
Development of Porcine Monoclonal Antibodies with In Vitro Neutralizing Activity against Classical Swine Fever Virus from C-Strain E2-Specific Single B Cells

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Lines and Virus Strains
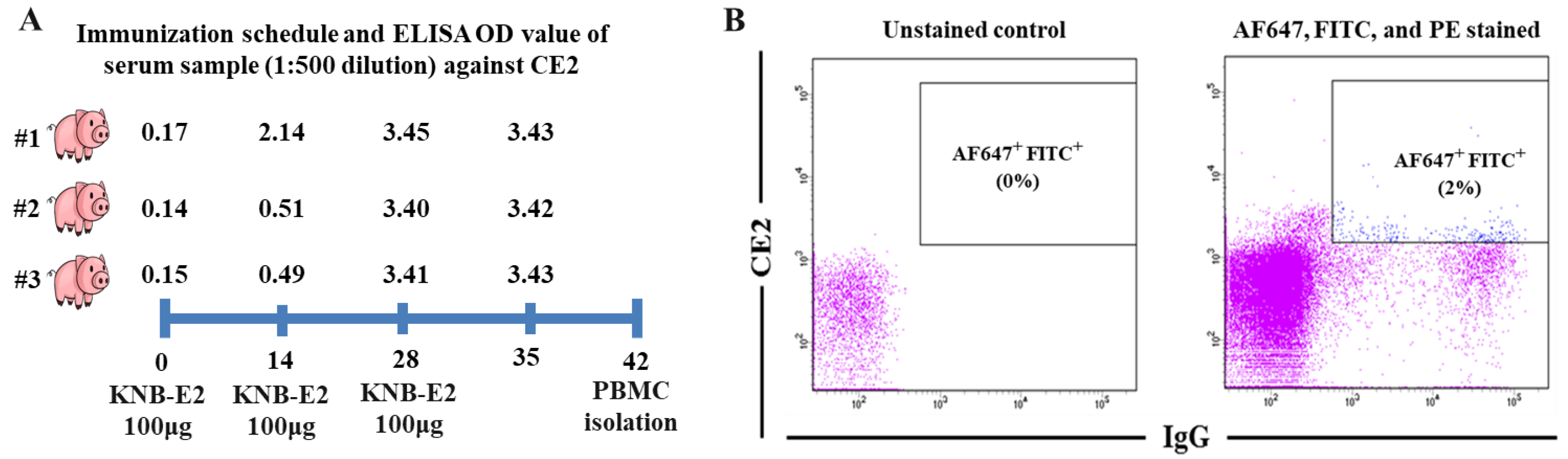
2.3. Animal Immunization and E2 Antibody Testing
2.4. Cell Staining and Porcine Single B Cell Sorting
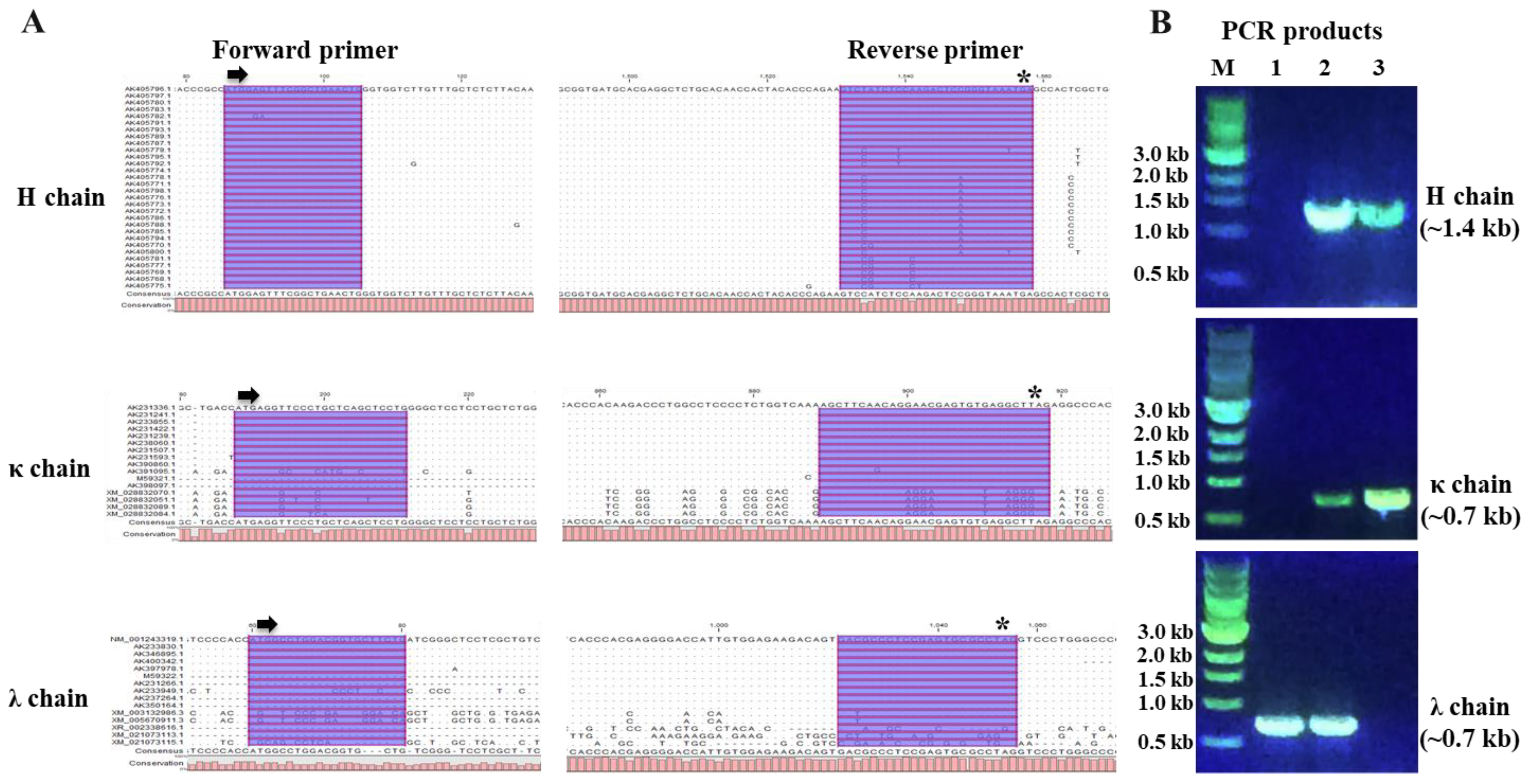
2.5. Porcine IgG Gene-Specific Single B Cell RT-PCR
2.6. Cloning and Expression of Porcine Monoclonal Antibodies in Mammalian Expression System
2.7. Western Blotting and Indirect Immunofluorescence (IFA) Testing of Porcine Monoclonal Antibodies
3. Results
3.1. Isolation of CSFV C-Strain E2 Protein-Induced Single Porcine B Cells
3.2. Amplification of IgG Heavy and Light Chains from Porcine Single B Cell
3.3. Genetic Characterizon of the Paired Chains Recovered from CE2+IgG+ Porcine Single B Cells
3.4. The Porcine mAbs Can Specifically Bind to CSFV CE2 Protein
3.5. The Porcine mAbs Exhibited High Potency in Neutralizing CSFVs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moennig, V. Introduction to classical swine fever: Virus, disease and control policy. Vet. Microbiol. 2000, 73, 93–102. [Google Scholar] [CrossRef]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical signs and epidemiology of classical swine fever: A review of new knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Madera, R.; Li, Y.; McVey, D.S.; Drolet, B.S.; Shi, J. Recent Advances in the Diagnosis of Classical Swine Fever and Future Perspectives. Pathogens 2020, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A Review of Classical Swine Fever Virus and Routes of Introduction into the United States and the Potential for Virus Establishment. Front. Vet. Sci. 2018, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Blome, S.; Meindl-Bohmer, A.; Loeffen, W.; Thuer, B.; Moennig, V. Assessment of classical swine fever diagnostics and vaccine performance. Rev. Sci. Tech. 2006, 25, 1025–1038. [Google Scholar] [CrossRef]
- Shi, J.; Wang, L.; McVey, D.S. Of pigs and men: The best-laid plans for prevention and control of swine fevers. Anim. Front. 2021, 11, 6–13. [Google Scholar] [CrossRef]
- Li, F.; Li, B.; Niu, X.; Chen, W.; Li, Y.; Wu, K.; Li, X.; Ding, H.; Zhao, M.; Chen, J.; et al. The Development of Classical Swine Fever Marker Vaccines in Recent Years. Vaccines 2022, 10, 603. [Google Scholar] [CrossRef]
- Madera, R.; Gong, W.; Wang, L.; Burakova, Y.; Lleellish, K.; Galliher-Beckley, A.; Nietfeld, J.; Henningson, J.; Jia, K.; Li, P.; et al. Pigs immunized with a novel E2 subunit vaccine are protected from subgenotype heterologous classical swine fever virus challenge. BMC Vet. Res. 2016, 12, 197. [Google Scholar] [CrossRef] [Green Version]
- Madera, R.; Wang, L.; Gong, W.; Burakova, Y.; Buist, S.; Nietfeld, J.; Henningson, J.; Ozuna, A.G.C.; Tu, C.; Shi, J. Towards the development of a one-dose classical swine fever subunit vaccine: Antigen titration, onset and duration of immunity. J. Vet. Sci. 2018, 19, 393–405. [Google Scholar] [CrossRef]
- Moormann, R.J.; Bouma, A.; Kramps, J.A.; Terpstra, C.; De Smit, H.J. Development of a classical swine fever subunit marker vaccine and companion diagnostic test. Vet. Microbiol. 2000, 73, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Coronado, L.; Perera, C.L.; Rios, L.; Frías, M.T.; Pérez, L.J. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, R.C.; Madera, R.; Peres, Y.; Berquist, B.R.; Wang, L.; Buist, S.; Burakova, Y.; Palle, S.; Chung, C.J.; Rasmussen, M.V.; et al. Plant-made E2 glycoprotein single-dose vaccine protects pigs against classical swine fever. Plant Biotechnol. J. 2019, 7, 410–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eblé, P.L.; Geurts, Y.; Quak, S.; Moonen-Leusen, H.W.; Blome, S.; Hofmann, M.A.; Koenen, F.; Beer, M.; Loeffen, W.L.A. Efficacy of chimeric Pestivirus vaccine candidates against classical swine fever: Protection and DIVA characteristics. Vet. Microbiol. 2013, 162, 437–446. [Google Scholar] [CrossRef]
- Gabriel, C.; Blome, S.; Urniza, A.; Juanola, S.; Koenen, F.; Beer, M. Towards licensing of CP7_E2alf as marker vaccine against classical swine fever-Duration of immunity. Vaccine 2012, 30, 2928–2936. [Google Scholar] [CrossRef]
- Pannhorst, K.; Fröhlich, A.; Staubach, C.; Meyer, D.; Blome, S.; Becher, P. Evaluation of an Erns-based enzyme-linked immunosorbent assay to distinguish Classical swine fever virus-infected pigs from pigs vaccinated with CP7_E2alf. J. Vet. Diagn. Invest. 2015, 27, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Salazar, G.; Zhang, N.; Fu, T.M.; An, Z. Antibody therapies for the prevention and treatment of viral infections. NPJ Vaccines 2017, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Burton, D.R. Passive immunotherapy of viral infections: ‘Super-antibodies’ enter the fray. Nat. Rev. Immunol. 2018, 18, 297–308. [Google Scholar] [CrossRef]
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef]
- Hammarström, L.; Marcotte, H.; Piralla, A.; Baldanti, F.; Pan-Hammarström, Q. Antibody therapy for COVID-19. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 553–558. [Google Scholar] [CrossRef]
- Cruz-Teran, C.; Tiruthani, K.; McSweeney, M.; Ma, A.; Pickles, R.; Lai, S.K. Challenges and opportunities for antiviral monoclonal antibodies as COVID-19 therapy. Adv. Drug. Deliv. Rev. 2021, 169, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Galliher-Beckley, A.; Pappan, L.K.; Madera, R.; Burakova, Y.; Waters, A.; Nickles, M.; Li, X.; Nietfeld, J.; Schlup, J.R.; Zhong, Q.; et al. Characterization of a novel oil-in-water emulsion adjuvant for swine influenza virus and Mycoplasma hyopneumoniae vaccines. Vaccine 2015, 33, 2903–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Li, Z.; Li, J.; Huang, T.; Peng, G.; Tang, W.; Yi, G.; Zhang, L.; Song, Y.; Liu, T.; et al. Revisiting the Pig IGHC Gene Locus in Different Breeds Uncovers Nine Distinct IGHG Genes. J. Immunol. 2020, 205, 2137–2145. [Google Scholar] [CrossRef]
- Wang, L.; Mi, S.; Madera, R.; Ganges, L.; Borca, M.V.; Ren, J.; Cunningham, C.; Cino-Ozuna, A.G.; Li, H.; Tu, C.; et al. A neutralizing monoclonal antibody-based competitive ELISA for classical swine fever C-strain post-vaccination monitoring. BMC Vet. Res. 2020, 16, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legouffe, E.; Liautard, J.; Gaillard, J.P.; Rossi, J.F.; Wijdenes, J.; Bataille, R.; Klein, B.; Brochier, J. Human anti-mouse antibody response to the injection of murine monoclonal antibodies against IL-6. Clin. Exp. Immunol. 1994, 98, 323–329. [Google Scholar] [CrossRef]
- Zheng, D. Antibody gene therapy: An attractive approach for the treatment of cancers and other chronic diseases. Cell Res. 2007, 17, 303–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, J.M.; Rosensweig, C.J.; Faden, L.B.; Dewitz, M.C. Monoclonal antibody successes in the clinic. Nat. Biotechnol. 2005, 23, 1073–1078. [Google Scholar] [CrossRef]
- Pedrioli, A.; Oxenius, A. Single B cell technologies for monoclonal antibody discovery. Trends Immunol. 2021, 42, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Tiller, T. Single B cell antibody technologies. New Biotechnol. 2011, 28, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, N.; Yoshioka, M.; Fujimoto, R.; Yamagishi, F.; Isobe, M. Rapid production of antigen-specific monoclonal antibodies from a variety of animals. BMC Biol. 2012, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Starkie, D.O.; Compson, J.E.; Rapecki, S.; Lightwood, D.J. Generation of Recombinant Monoclonal Antibodies from Immunised Mice and Rabbits via Flow Cytometry and Sorting of Antigen-Specific IgG+ Memory B Cells. PLoS ONE 2016, 11, e0152282. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Morishita, S.; Uchida, Y.; Nagai, S.; Kojima, T.; Nakano, H. Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology. Antibodies 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Tran, K.; Wang, Y.; Steinhardt, J.J.; Xiao, Y.; Chiang, C.I.; Wyatt, R.T.; Li, Y. Antigen-Specific Single B Cell Sorting and Monoclonal Antibody Cloning in Guinea Pigs. Front. Microbiol. 2019, 10, 672. [Google Scholar] [CrossRef] [Green Version]
- Booth, B.J.; Ramakrishnan, B.; Narayan, K.; Wollacott, A.M.; Babcock, G.J.; Shriver, Z.; Viswanathan, K. Extending human IgG half-life using structure-guided design. mAbs 2018, 10, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Clements, T.; Rice, T.F.; Vamvakas, G.; Barnett, S.; Barnes, M.; Donaldson, B.; Jones, C.E.; Kampmann, B.; Holder, B. Update on Transplacental Transfer of IgG Subclasses: Impact of Maternal and Fetal Factors. Front. Immunol. 2020, 11, 1920. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | PCR Product |
|---|---|---|
| D-WHF1 | CGGAATTCGCCATGGAGTTTCGGCTGAACTG | whole coding region of porcine IgH chain |
| D-WHF2 | CGGAATTCGCCATGGGATTTCGGCTGAACTG | |
| D-WHR1 | GCTCTAGATCATTTACCCGGAGTCTTGGAGATAGAC | |
| D-WHR2 | GCTCTAGATCATTTACCCKGAGTCTKGRAGAYGGAC | |
| D-WκF1 | CGGAATTCACCATGAGGTTCCCTGCTCAGCTCCTG | whole coding region of porcine Igκ chain |
| D-WκF2 | CGGAATTCACCATGAGGGCCCCCATGCACCTCCTTG | |
| D-WκF3 | CGGAATTCACCATGAGGGTCCCCGCTCAGCTCCTG | |
| D-WκR1 | GCTCTAGACTAAGCCTCACACTCGTTCCTGYTGAAGCT | |
| D-WκR2 | GCTCTAGACTAACACTCTCCTCTGTTGAAGCTCTTGG | |
| D-WλF1 | CGGAATTCACCATGGCCTGGAYCCCTCTCCT | whole coding region of porcine Igλ chain |
| D-WλF2 | CGGAATTCACCATGGCCTGGACGGTGCTTCTG | |
| D-WλF3 | CGGAATTCACCATGAGGCCCAGGTCAGGCCAG | |
| D-WλR1 | GCTCTAGACTAGGCGCACTCGGAGGGCRT |
| Subclasses | IgG3 | H9 | H11 | IgG4 | IgG2b | IgG2c | IgG2a | IgG1 | H1 | IgG5a | IgG5b | IgG5c |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IgG3 | ID | 0.80 | 0.80 | 0.81 | 0.83 | 0.83 | 0.82 | 0.82 | 0.79 | 0.75 | 0.75 | 0.75 |
| H9 | 0.80 | ID | 1.00 | 0.95 | 0.88 | 0.90 | 0.89 | 0.87 | 0.85 | 0.82 | 0.81 | 0.81 |
| H11 | 0.80 | 1.00 | ID | 0.95 | 0.88 | 0.89 | 0.89 | 0.87 | 0.85 | 0.82 | 0.80 | 0.81 |
| IgG4 | 0.81 | 0.95 | 0.95 | ID | 0.91 | 0.94 | 0.91 | 0.88 | 0.84 | 0.80 | 0.79 | 0.81 |
| IgG2b | 0.83 | 0.88 | 0.88 | 0.91 | ID | 0.97 | 0.96 | 0.92 | 0.86 | 0.84 | 0.80 | 0.81 |
| IgG2c | 0.83 | 0.90 | 0.89 | 0.94 | 0.97 | ID | 0.94 | 0.90 | 0.86 | 0.82 | 0.78 | 0.81 |
| IgG2a | 0.82 | 0.89 | 0.89 | 0.91 | 0.96 | 0.94 | ID | 0.92 | 0.86 | 0.85 | 0.81 | 0.81 |
| IgG1 | 0.82 | 0.87 | 0.87 | 0.88 | 0.92 | 0.90 | 0.92 | ID | 0.92 | 0.87 | 0.83 | 0.84 |
| H1 | 0.79 | 0.85 | 0.85 | 0.84 | 0.86 | 0.86 | 0.86 | 0.92 | ID | 0.82 | 0.80 | 0.80 |
| IgG5a | 0.75 | 0.82 | 0.82 | 0.80 | 0.84 | 0.82 | 0.85 | 0.87 | 0.82 | ID | 0.90 | 0.88 |
| IgG5b | 0.75 | 0.81 | 0.80 | 0.79 | 0.80 | 0.78 | 0.81 | 0.83 | 0.80 | 0.90 | ID | 0.89 |
| IgG5C | 0.75 | 0.81 | 0.81 | 0.81 | 0.81 | 0.81 | 0.81 | 0.84 | 0.80 | 0.88 | 0.89 | ID |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Madera, R.; Li, Y.; Gladue, D.P.; Borca, M.V.; McIntosh, M.T.; Shi, J. Development of Porcine Monoclonal Antibodies with In Vitro Neutralizing Activity against Classical Swine Fever Virus from C-Strain E2-Specific Single B Cells. Viruses 2023, 15, 863. https://doi.org/10.3390/v15040863
Wang L, Madera R, Li Y, Gladue DP, Borca MV, McIntosh MT, Shi J. Development of Porcine Monoclonal Antibodies with In Vitro Neutralizing Activity against Classical Swine Fever Virus from C-Strain E2-Specific Single B Cells. Viruses. 2023; 15(4):863. https://doi.org/10.3390/v15040863
Chicago/Turabian StyleWang, Lihua, Rachel Madera, Yuzhen Li, Douglas P. Gladue, Manuel V. Borca, Michael T. McIntosh, and Jishu Shi. 2023. "Development of Porcine Monoclonal Antibodies with In Vitro Neutralizing Activity against Classical Swine Fever Virus from C-Strain E2-Specific Single B Cells" Viruses 15, no. 4: 863. https://doi.org/10.3390/v15040863