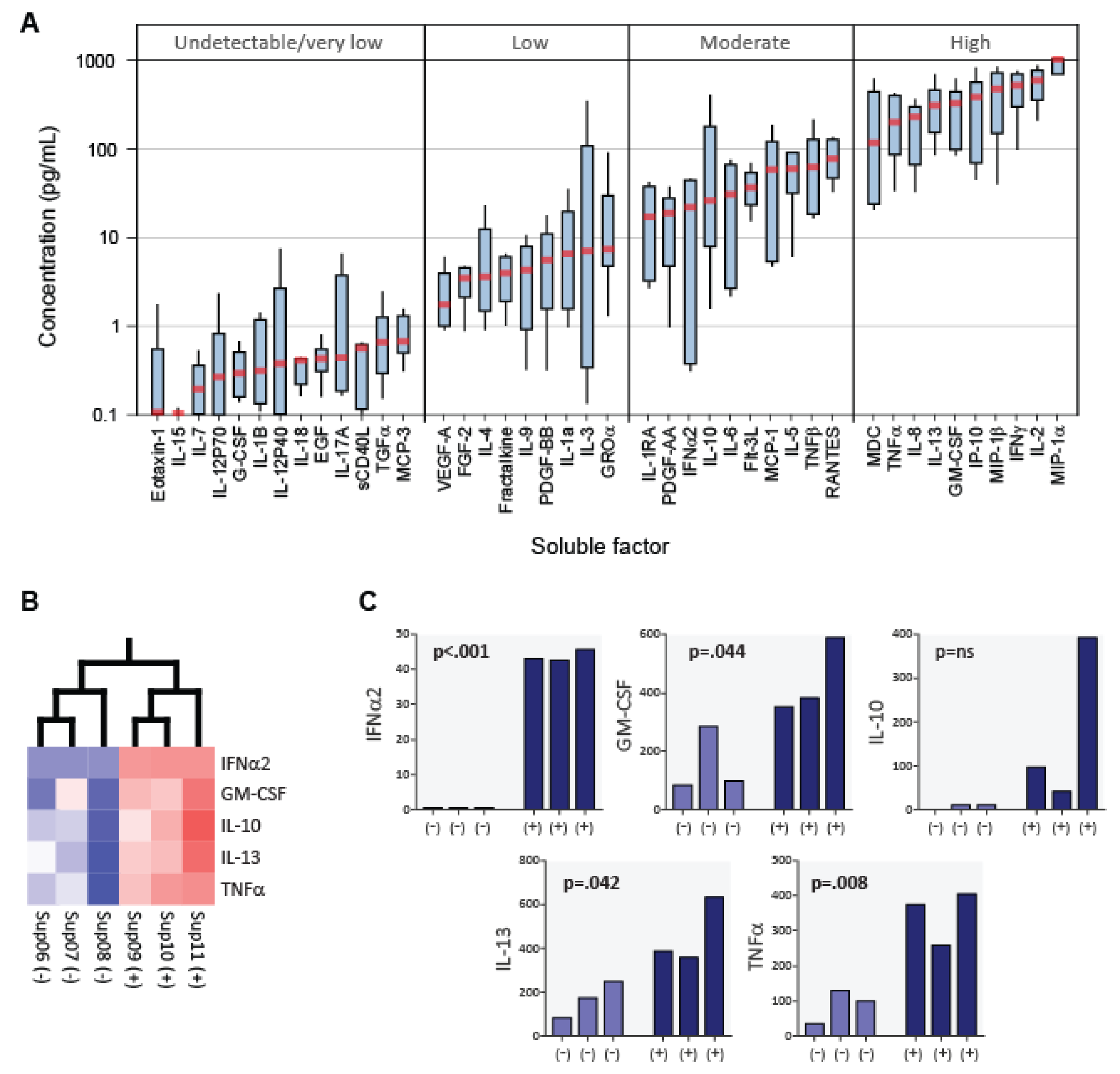
3.4. Cytokine Profiles of CD8+ T Cell Fluids from Healthy Blood Donors
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richard, A.C. Divide and Conquer: Phenotypic and Temporal Heterogeneity Within CD8(+) T Cell Responses. Front. Immunol. 2022, 13, 949423. [Google Scholar] [CrossRef]
- Al Moussawy, M.; Abdelsamed, H.A. Non-cytotoxic functions of CD8 T cells: “Repentance of a serial killer”. Front. Immunol. 2022, 13, 1001129. [Google Scholar] [CrossRef]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. CD8(+) T cells in HIV control, cure and prevention. Nat. Rev. Immunol. 2020, 20, 471–482. [Google Scholar] [CrossRef]
- Jin, X.; Bauer, D.E.; Tuttleton, S.E.; Lewin, S.; Gettie, A.; Blanchard, J.; Irwin, C.E.; Safrit, J.T.; Mittler, J.; Weinberger, L.; et al. Dramatic rise in plasma viremia after CD8(+) T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 1999, 189, 991–998. [Google Scholar] [CrossRef]
- Saez-Cirion, A.; Lacabaratz, C.; Lambotte, O.; Versmisse, P.; Urrutia, A.; Boufassa, F.; Barre-Sinoussi, F.; Delfraissy, J.F.; Sinet, M.; Pancino, G.; et al. HIV controllers exhibit potent CD8 T cell capacity to suppress HIV infection ex vivo and peculiar cytotoxic T lymphocyte activation phenotype. Proc. Natl. Acad. Sci. USA 2007, 104, 6776–6781. [Google Scholar] [CrossRef]
- Pereyra, F.; Jia, X.; McLaren, P.J.; Telenti, A.; de Bakker, P.I.; Walker, B.D.; Ripke, S.; Brumme, C.J.; Pulit, S.L.; Carrington, M.; et al. The major genetic determinants of HIV-1 control affect HLA class I peptide presentation. Science 2010, 330, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Andrieu, J.M.; Lu, W. Evidence of a tolerogenic vaccine against AIDS in the Chinese macaque prefigures a potential human vaccine. Arch. Virol. 2021, 166, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, J.V.; Lyles, R.H.; Matud, J.L.; Yamashita, T.E.; Mellors, J.W.; Hultin, L.E.; Jamieson, B.D.; Margolick, J.B.; Rinaldo, C.R., Jr.; Phair, J.P.; et al. Predictive value of immunologic and virologic markers after long or short duration of HIV-1 infection. J. Acquir. Immune. Defic. Syndr. 2002, 29, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.M.; Moody, D.J.; Stites, D.P.; Levy, J.A. CD8+ lymphocytes can control HIV infection in vitro by suppressing virus replication. Science 1986, 234, 1563–1566. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.D.; Chakrabarti, S.; Moss, B.; Paradis, T.J.; Flynn, T.; Durno, A.G.; Blumberg, R.S.; Kaplan, J.C.; Hirsch, M.S.; Schooley, R.T. HIV-specific cytotoxic T lymphocytes in seropositive individuals. Nature 1987, 328, 345–348. [Google Scholar] [CrossRef]
- McMichael, A.J. Is a Human CD8 T-Cell Vaccine Possible, and if So, What Would It Take? Could a CD8(+) T-Cell Vaccine Prevent Persistent HIV Infection? Cold Spring Harb. Perspect. Biol. 2018, 10, a029124. [Google Scholar] [CrossRef]
- Lee, J.H.; Crotty, S. HIV vaccinology: 2021 update. Semin. Immunol. 2021, 51, 101470. [Google Scholar] [CrossRef]
- Barouch, D.H.; Tomaka, F.L.; Wegmann, F.; Stieh, D.J.; Alter, G.; Robb, M.L.; Michael, N.L.; Peter, L.; Nkolola, J.P.; Borducchi, E.N.; et al. Evaluation of a mosaic HIV-1 vaccine in a multicentre, randomised, double-blind, placebo-controlled, phase 1/2a clinical trial (APPROACH) and in rhesus monkeys (NHP 13-19). Lancet 2018, 392, 232–243. [Google Scholar] [CrossRef]
- Moriuchi, H.; Moriuchi, M.; Combadiere, C.; Murphy, P.M.; Fauci, A.S. CD8+ T-cell-derived soluble factor(s), but not beta-chemokines RANTES, MIP-1 alpha, and MIP-1 beta, suppress HIV-1 replication in monocyte/macrophages. Proc. Natl. Acad. Sci. USA 1996, 93, 15341–15345. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, W.; He, T.; Yu, J.; Caffrey, R.E.; Dalmasso, E.A.; Fu, S.; Pham, T.; Mei, J.; Ho, J.J.; et al. Contribution of human alpha-defensin 1, 2, and 3 to the anti-HIV-1 activity of CD8 antiviral factor. Science 2002, 298, 995–1000. [Google Scholar] [CrossRef]
- Cocchi, F.; DeVico, A.L.; Garzino-Demo, A.; Arya, S.K.; Gallo, R.C.; Lusso, P. Identification of RANTES, MIP-1 alpha, and MIP-1 beta as the major HIV-suppressive factors produced by CD8+ T cells. Science 1995, 270, 1811–1815. [Google Scholar] [CrossRef]
- Levy, J.A. The search for the CD8+ cell anti-HIV factor (CAF). Trends Immunol. 2003, 24, 628–632. [Google Scholar] [CrossRef]
- Blazek, D.; Teque, F.; Mackewicz, C.; Peterlin, M.; Levy, J.A. The CD8+ cell non-cytotoxic antiviral response affects RNA polymerase II-mediated human immunodeficiency virus transcription in infected CD4+ cells. J. Gen. Virol. 2016, 97, 220–224. [Google Scholar] [CrossRef]
- Ye, P.; Kazanjian, P.; Kunkel, S.L.; Kirschner, D.E. Lack of good correlation of serum CC-chemokine levels with human immunodeficiency virus-1 disease stage and response to treatment. J. Lab. Clin. Med. 2004, 143, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.S.; Teque, F.; Walker, R.L.; Meltzer, P.S.; Killian, J.K. CD8(+) lymphocytes suppress human immunodeficiency virus 1 replication by secreting type I interferons. J. Interferon Cytokine Res. 2013, 33, 632–645. [Google Scholar] [CrossRef]
- Wheelock, E.F. Interferon-Like Virus-Inhibitor Induced in Human Leukocytes by Phytohemagglutinin. Science 1965, 149, 310–311. [Google Scholar] [CrossRef]
- Roff, S.R.; Noon-Song, E.N.; Yamamoto, J.K. The Significance of Interferon-gamma in HIV-1 Pathogenesis, Therapy, and Prophylaxis. Front. Immunol. 2014, 4, 498. [Google Scholar] [CrossRef]
- Schlaepfer, E.; Fahrny, A.; Gruenbach, M.; Kuster, S.P.; Simon, V.; Schreiber, G.; Speck, R.F. Dose-Dependent Differences in HIV Inhibition by Different Interferon Alpha Subtypes While Having Overall Similar Biologic Effects. mSphere 2019, 4, e00637-18. [Google Scholar] [CrossRef]
- Tavel, J.A.; Huang, C.Y.; Shen, J.; Metcalf, J.A.; Dewar, R.; Shah, A.; Vasudevachari, M.B.; Follmann, D.A.; Herpin, B.; Davey, R.T.; et al. Interferon-alpha produces significant decreases in HIV load. J. Interferon Cytokine Res. 2010, 30, 461–464. [Google Scholar] [CrossRef]
- Pitha, P.M. Multiple effects of interferon on the replication of human immunodeficiency virus type 1. Antivir. Res. 1994, 24, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Sivro, A.; Su, R.C.; Plummer, F.A.; Ball, T.B. Interferon responses in HIV infection: From protection to disease. AIDS Rev. 2014, 16, 43–51. [Google Scholar] [PubMed]
- Soper, A.; Kimura, I.; Nagaoka, S.; Konno, Y.; Yamamoto, K.; Koyanagi, Y.; Sato, K. Type I Interferon Responses by HIV-1 Infection: Association with Disease Progression and Control. Front. Immunol. 2017, 8, 1823. [Google Scholar] [CrossRef]
- Siegel, J.P. Effects of interferon-gamma on the activation of human T lymphocytes. Cell Immunol. 1988, 111, 461–472. [Google Scholar] [CrossRef]
- Marrack, P.; Kappler, J.; Mitchell, T. Type I interferons keep activated T cells alive. J. Exp. Med. 1999, 189, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Yamada, O.; Hattori, N.; Kurimura, T.; Kita, M.; Kishida, T. Inhibition of growth of HIV by human natural interferon in vitro. AIDS Res. Hum. Retrovir. 1988, 4, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L.; Neumeyer, D.; Vogt, M.W.; Schooley, R.T.; Hirsch, M.S. Activity of interferons alpha, beta, and gamma against human immunodeficiency virus replication in vitro. AIDS Res. Hum. Retrovir. 1987, 3, 125–133. [Google Scholar] [CrossRef]
- Wegehaupt, A.K.; Roufs, E.K.; Hewitt, C.R.; Killian, M.L.; Gorbatenko, O.; Anderson, C.M.; Killian, M.S. Recovery and assessment of leukocytes from LR Express filters. Biologicals 2017, 49, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.S.; Johnson, C.; Teque, F.; Fujimura, S.; Levy, J.A. Natural suppression of human immunodeficiency virus type 1 replication is mediated by transitional memory CD8+ T cells. J. Virol. 2011, 85, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.S.; Ng, S.; Mackewicz, C.E.; Levy, J.A. A screening assay for detecting CD8+ cell non-cytotoxic anti-HIV responses. J. Immunol. Methods 2005, 304, 137–150. [Google Scholar] [CrossRef]
- Mackewicz, C.E.; Barker, E.; Greco, G.; Reyes-Teran, G.; Levy, J.A. Do beta-chemokines have clinical relevance in HIV infection? J. Clin. Invest. 1997, 100, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Remoli, M.E.; Giacomini, E.; Lutfalla, G.; Dondi, E.; Orefici, G.; Battistini, A.; Uze, G.; Pellegrini, S.; Coccia, E.M. Selective expression of type I IFN genes in human dendritic cells infected with Mycobacterium tuberculosis. J. Immunol. 2002, 169, 366–374. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Walker, C.M.; Levy, J.A. A diffusible lymphokine produced by CD8+ T lymphocytes suppresses HIV replication. Immunology 1989, 66, 628–630. [Google Scholar]
- Brandt, E.R.; Linnane, A.W.; Devenish, R.J. Expression of IFN A genes in subpopulations of peripheral blood cells. Br. J. Haematol. 1994, 86, 717–725. [Google Scholar] [CrossRef]
- Matsuyama, M.; Sairenji, T.; Yonemura, K.; Hinuma, Y. Interferon production potentials of various human lymphoblastoid cell lines. Microbiol. Immunol. 1982, 26, 1149–1158. [Google Scholar] [CrossRef]
- Adolf, G.R.; Swetly, P. Interferon production in human hematopoietic cell lines: Response to chemicals and characterization of interferons. J. Interferon. Res. 1982, 2, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Greenway, A.L.; Hertzog, P.J.; Devenish, R.J.; Linnane, A.W. Constitutive and virus-induced interferon production by peripheral blood leukocytes. Exp. Hematol. 1995, 23, 229–235. [Google Scholar] [PubMed]
- Buckheit, R.W., 3rd; Salgado, M.; Silciano, R.F.; Blankson, J.N. Inhibitory potential of subpopulations of CD8+ T cells in HIV-1-infected elite suppressors. J. Virol. 2012, 86, 13679–13688. [Google Scholar] [CrossRef] [PubMed]
- Barker, E. CD8+ cell-derived anti-human immunodeficiency virus inhibitory factor. J. Infect. Dis. 1999, 179 (Suppl. S3), S485–S488. [Google Scholar] [CrossRef] [PubMed]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Lacey, S.F.; McDanal, C.B.; Ferrari, G.; Weinhold, K.J.; Greenberg, M.L. CD8+ T cell-mediated suppressive activity inhibits HIV-1 after virus entry with kinetics indicating effects on virus gene expression. Proc. Natl. Acad. Sci. USA 2000, 97, 3503–3508. [Google Scholar] [CrossRef] [PubMed]
- Morvan, M.G.; Teque, F.C.; Locher, C.P.; Levy, J.A. The CD8(+) T Cell Noncytotoxic Antiviral Responses. Microbiol. Mol. Biol. Rev. 2021, 85, e00155-20. [Google Scholar] [CrossRef]
- Shridhar, V.; Chen, Y.; Gupta, P. The CD8 antiviral factor (CAF) can suppress HIV-1 transcription from the long terminal repeat (LTR) promoter in the absence of elements upstream of the CATATAA box. Virol. J. 2014, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, M.; Palesch, D.; Pinacchio, C.; Statzu, M.; Tharp, G.K.; Paiardini, M.; Chahroudi, A.; Bosinger, S.E.; Yoon, J.; Cox, B.; et al. Innate, non-cytolytic CD8+ T cell-mediated suppression of HIV replication by MHC-independent inhibition of virus transcription. PLoS Pathog. 2020, 16, e1008821. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.; et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef]
- Mackewicz, C.E.; Ortega, H.; Levy, J.A. Effect of cytokines on HIV replication in CD4+ lymphocytes: Lack of identity with the CD8+ cell antiviral factor. Cell Immunol. 1994, 153, 329–343. [Google Scholar] [CrossRef]
- Koubek, K.; Stary, J.; Kumberova, A.; Klamova, H.; Filipec, M. Occurrence of cytokine receptors on different lymphoid leukaemic cells. Eur. J. Haematol. 1999, 63, 1–10. [Google Scholar] [CrossRef]
- Kootstra, N.A.; van‘t Wout, A.; Huisman, H.G.; Miedema, F.; Schuitemaker, H. Interference of interleukin-10 with human immunodeficiency virus type 1 replication in primary monocyte-derived macrophages. J. Virol. 1994, 68, 6967–6975. [Google Scholar] [CrossRef]
- Montaner, L.J.; Doyle, A.G.; Collin, M.; Herbein, G.; Illei, P.; James, W.; Minty, A.; Caput, D.; Ferrara, P.; Gordon, S. Interleukin 13 inhibits human immunodeficiency virus type 1 production in primary blood-derived human macrophages in vitro. J. Exp. Med. 1993, 178, 743–747. [Google Scholar] [CrossRef]
- Poli, G.; Fauci, A.S. The effect of cytokines and pharmacologic agents on chronic HIV infection. AIDS Res. Hum. Retrovir. 1992, 8, 191–197. [Google Scholar] [CrossRef]
- Killian, M.S.; Fujimura, S.H.; Sudhagoni, R.G. Increased Expression of the Type I Interferon Receptor on CD4+ T Lymphocytes in HIV-1 Infected Individuals. J. Acquir. Immune Defic. Syndr. 2016, 74, 473–478. [Google Scholar] [CrossRef]
- Crow, M.K.; Ronnblom, L. Type I interferons in host defence and inflammatory diseases. Lupus Sci. Med. 2019, 6, e000336. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Type 1 Interferon (IFNα, β, ι) | Type 2 Interferon (IFNγ) | References |
|---|---|---|---|
| Secreted by CD8+ T cells from healthy HIV-infected subjects | + | + | [20,22] |
| Associated with HIV-1 disease progression | + | + | [22,26] |
| Associated with prevention of HIV infection | + | + | [22,27] |
| Affects CD4+ T cell growth in vitro | − | − | [28,29] |
| Affects HIV transcription | + | − | [27] |
| Affects all HIV isolates tested | + | − | [20,30,31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teque, F.; Wegehaupt, A.; Roufs, E.; Killian, M.S. CD8+ Lymphocytes from Healthy Blood Donors Secrete Antiviral Levels of Interferon-Alpha. Viruses 2023, 15, 894. https://doi.org/10.3390/v15040894
Teque F, Wegehaupt A, Roufs E, Killian MS. CD8+ Lymphocytes from Healthy Blood Donors Secrete Antiviral Levels of Interferon-Alpha. Viruses. 2023; 15(4):894. https://doi.org/10.3390/v15040894
Chicago/Turabian StyleTeque, Fernando, Abby Wegehaupt, Ellen Roufs, and M. Scott Killian. 2023. "CD8+ Lymphocytes from Healthy Blood Donors Secrete Antiviral Levels of Interferon-Alpha" Viruses 15, no. 4: 894. https://doi.org/10.3390/v15040894