
Positive Selection and Duplication of Bat TRIM Family Proteins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bat Genome Data Retrieval
2.2. Identification of TRIM Proteins from Bats
2.3. Phylogeny and Motif Analysis
2.4. Positive Selection Analysis
2.5. RNA-Seq Data Analysis
3. Results
3.1. Number of Bat TRIM Family Members
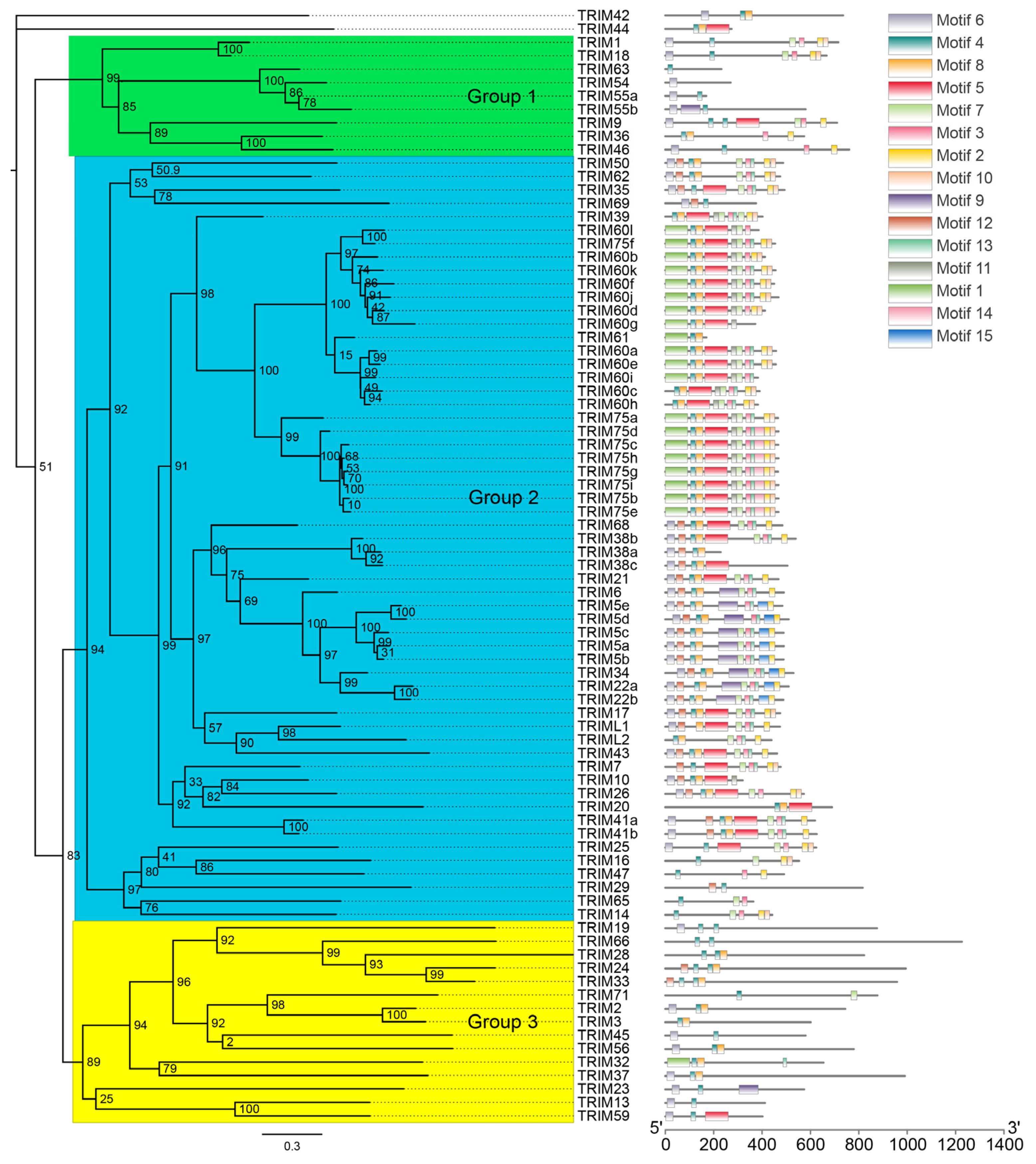
3.2. Phylogeny and Motif Distribution of Bat TRIM Family Proteins
3.3. Positive Selection Analysis of Bat TRIM Family Genes
3.4. Tissue Expression Analysis of Bat TRIM Genes
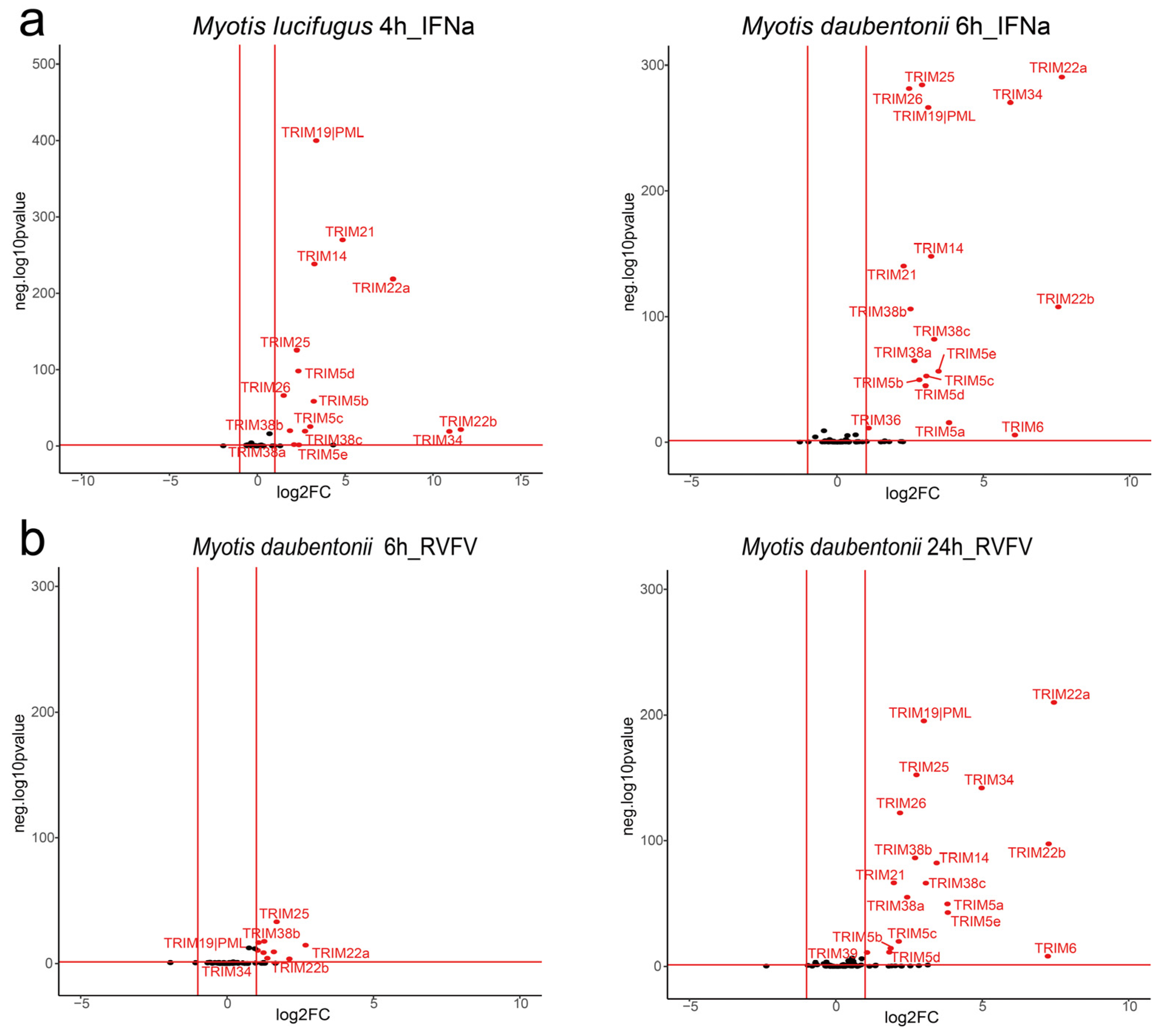
3.5. Bat TRIM Genes Regulated by Virus Infection or Type I IFN Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moratelli, R.; Calisher, C.H. Bats and zoonotic viruses: Can we confidently link bats with emerging deadly viruses? Mem. Do Inst. Oswaldo Cruz 2015, 110, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.F.; Shi, Z.L.; Zhang, S.Y.; Field, H.; Daszak, P.; Eaton, B.T. Review of bats and SARS. Emerg. Infect. Dis. 2006, 12, 1834–1840. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef]
- Wang, L.-F.; Gamage, A.M.; Chan, W.O.Y.; Hiller, M.; Teeling, E.C. Decoding bat immunity: The need for a coordinated research approach. Nat. Rev. Immunol. 2021, 21, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Teeling, E.C.; Vernes, S.C.; Dávalos, L.M.; Ray, D.A.; Gilbert, M.T.P.; Myers, E. Bat Biology, Genomes, and the Bat1K Project: To Generate Chromosome-Level Genomes for All Living Bat Species. Annu. Rev. Anim. Biosci. 2018, 6, 23–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Tachedjian, M.; Wynne, J.W.; Boyd, V.; Cui, J.; Smith, I.; Cowled, C.; Ng, J.H.J.; Mok, L.; Michalski, W.P.; et al. Contraction of the type I IFN locus and unusual constitutive expression of IFN-alpha in bats. Proc. Natl. Acad. Sci. USA 2016, 113, 2696–2701. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Li, Y.; Shen, X.; Goh, G.; Zhu, Y.; Cui, J.; Wang, L.-F.; Shi, Z.-L.; Zhou, P. Dampened STING-Dependent Interferon Activation in Bats. Cell Host Microbe 2018, 23, 297–301.e294. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.; Anderson, D.E.; Zhang, Q.; Tan, C.W.; Lim, B.L.; Luko, K.; Wen, M.; Chia, W.N.; Mani, S.; Wang, L.C.; et al. Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host. Nat. Microbiol. 2019, 4, 789–799. [Google Scholar] [CrossRef]
- Watanabe, M.; Hatakeyama, S. TRIM proteins and diseases. J. Biochem. 2017, 161, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Munir, M. TRIM Proteins: Another Class of Viral Victims. Sci. Signal. 2010, 3, jc2. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.-M.; Shu, H.-B. Multifaceted roles of TRIM38 in innate immune and inflammatory responses. Cell. Mol. Immunol. 2017, 14, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Gack, M.U.; Shin, Y.C.; Joo, C.-H.; Urano, T.; Liang, C.; Sun, L.; Takeuchi, O.; Akira, S.; Chen, Z.; Inoue, S.; et al. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 2007, 446, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Higgs, R.; Gabhann, J.N.; Ben Larbi, N.; Breen, E.P.; Fitzgerald, K.A.; Jefferies, C.A. The E3 ubiquitin ligase Ro52 negatively regulates IFN-beta production post-pathogen recognition by polyubiquitin-mediated degradation of IRF3. J. Immunol. 2008, 181, 1780–1786. [Google Scholar] [CrossRef] [Green Version]
- Barr, S.D.; Smiley, J.R.; Bushman, F.D. The interferon response inhibits HIV particle production by induction of TRIM22. PLoS Pathog. 2008, 4, e1000007. [Google Scholar] [CrossRef] [Green Version]
- Stremlau, M.; Perron, M.; Lee, M.; Li, Y.; Song, B.; Javanbakht, H.; Diaz-Griffero, F.; Anderson, D.J.; Sundquist, W.I.; Sodroski, J. Specific recognition and accelerated uncoating of retroviral capsids by the TRIM5alpha restriction factor. Proc. Natl. Acad. Sci. USA 2006, 103, 5514–5519. [Google Scholar] [CrossRef] [Green Version]
- Stremlau, M.; Perron, M.; Welikala, S.; Sodroski, J. Species-specific variation in the B30.2(SPRY) domain of TRIM5alpha determines the potency of human immunodeficiency virus restriction. J. Virol. 2005, 79, 3139–3145. [Google Scholar] [CrossRef] [Green Version]
- Rajsbaum, R.; Garcia-Sastre, A.; Versteeg, G.A. TRIMmunity: The Roles of the TRIM E3-Ubiquitin Ligase Family in Innate Antiviral Immunity. J. Mol. Biol. 2014, 426, 1265–1284. [Google Scholar] [CrossRef] [Green Version]
- Malfavon-Borja, R.; Sawyer, S.L.; Wu, L.I.; Emerman, M.; Malik, H.S. An Evolutionary Screen Highlights Canonical and Noncanonical Candidate Antiviral Genes within the Primate TRIM Gene Family. Genome Biol. Evol. 2013, 5, 2141–2154. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.P.; Águeda-Pinto, A.; Pinheiro, A.; Rebelo, H.; Esteves, P.J. Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process. Viruses 2022, 14, 345. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBIBLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2-Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Pond, S.L.K.; Frost, S.D.W.; Muse, S.V. HyPhy: Hypothesis testing using phylogenies. Bioinformatics 2005, 21, 676–679. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Amador, L.I.; Moyers Arevalo, R.L.; Almeida, F.C.; Catalano, S.A.; Giannini, N.P. Bat Systematics in the Light of Unconstrained Analyses of a Comprehensive Molecular Supermatrix. J. Mamm. Evol. 2018, 25, 37–70. [Google Scholar] [CrossRef]
- Sardiello, M.; Cairo, S.; Fontanella, B.; Ballabio, A.; Meroni, G. Genomic analysis of the TRIM family reveals two groups of genes with distinct evolutionary properties. BMC Evol. Biol. 2008, 8, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallery, D.L.; McEwan, W.A.; Bidgood, S.R.; Towers, G.J.; Johnson, C.M.; James, L.C. Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21). Proc. Natl. Acad. Sci. USA 2010, 107, 19985–19990. [Google Scholar] [CrossRef] [Green Version]
- Bratsch, S.; Wertz, N.; Chaloner, K.; Kunz, T.H.; Butler, J.E. The little brown bat, M. lucifugus, displays a highly diverse V-H, D-H and J(H) repertoire but little evidence of somatic hypermutation. Dev. Comp. Immunol. 2011, 35, 421–430. [Google Scholar] [CrossRef]
- Jebb, D.; Huang, Z.; Pippel, M.; Hughes, G.M.; Lavrichenko, K.; Devanna, P.; Winkler, S.; Jermiin, L.S.; Skirmuntt, E.C.; Katzourakis, A.; et al. Six reference-quality genomes reveal evolution of bat adaptations. Nature 2020, 583, 578. [Google Scholar] [CrossRef]
- Perera, S.; Mankoo, B.; Gautel, M. Developmental regulation of MURF E3 ubiquitin ligases in skeletal muscle. J. Muscle Res. Cell Motil. 2012, 33, 107–122. [Google Scholar] [CrossRef] [Green Version]
- Vuong, J.K.; Ergin, V.; Chen, L.; Zheng, S. Multilayered regulations of alternative splicing, NMD, and protein stability control temporal induction and tissue-specific expression of TRIM46 during axon formation. Nat. Commun. 2022, 13, 1–18. [Google Scholar] [CrossRef]
- Menon, S.; Boyer, N.P.; Winkle, C.C.; McClain, L.M.; Hanlin, C.C.; Pandey, D.; Rothenfusser, S.; Taylor, A.M.; Gupton, S.L. The E3 Ubiquitin Ligase TRIM9 Is a Filopodia Off Switch Required for Netrin-Dependent Axon Guidance. Dev. Cell 2015, 35, 698–712. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xu, H.; Liu, J.; Xie, N.; Gao, L.; He, Y.; Yao, Y.; Lv, F.; Zhang, Y.; Lu, J.; et al. Functional and Adaptive Significance of Promoter Mutations That Affect Divergent Myocardial Expressions of TRIM72 in Primates. Mol. Biol. Evol. 2021, 38, 2930–2945. [Google Scholar] [CrossRef] [PubMed]
- Pasquesi, G.I.M.; Kelly, C.J.; Ordonez, A.D.; Chuong, E.B. Transcriptional dynamics of transposable elements in the type I IFN response in Myotis lucifugus cells. Mob. DNA 2022, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Hölzer, M.; Schoen, A.; Wulle, J.; Müller, M.A.; Drosten, C.; Marz, M.; Weber, F. Virus- and Interferon Alpha-Induced Transcriptomes of Cells from the Microbat Myotis daubentonii. iScience 2019, 19, 647–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Lou, D.I.; Sawyer, S.L. Identification of a Genomic Reservoir for New TRIM Genes in Primate Genomes. PLoS Genet. 2011, 7, e1002388. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.Y.; Liang, L.; Zhu, Z.H.; Zhou, W.P.; Irwin, D.M.; Zhang, Y.P. Adaptive evolution of energy metabolism genes and the origin of flight in bats. Proc. Natl. Acad. Sci. USA 2010, 107, 8666–8671. [Google Scholar] [CrossRef] [Green Version]
- Carthagena, L.; Bergamaschi, A.; Luna, J.M.; David, A.; Uchil, P.D.; Margottin-Goguet, F.; Mothes, W.; Hazan, U.; Transy, C.; Pancino, G.; et al. Human TRIM Gene Expression in Response to Interferons. PLoS ONE 2009, 4, e4894. [Google Scholar] [CrossRef] [Green Version]
- Pagani, I.; Poli, G.; Vicenzi, E. TRIM22. A Multitasking Antiviral Factor. Cells 2021, 10, 1864. [Google Scholar] [CrossRef]
- Gu, Z.; Chen, X.; Yang, W.; Qi, Y.; Yu, H.; Wang, X.; Gong, Y.; Chen, Q.; Zhong, B.; Dai, L.; et al. The SUMOylation of TAB2 mediated by TRIM60 inhibits MAPK/NF-kappa B activation and the innate immune response. Cell. Mol. Immunol. 2021, 18, 1981–1994. [Google Scholar] [CrossRef]
- Huang, C.-J.; Huang, C.-C.; Chang, C.-C. Association of the testis-specific TRIM/RBCC protein RNF33/TRIM60 with the cytoplasmic motor proteins KIF3A and KIF3B. Mol. Cell. Biochem. 2012, 360, 121–131. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, B.; Racey, P.A.; Wang, Y.-L.; Zhang, S.-Y. Sperm storage, delayed ovulation, and menstruation of the female rickett’s big-footed bat (Myotis ricketti). Zool. Stud. 2008, 47, 215–221. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Suborder | Family | Species | Assembly Name | Coverage (×) | Scaffold N50 (kb) |
|---|---|---|---|---|---|
| Yinptero- chiroptera | Pteropodidae | Pteropus vampyrus | Pvam_2.0 | 188 | 5954 |
| Pteropus alecto | ASM32557v1 | 110 | 15,955 | ||
| Rousettus aegyptiacus | mRouAeg1.p | 64.6 | 113,812 | ||
| Hipposideridae | Hipposideros armiger | ASM189008v1 | 218.6 | 2328 | |
| Yango- chiroptera | Molossidae | Molossus molossus | mMolMol1.p | 58.3 | 110,665 |
| Vespertilionidae | Myotis davidii | ASM32734v1 | 110 | 3454 | |
| Myotis myotis | mMyoMyo1.p | 90.9 | 94,449 | ||
| Myotis brandtii | ASM41265v1 | 120 | 3226 | ||
| Myotis lucifugus | Myoluc2.0 | 7 | 4293 | ||
| Eptesicus fuscus | EptFus1.0 | 84 | 13,455 | ||
| Pipistrellus kuhlii | mPipKuh1.p | 80.8 | 80,237 | ||
| Miniopterus natalensis | Mnat.v1 | 77.0 | 4315 | ||
| Phyllostomidae | Phyllostomus discolor | mPhyDis1.pri.v3 | 65.8 | 171,743 | |
| Sturnira hondurensis | WHU_Shon_v2 | 158.0 | 10,165 | ||
| Artibeus jamaicensis | WHU_Ajam_v2 | 202.0 | 2589 | ||
| Desmodus rotundus | ASM294091v2 | 94 | 26,870 |
| TRIM Gene | lnL (m7) | lnL (M8) | 2ΔlnL | p-Value | Positive Selection Sites Identified by Both PAML and HYPHY |
|---|---|---|---|---|---|
| TRIM5 | −32,644.82 | −32,434.64 | 420.38 | <0.001 | 7, 71, 93, 145, 162, 165, 179, 181, 188, 209, 230, 270, 283, 311, 324, 325, 326, 374, 378, 381, 383, 389, 401, 424, 433, 484, 490 |
| TRIM13 | −5282.01 | −5250.82 | 62.37 | <0.001 | 312, 346 |
| TRIM20 | −14,407.60 | −14,385.42 | 44.37 | <0.001 | 55, 183, 195, 225, 229, 255, 273, 371, 395, 671, 676, 677, 689 |
| TRIM21 | −5451.73 | −5435.53 | 32.40 | <0.001 | 46, 50, 60, 405 |
| TRIM22 | −21,774.11 | −21,705.86 | 136.50 | <0.001 | 4, 19, 45, 53, 89, 120, 146, 248, 290, 293, 302, 323, 353, 390, 404, 407, 409, 427, 453 |
| TRIM23 | −4970.69 | −4964.70 | 11.97 | 0.003 | 561, 566 |
| TRIM24 | −11,079.17 | −11,022.77 | 112.81 | <0.001 | 3, 4, 6, 7, 21, 25 |
| TRIM25 | −10,989.77 | −10,960.90 | 57.75 | <0.001 | 93, 313, 337, 350, 412, 423, 429 |
| TRIM29 | −12,198.15 | −12,190.60 | 15.09 | 0.001 | 770 |
| TRIM33 | −10,163.69 | −10,159.81 | 7.76 | 0.021 | 5 |
| TRIM34 | −8042.75 | −8035.51 | 14.48 | 0.001 | 9, 107, 163, 505 |
| TRIM37 | −10,419.03 | −10,181.11 | 475.82 | <0.001 | 972, 980 |
| TRIM38 | −19,320.11 | −19,312.62 | 14.98 | 0.001 | 47, 181, 229 |
| TRIM43 | −8556.49 | −8538.28 | 36.42 | <0.001 | 50, 169, 283, 314, 332, 429, 432, 442 |
| TRIM44 | −7106.06 | −7072.26 | 67.60 | <0.001 | 6, 271, 272, 274 |
| TRIM45 | −8511.14 | −8505.68 | 10.91 | 0.004 | 551, 580 |
| TRIM54 | −4339.02 | −4326.17 | 25.71 | <0.001 | 270 |
| TRIM55 | −8167.15 | −8128.54 | 77.23 | <0.001 | 169 |
| TRIM56 | −9273.58 | −9266.46 | 14.24 | 0.001 | 1, 6, 10 |
| TRIM60 | −37,265.91 | −37,192.27 | 147.27 | <0.001 | 37, 100, 168, 234, 275, 308, 311, 320, 324, 354, 358, 375, 385, 386, 387, 394, 402, 458 |
| TRIM65 | −6766.77 | −6763.43 | 6.68 | 0.036 | 34, 36, 364 |
| TRIM69 | −5868.04 | −5856.62 | 22.85 | <0.001 | 123, 202, 375 |
| TRIM75 | −29,293.72 | −29,277.40 | 32.65 | <0.001 | 134, 167, 462 |
| TRIML2 | −10,138.56 | −10,125.39 | 26.33 | <0.001 | 11, 14, 16, 26, 54, 101, 191, 335, 339, 438 |
| Species | Cell Type | Accession Number | Treatment |
|---|---|---|---|
| Myotis lucifugus | embryonic fibroblast cell | SRR18761563–SRR18761565 | 4 h_IFNa |
| SRR18761566–SRR18761568 | 4 h_control | ||
| Myotis daubentonii | kidney cell line | SRR8062281–SRR8062283 | 6 h_control |
| SRR8062284–SRR8062286 | 24 h_control | ||
| SRR8062287–SRR8062289 | 6 h_IFNa | ||
| SRR8062293–SRR8062296 | 6 h_RVFV | ||
| SRR8062297–SRR8062299 | 24 h_RVFV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, J.; Tan, B.; Zhang, Y. Positive Selection and Duplication of Bat TRIM Family Proteins. Viruses 2023, 15, 875. https://doi.org/10.3390/v15040875
Xie J, Tan B, Zhang Y. Positive Selection and Duplication of Bat TRIM Family Proteins. Viruses. 2023; 15(4):875. https://doi.org/10.3390/v15040875
Chicago/Turabian StyleXie, Jiazheng, Bowen Tan, and Yi Zhang. 2023. "Positive Selection and Duplication of Bat TRIM Family Proteins" Viruses 15, no. 4: 875. https://doi.org/10.3390/v15040875