
Development and Evaluation of an Enzyme-Linked Immunosorbent Assay Targeting Rabies-Specific IgM and IgG in Human Sera

Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. RFFIT Serological Testing
2.3. Indirect ELISA Serological Testing
2.4. Development of Anti-Rabies IgM and IgG ELISA for Human Samples
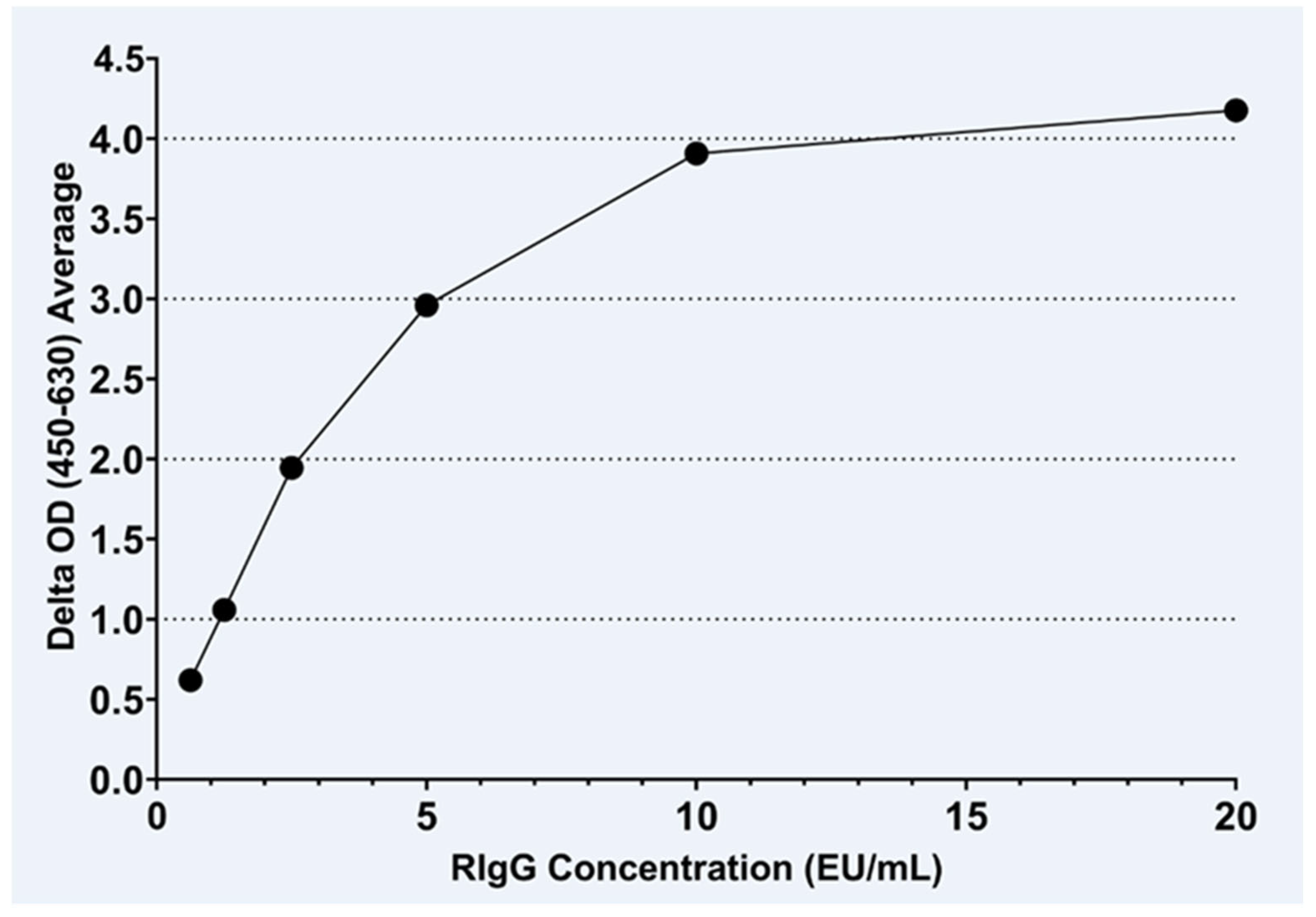
2.4.1. Rabies Anti-IgG ELISA Development
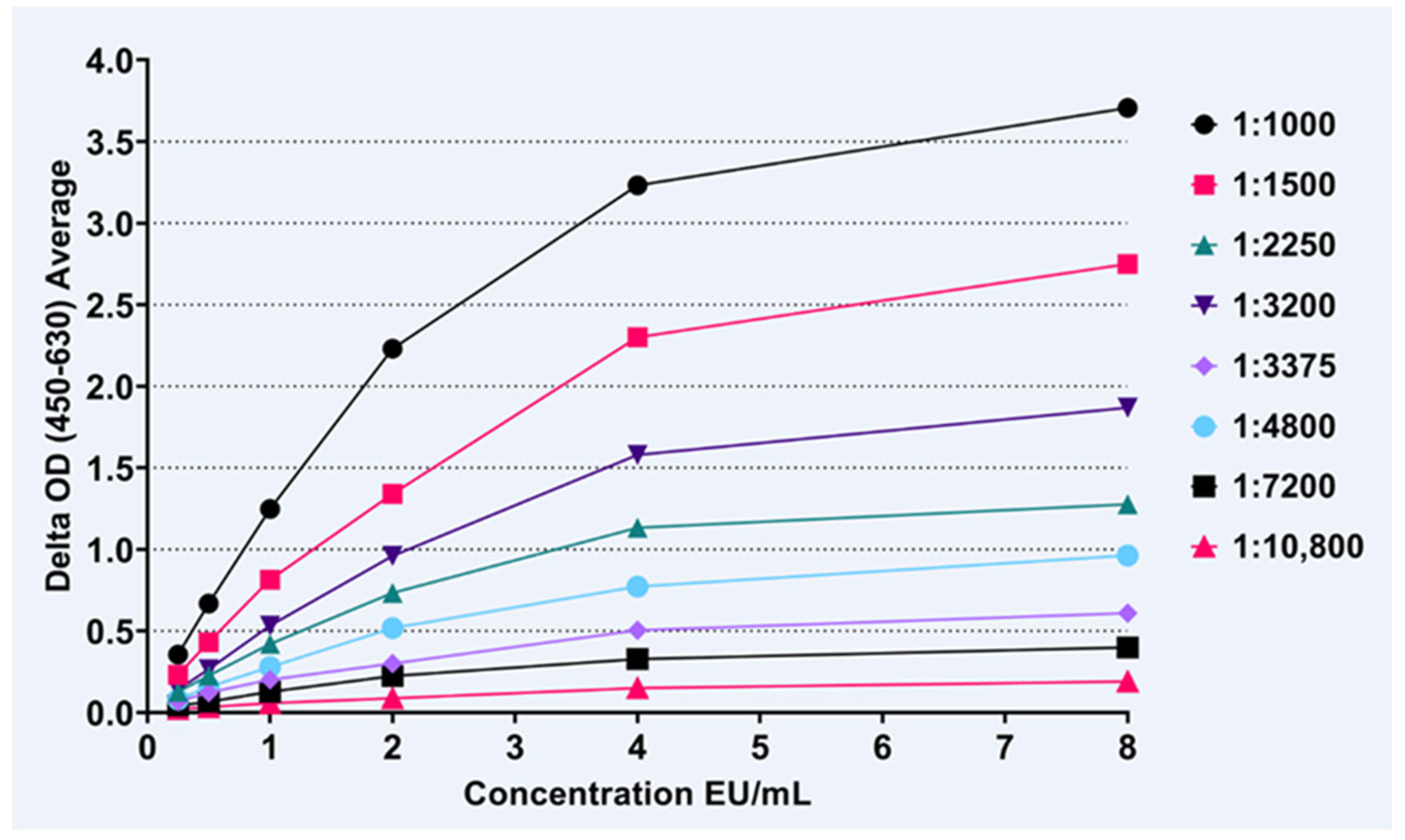
2.4.2. Rabies Anti-IgM ELISA Development
2.5. Statistical Methods
3. Results
3.1. IgG Assay
3.2. IgM Assay
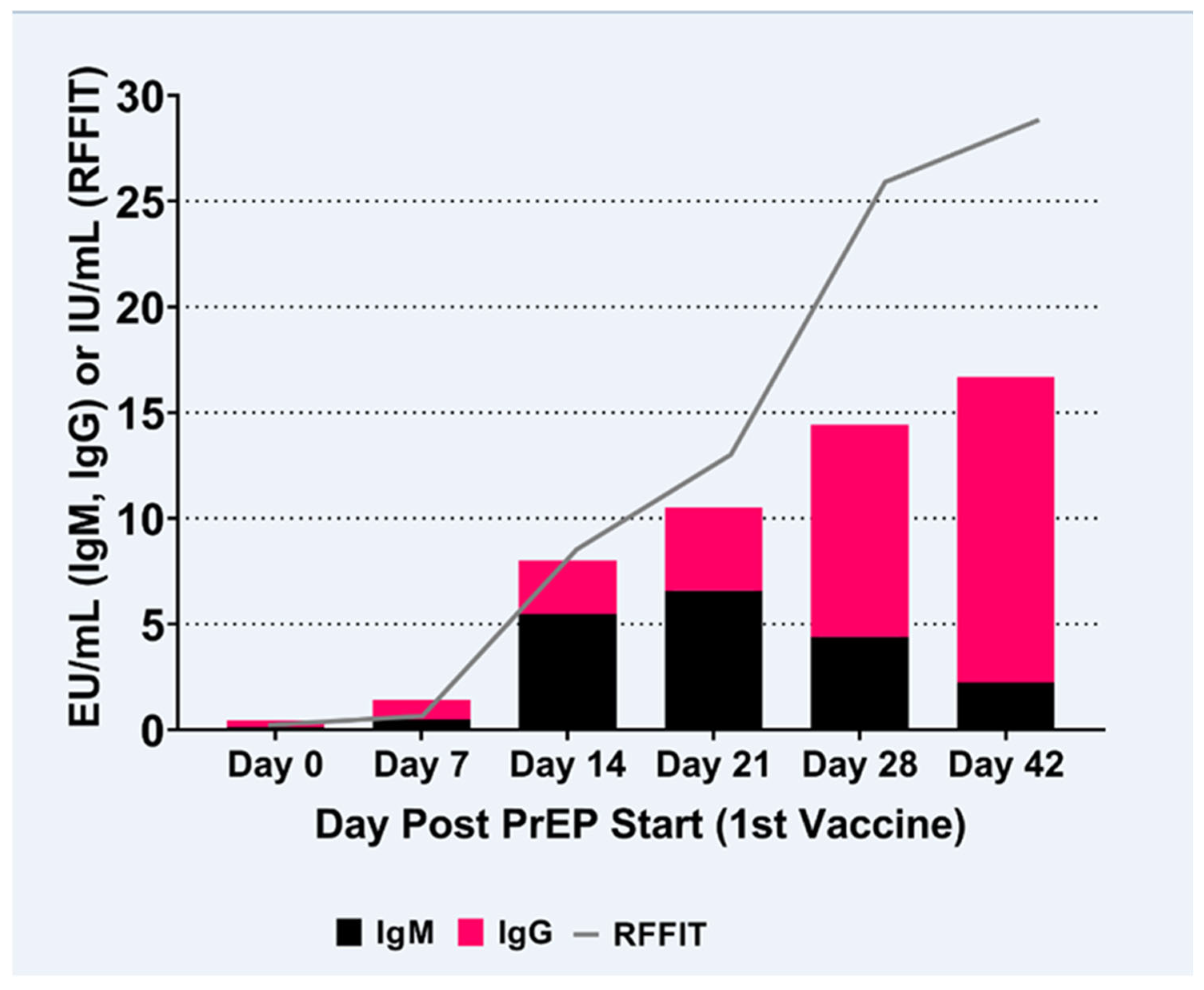
3.3. IgM to IgG ELISA Ratio

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DPV | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| 7 | 522.4 | 39.9 | 221.4 | 373.2 | 39.9 | 113.4 | 39.9 | |||
| 10 | 119.8 | |||||||||
| 14–16 | 421.1 | 537.3 | 191.7 | 369.4 | 449.8 | 337.3 | 39.9 | 25.2 | ||
| 21 | 383.3 | 278.4 | 107.6 | 226.3 | 71.7 | 147.5 | 96.8 | |||
| 28–31 | 35 | 84.8 | 20.9 | 21.4 | 34.2 | 364.5 | 26.8 | 134.1 | 49.1 | |
| 42 | 16.1 | 25.5 | 6.7 | 16.4 | 15.6 | 10.9 | ||||
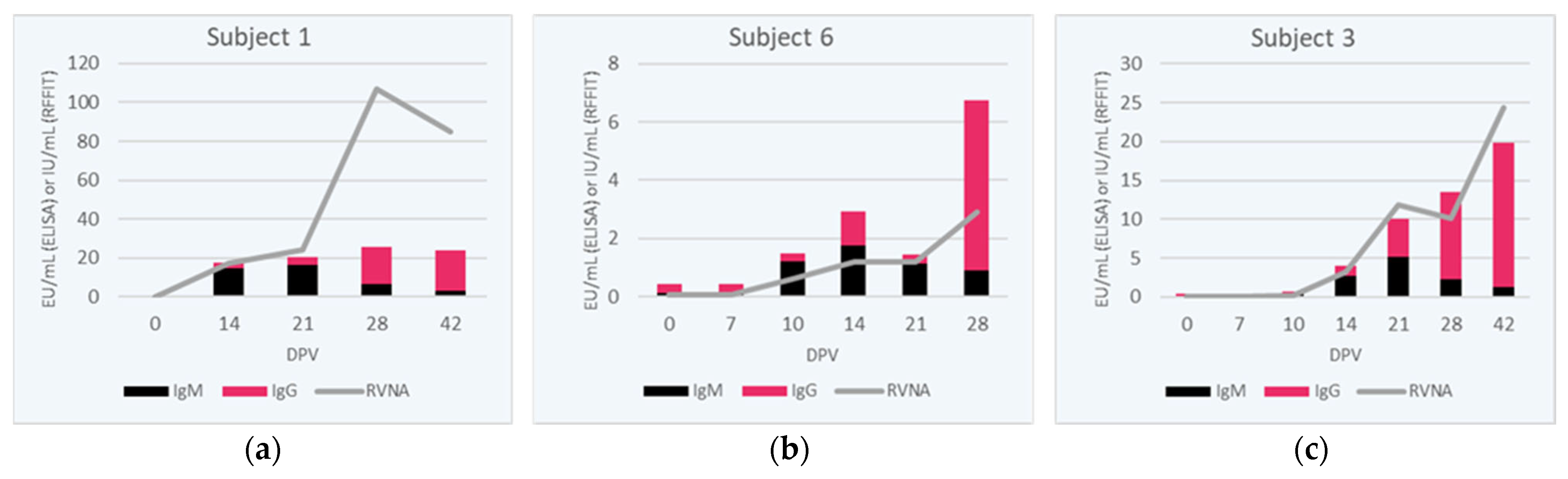
3.4. RVNA Isotype Switching Kinetics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rupprecht, C.E.; Abela-Ridder, B.; Abila, R.; Amparo, A.C.; Banyard, A.; Blanton, J.; Chanachai, K.; Dallmeier, K.; de Balogh, K.; Del Rio Vilas, V.; et al. Towards rabies elimination in the Asia-Pacific region: From theory to practice. Biologicals 2020, 64, 83–95. [Google Scholar] [CrossRef]
- Fahrion, A.S.; Taylor, L.H.; Torres, G.; Muller, T.; Durr, S.; Knopf, L.; de Balogh, K.; Nel, L.H.; Gordoncillo, M.J.; Abela-Ridder, B. The Road to Dog Rabies Control and Elimination-What Keeps Us from Moving Faster? Front. Public Health 2017, 5, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO Expert Consultation on Rabies, Third Report; World Health Organization: Geneva, Switzerland, 2018; pp. 1–195. [Google Scholar]
- Rao, A.K.; Briggs, D.; Moore, S.M.; Whitehill, F.; Campos-Outcalt, D.; Morgan, R.L.; Wallace, R.M.; Romero, J.R.; Bahta, L.; Frey, S.E.; et al. Use of a Modified Preexposure Prophylaxis Vaccination Schedule to Prevent Human Rabies: Recommendations of the Advisory Committee on Immunization Practices—United States, 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 619–627. [Google Scholar] [CrossRef] [PubMed]
- WHO. Rabies Vaccines: WHO Position Paper—April 2018; Abela Ridder, B., Ed.; World Health Organization: Geneva, Switzerland, 2018; pp. 201–220. [Google Scholar]
- Kessels, J.A.; Recuenco, S.; Navarro-Vela, A.M.; Deray, R.; Vigilato, M.; Ertl, H.; Durrheim, D.; Rees, H.; Nel, L.H.; Abela-Ridder, B.; et al. Pre-exposure rabies prophylaxis: A systematic review. Bull. World Health Organ. 2017, 95, 210C–219C. [Google Scholar] [CrossRef] [PubMed]
- Kessels, J.; Tarantola, A.; Salahuddin, N.; Blumberg, L.; Knopf, L. Rabies post-exposure prophylaxis: A systematic review on abridged vaccination schedules and the effect of changing administration routes during a single course. Vaccine 2019, 37 (Suppl. 1), A107–A117. [Google Scholar] [CrossRef]
- Soentjens, P.; Andries, P.; Aerssens, A.; Tsoumanis, A.; Ravinetto, R.; Heuninckx, W.; van Loen, H.; Brochier, B.; Van Gucht, S.; Van Damme, P.; et al. Preexposure Intradermal Rabies Vaccination: A Noninferiority Trial in Healthy Adults on Shortening the Vaccination Schedule From 28 to 7 Days. Clin. Infect. Dis. 2019, 68, 607–614. [Google Scholar] [CrossRef]
- Warrell, M.J.; Riddell, A.; Yu, L.M.; Phipps, J.; Diggle, L.; Bourhy, H.; Deeks, J.J.; Fooks, A.R.; Audry, L.; Brookes, S.M.; et al. A simplified 4-site economical intradermal post-exposure rabies vaccine regimen: A randomised controlled comparison with standard methods. PLoS Negl. Trop. Dis. 2008, 2, e224. [Google Scholar] [CrossRef] [Green Version]
- Briggs, D.J.; Moore, S.M. The Route of Administration of Rabies Vaccines: Comparing the Data. Viruses 2021, 13, 1252. [Google Scholar] [CrossRef]
- Johnson, N.; Cunningham, A.F.; Fooks, A.R. The immune response to rabies virus infection and vaccination. Vaccine 2010, 28, 3896–3901. [Google Scholar] [CrossRef]
- Turner, G.S. Immunoglobulin (IgG) and (IgM) antibody responses to rabies vaccine. J. Gen. Virol. 1978, 40, 595–604. [Google Scholar] [CrossRef]
- Jackson, A.C. Therapy of human rabies. Adv. Virus Res. 2011, 79, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Dorfmeier, C.L.; Lytle, A.G.; Dunkel, A.L.; Gatt, A.; McGettigan, J.P. Protective vaccine-induced CD4(+) T cell-independent B cell responses against rabies infection. J. Virol. 2012, 86, 11533–11540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGettigan, J.P. Experimental rabies vaccines for humans. Expert Rev. Vaccines 2010, 9, 1177–1186. [Google Scholar] [CrossRef]
- Dorfmeier, C.L.; Shen, S.; Tzvetkov, E.P.; McGettigan, J.P. Reinvestigating the role of IgM in rabies virus postexposure vaccination. J. Virol. 2013, 87, 9217–9222. [Google Scholar] [CrossRef] [Green Version]
- Flamand, A.; Raux, H.; Gaudin, Y.; Ruigrok, R.W. Mechanisms of rabies virus neutralization. Virology 1993, 194, 302–313. [Google Scholar] [CrossRef]
- Moore, S.M.; Hanlon, C.A. Rabies-specific antibodies: Measuring surrogates of protection against a fatal disease. PLoS Negl. Trop. Dis. 2010, 4, e595. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.M.; Gordon, C.R.; Hanlon, C.A. Measures of Rabies Immunity. In Rabies; Jackson, A.C., Ed.; Elsevier: San Diego, CA, USA, 2013; pp. 461–496. [Google Scholar]
- Ertl, H.C.J. Human immune responses to traditional and novel rabies vaccines. Rev. Sci. Tech. 2018, 37, 649–656. [Google Scholar] [CrossRef]
- CDC. Human Rabies Prevention—United States, 2008 Recommendatoins of the Advisory Committee On Immunization Practices. In MMWR; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2008; pp. 1–28. [Google Scholar]
- Yager, M.L.; Moore, S. The Rapid Fluorescent Focus Inhibition Test. In Rabies Diagnosis, Research, and Prevention, Volume Two; Rupprecht, C.E., Nagarajan, T., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 199–215. [Google Scholar]
- Feyssaguet, M.; Dacheux, L.; Audry, L.; Compoint, A.; Morize, J.L.; Blanchard, I.; Bourhy, H. Multicenter comparative study of a new ELISA, PLATELIA RABIES II, for the detection and titration of anti-rabies glycoprotein antibodies and comparison with the rapid fluorescent focus inhibition test (RFFIT) on human samples from vaccinated and non-vaccinated people. Vaccine 2007, 25, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M. Development and Evaluation of a Rabies Enzyme-Linked Immunosorbent Assay (ELISA) Targeting IgM and IgG in Human Sera. Master’s Thesis, Department of Diagnostic Medicine/Pathobiology, Kansas State University, Manhattan, KS, USA, 2019. [Google Scholar]
- Dietzschold, B. Antibody-mediated clearance of viruses from the mammalian central nervous system. Trends Microbiol. 1993, 1, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.C.; Morimoto, K.; Bette, M.; Weihe, E.; Koprowski, H.; Dietzschold, B. Collaboration of antibody and inflammation in clearance of rabies virus from the central nervous system. J. Virol. 1998, 72, 3711–3719. [Google Scholar] [CrossRef] [Green Version]
- The Immunological Basis for Immunization Series: Module 17: Rabies Vaccines; World Health Organization: Geneva, Switzerland, 2017.
- Irie, T.; Kawai, A. Further studies on the mechanism of rabies virus neutralization by a viral glycoprotein-specific monoclonal antibody, #1-46-12. Microbiol. Immunol. 2005, 49, 721–731. [Google Scholar]
- Dorfmeier, C.L.; Tzvetkov, E.P.; Gatt, A.; McGettigan, J.P. Investigating the role for IL-21 in rabies virus vaccine-induced immunity. PLoS Negl. Trop. Dis. 2013, 7, e2129. [Google Scholar] [CrossRef] [Green Version]
- Riepenhoff-Talty, M.; Bogger-Goren, S.; Li, P.; Carmody, P.J.; Barrett, H.J.; Ogra, P.L. Development of serum and intestinal antibody response to rotavirus after naturally acquired rotavirus infection in man. J. Med. Virol. 1981, 8, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Pas, S.D.; Streefkerk, R.H.; Pronk, M.; de Man, R.A.; Beersma, M.F.; Osterhaus, A.D.; van der Eijk, A.A. Diagnostic performance of selected commercial HEV IgM and IgG ELISAs for immunocompromised and immunocompetent patients. J. Clin. Virol. 2013, 58, 629–634. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, X.; Jiang, H.; Yan, Q.; Ai, X.; Wang, Y.; Cai, J.; Jiang, L.; Wu, T.; Wang, Z.; et al. Profile of acute infectious markers in sporadic hepatitis E. PLoS ONE 2010, 5, e13560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nystad, T.W.; Myrmel, H. Prevalence of primary versus reactivated Epstein-Barr virus infection in patients with VCA IgG-, VCA IgM- and EBNA-1-antibodies and suspected infectious mononucleosis. J. Clin. Virol. 2007, 38, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Jespersen, D.J.; Harring, J.A.; Binnicker, M.J. Evaluation of the Bio-Rad BioPlex 2200 syphilis multiplex flow immunoassay for the detection of IgM- and IgG-class antitreponemal antibodies. Clin. Vaccine Immunol. 2010, 17, 966–968. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.L.; Edjlalipour, M.; Luger, A. Comparative evaluation of nine different enzyme-linked immunosorbent assays for determination of antibodies against Treponema pallidum in patients with primary syphilis. J. Clin. Microbiol. 2000, 38, 1279–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.S.; Petersen, C.S.; Axelsen, N.H. Enzyme-linked immunosorbent assay for detection of immunoglobulin M antibody against the Reiter treponeme flagellum in syphilis. J. Clin. Microbiol. 1982, 16, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Servat, A.; Feyssaguet, M.; Blanchard, I.; Morize, J.L.; Schereffer, J.L.; Boue, F.; Cliquet, F. A quantitative indirect ELISA to monitor the effectiveness of rabies vaccination in domestic and wild carnivores. J. Immunol. Methods 2007, 318, 1–10. [Google Scholar] [CrossRef]
- Denis, M.; Knezevic, I.; Wilde, H.; Hemachudha, T.; Briggs, D.; Knopf, L. An overview of the immunogenicity and effectiveness of current human rabies vaccines administered by intradermal route. Vaccine 2019, 37 (Suppl. 1), A99–A106. [Google Scholar] [CrossRef]
- Giesen, A.; Gniel, D.; Malerczyk, C. 30 Years of rabies vaccination with Rabipur: A summary of clinical data and global experience. Expert Rev. Vaccines 2015, 14, 351–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwert, E.; Barsenbach, J.; Werner, J.; Mardus, I.; Scheiermann, N.; Grosse-Wilde, H. Early/high and late/low responders among HDCS vaccinees. In Cell Culture Rabies Vaccines and Their Protective Effect in Man; Wiktor, T.J., Koprowski, H., Eds.; International Green Cross: Geneva, Switzerland, 1981; pp. 160–168. [Google Scholar]
- Strady, C.; Jaussaud, R.; Beguinot, I.; Lienard, M.; Strady, A. Predictive factors for the neutralizing antibody response following pre-exposure rabies immunization: Validation of a new booster dose strategy. Vaccine 2000, 18, 2661–2667. [Google Scholar] [CrossRef] [PubMed]
- Aubert, M.F. Practical significance of rabies antibodies in cats and dogs. Rev. Sci. Tech. 1992, 11, 735–760. [Google Scholar] [CrossRef] [PubMed]
- Bunn, T.O.; Ridpath, H.D. The relationship between rabies antibody titers in dogs and cats and protection from challenge. Public Health 1984, 11, 43–45. [Google Scholar]
- Moore, S.M.; Gilbert, A.; Vos, A.; Freuling, C.M.; Ellis, C.; Kliemt, J.; Muller, T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Trop. Med. Infect. Dis. 2017, 2, 31. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajac, M.D.; Ortega, M.T.; Moore, S.M. Development and Evaluation of an Enzyme-Linked Immunosorbent Assay Targeting Rabies-Specific IgM and IgG in Human Sera. Viruses 2023, 15, 874. https://doi.org/10.3390/v15040874
Zajac MD, Ortega MT, Moore SM. Development and Evaluation of an Enzyme-Linked Immunosorbent Assay Targeting Rabies-Specific IgM and IgG in Human Sera. Viruses. 2023; 15(4):874. https://doi.org/10.3390/v15040874
Chicago/Turabian StyleZajac, Michelle D., Maria Teresa Ortega, and Susan M. Moore. 2023. "Development and Evaluation of an Enzyme-Linked Immunosorbent Assay Targeting Rabies-Specific IgM and IgG in Human Sera" Viruses 15, no. 4: 874. https://doi.org/10.3390/v15040874