
Leaf Curl Epidemic Risk in Chilli as a Consequence of Vector Migration Rate and Contact Rate Dynamics: A Critical Guide to Management

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
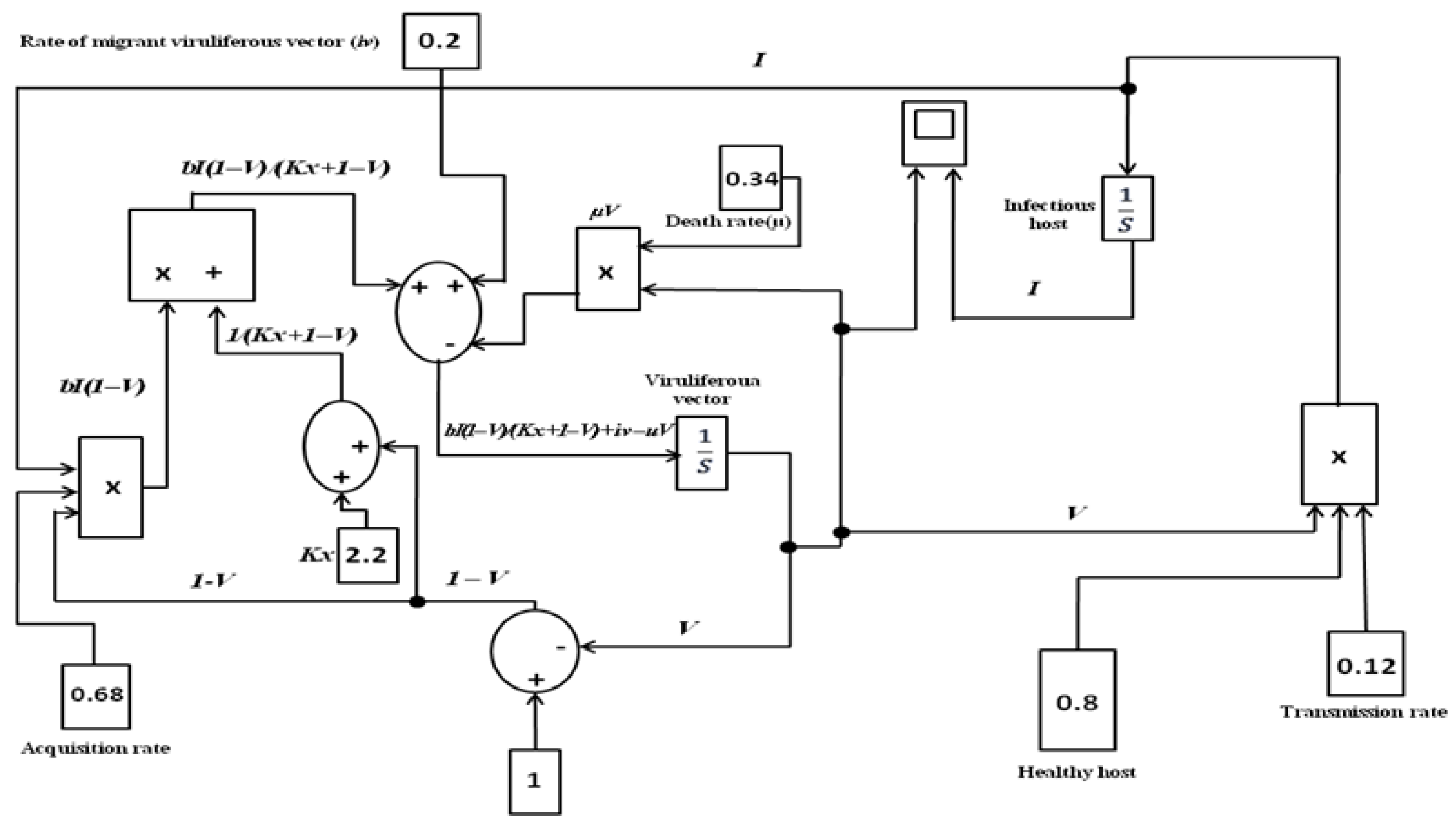
2.1. Prognostication of Major Driver in Chilli Leaf Curl Epidemic in Population Dynamic Modelling Framework
2.2. Field Implementation to Assess the Effect of Complete Interception of Migrant Viruliferous Vector Population on Leaf Curl Epidemic
2.2.1. Experimental Trial for Interception of Migrant Vectors
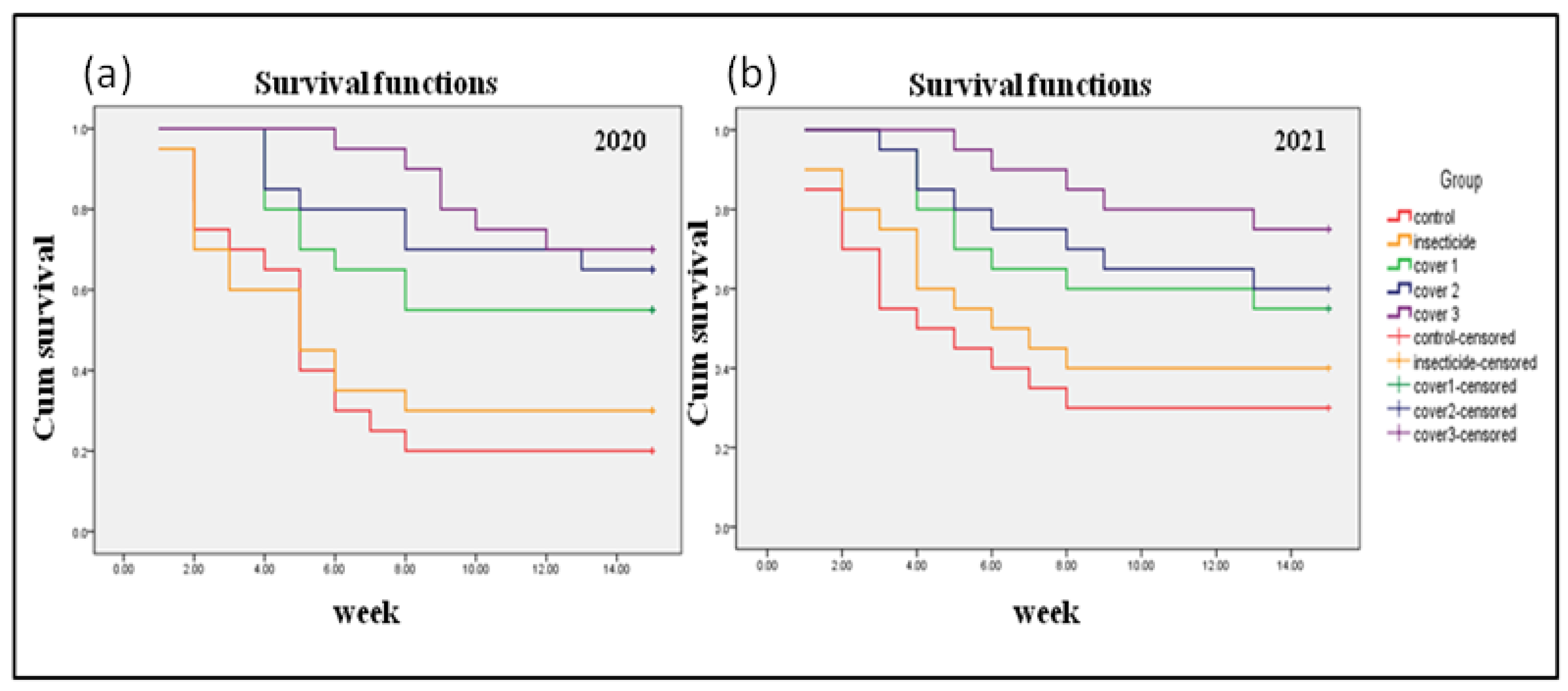
2.2.2. Survival Time (Plants Remain Uninfected by Virus) Analysis for Vector Interception
2.3. Estimation of Crop Growth Parameters and Fresh Fruit Yield under Interception
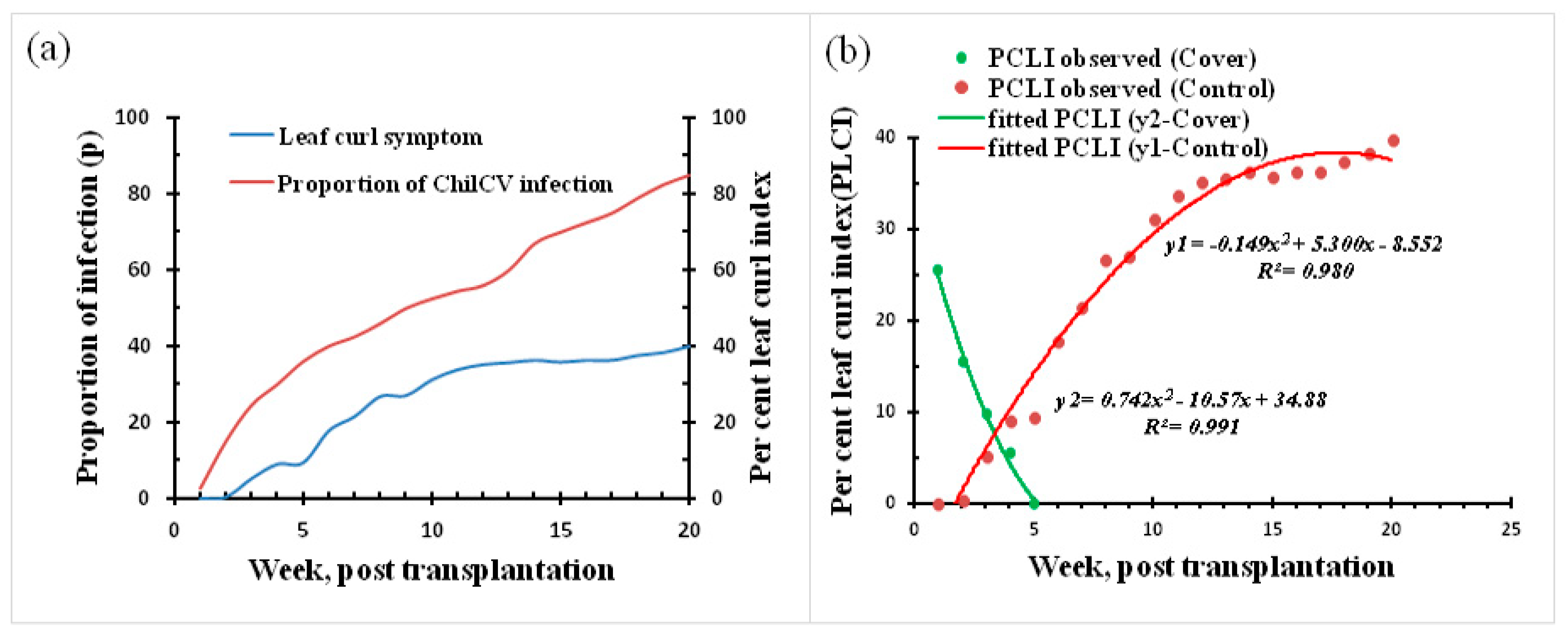
2.4. Leaf Curl Incidence in Terms of Characteristic Leaf Curl Symptom Components under Natural Field Conditions and Interception Covers
2.5. Eventual Assessment of Leaf Curl Incidence under Enhanced Protection in the Interception
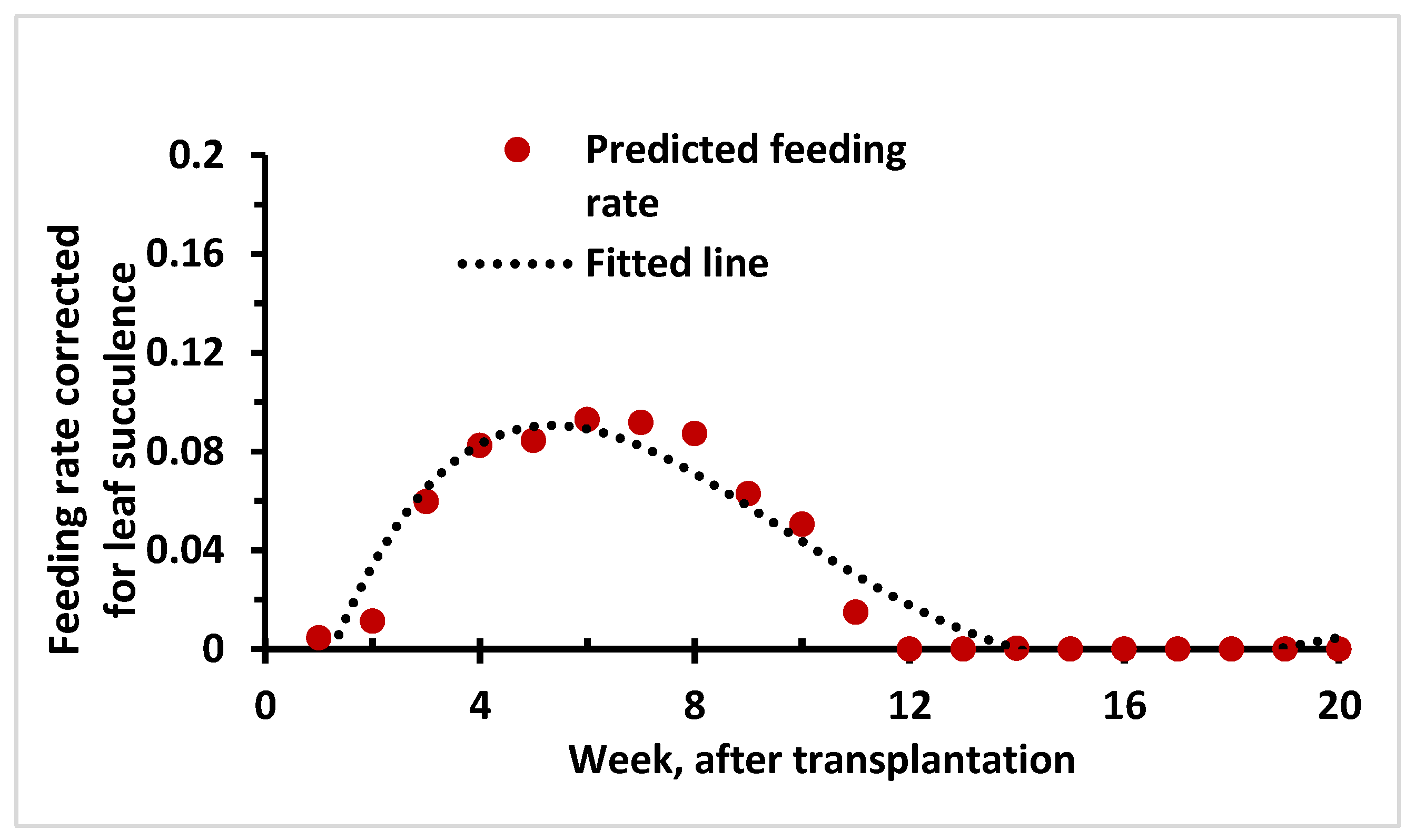
2.6. Prediction of Feeding Rate and Approximation of Contact Rate Dynamics Based on Host Density (Leaf Number and Leaf Area) and Succulence Level
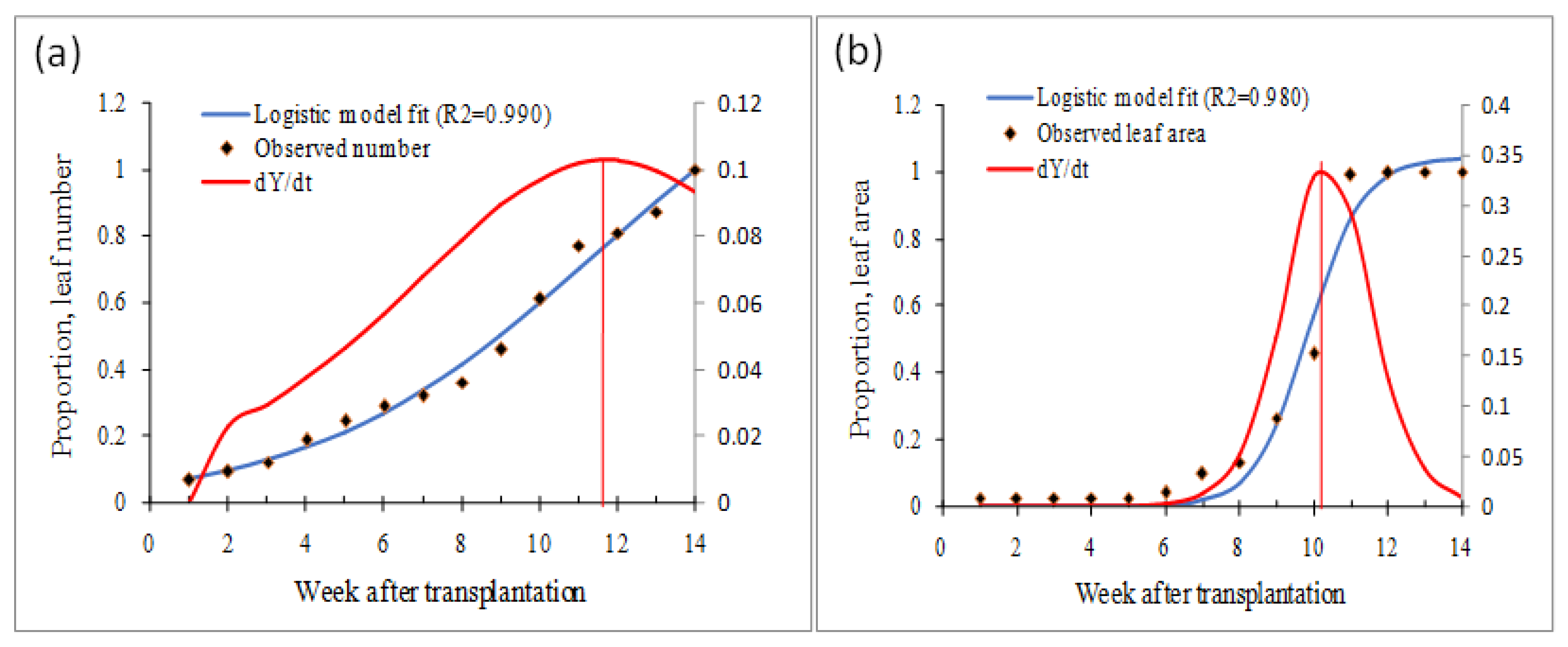
2.6.1. Estimation of Host Density (Number of Leaves and Area) and Succulence Level
2.6.2. Plant Age and Virus Transmission Potential
3. Results
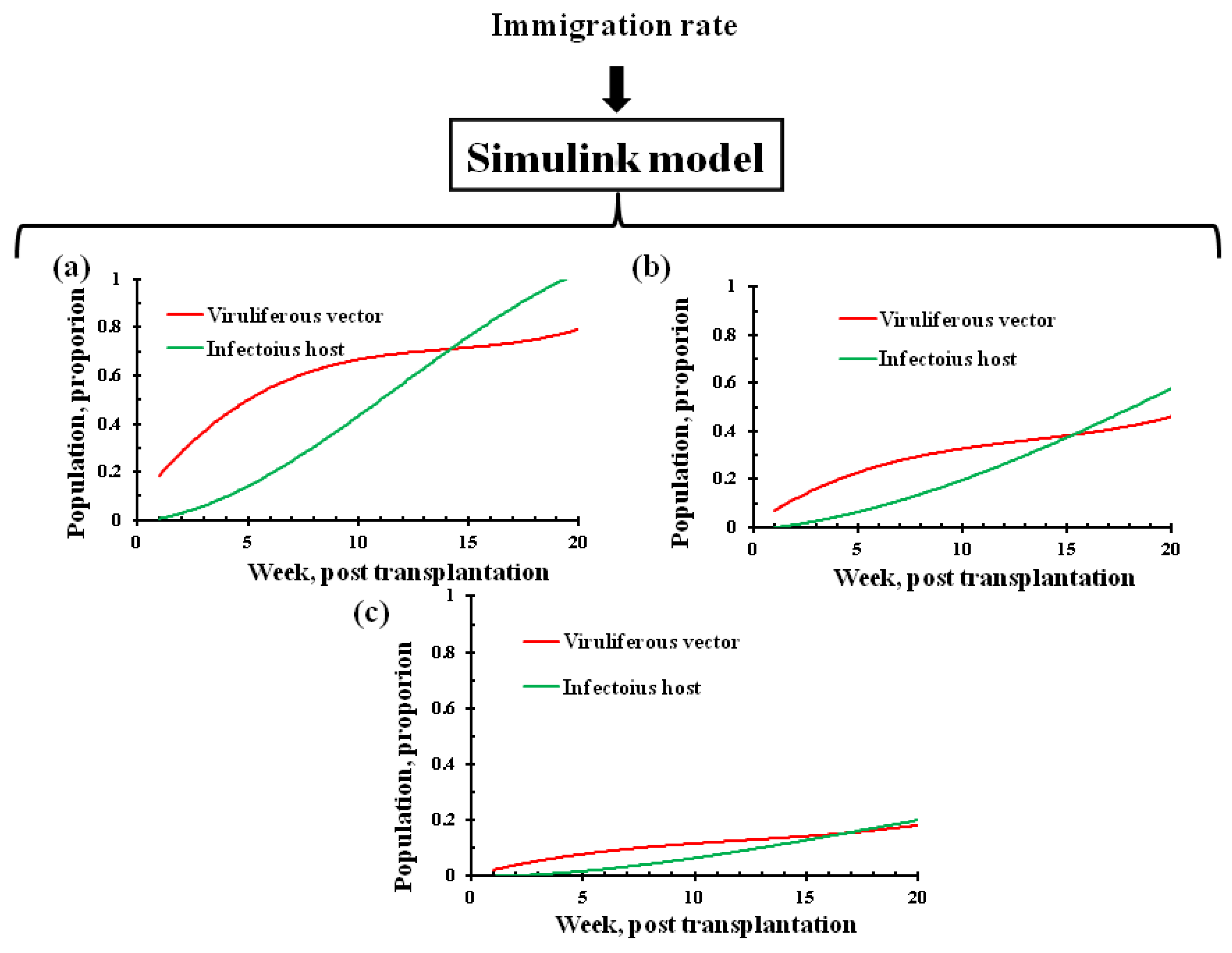
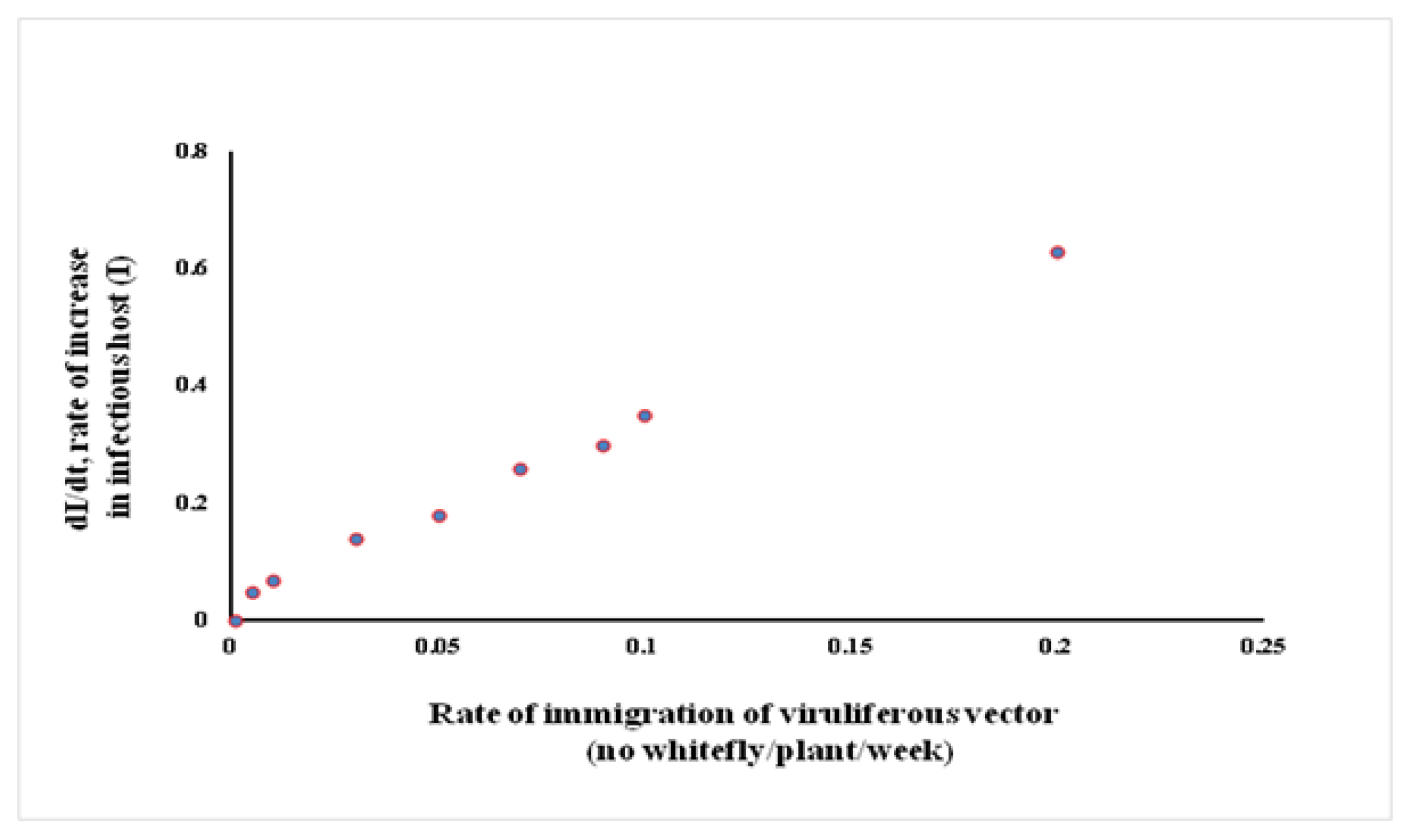
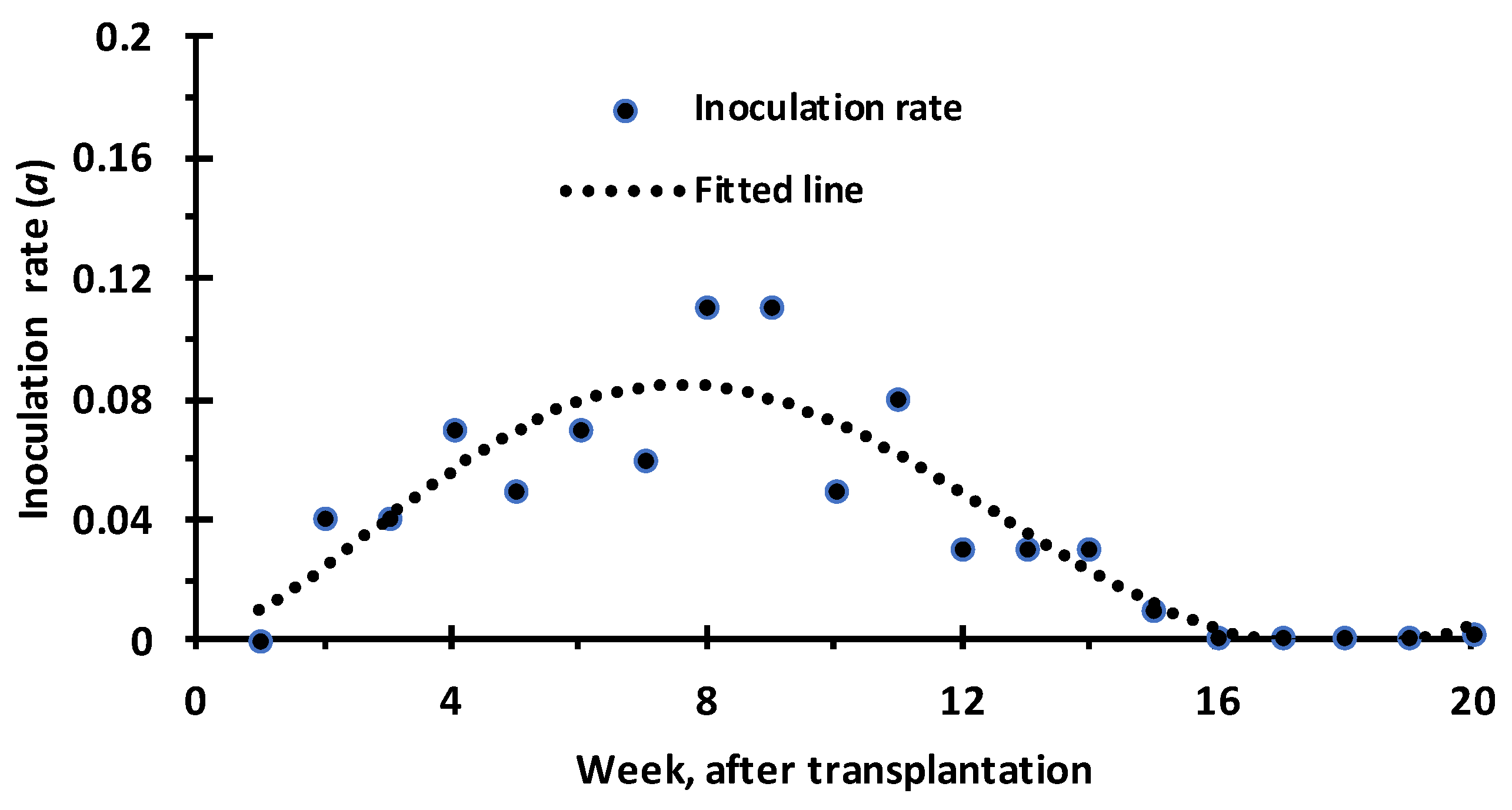
3.1. Prediction of Leaf Curl Epidemic in Response to Migration Rate Parameter
3.2. Field Assay for Complete Interception and Its Impact on Leaf Curl Epidemic in Chilli during the 2020 and 2021 Season
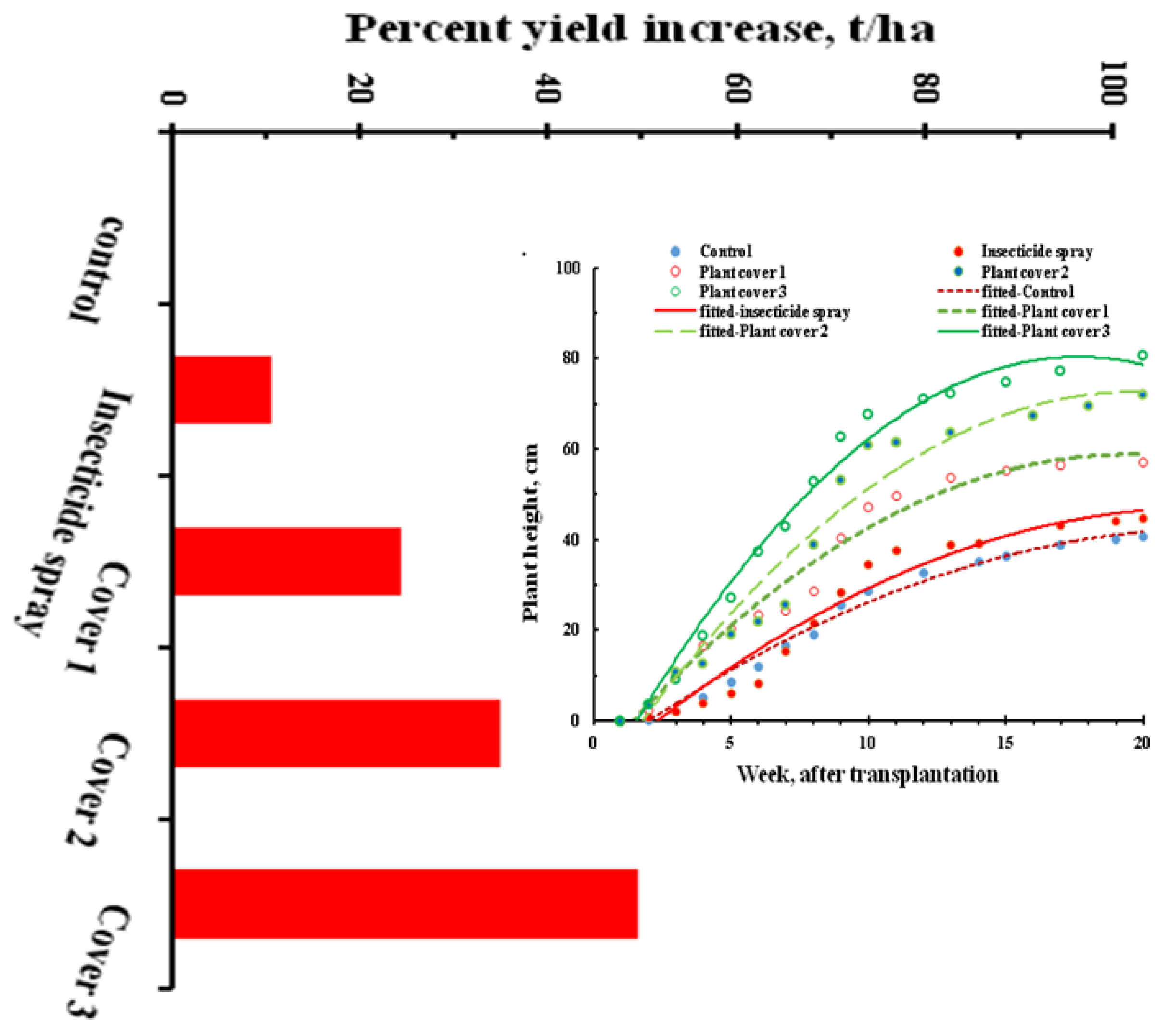
3.3. Assessment of Plant Growth Performance under Interception
3.4. Optimization of Interception Time for Field Implementation
3.5. Prediction of Vector Feeding Rate and Approximation of Contact Rate Dynamics Based on Host Density (Leaf Number and Leaf Area) and Succulence Level
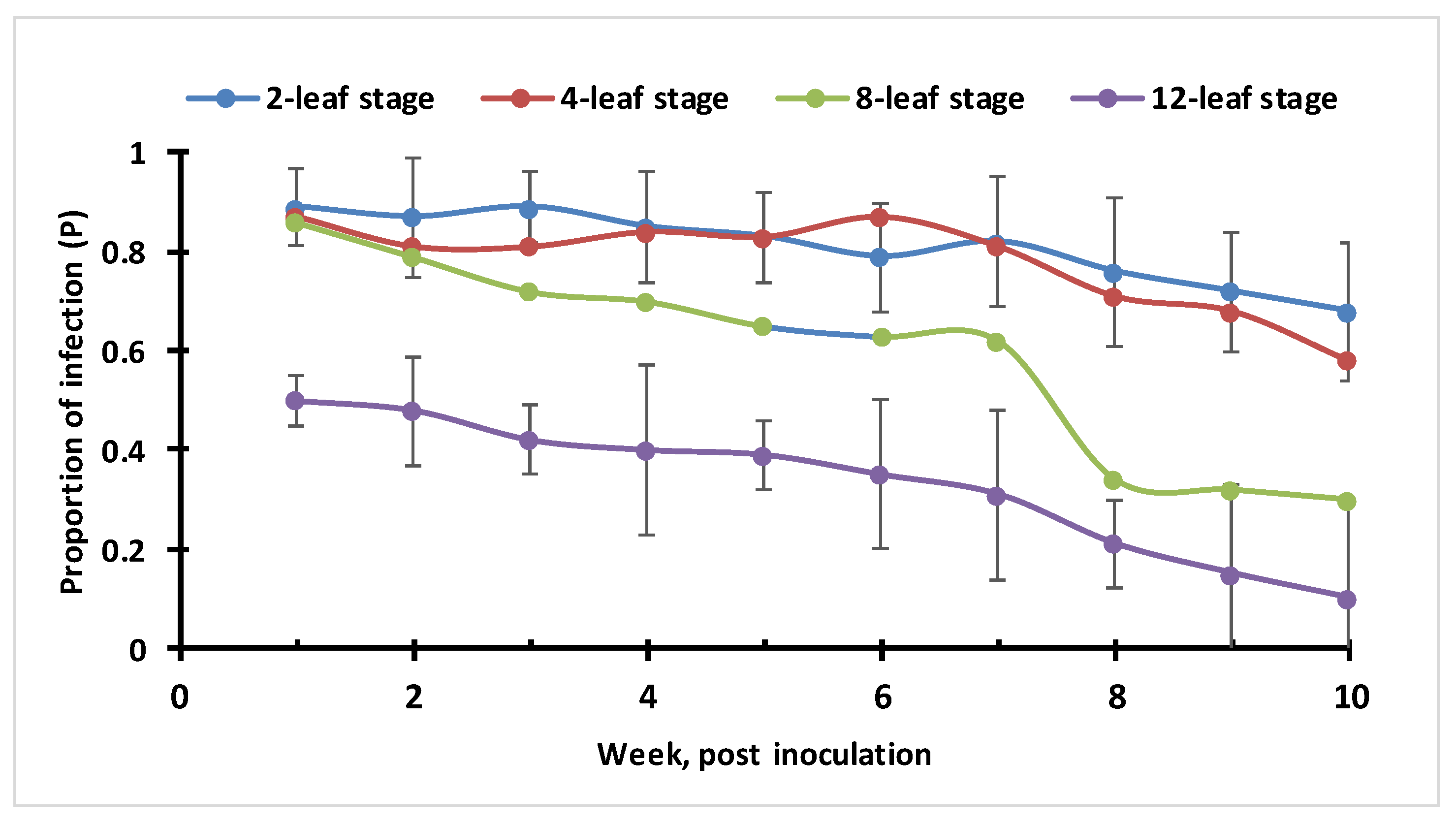
3.6. ChiLCV Transmission Potential in Plants with Different Ages (Leaf-Stage)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thakur, H.; Jindal, S.; Sharma, A.; Dhaliwal, M. Chilli leaf curl virus disease a serious threat for chilli cultivation. J. Plant Dis. Prot. 2018, 125, 239–249. [Google Scholar] [CrossRef]
- Prajapat, R.; Marwal, A.; Gaur, R.K. Begomovirus associated with alternative host weeds: A critical appraisal. Arch. Phytopathol. Pflanzenschutz. 2014, 47, 157–170. [Google Scholar] [CrossRef]
- Wisler, G.C.; Norris, R.F. Interactions between weeds and cultivated plants as related to management of plant pathogens. Weed Sci. 2005, 53, 914–917. [Google Scholar] [CrossRef]
- Duffus, J.E. Whitefly vectors: Increasing threat to world agriculture. In Proceedings of the XIX International Congress of Entomology, Beijing, China, 28 June–4 July1992; Thomas Say Publications in Entomology: Beijing, China, 1992; p. 352. [Google Scholar]
- Cohen, S. Epidemiology of whitefly—Transmitted viruses. In Bemisia: 1995. Taxonomy, Biology, Damage, Control and Management; Gerling, D., Mayer, R.T., Eds.; Intercept: London, UK, 1996; pp. 211–225. [Google Scholar]
- Polston, J.E.; Anderson, P.K. The emergence of whitefly-transmitted geminiviruses in tomato in Western Hemisphere. Plant Dis. 1997, 81, 1358–1369. [Google Scholar] [CrossRef] [Green Version]
- Mishra, G.P.; Singh, B.; Seth, T.; Singh, A.K.; Halder, J.; Krishnan, N.; Tiwari, S.K.; Singh, P.M. Biotechnological Advancements and Begomovirus Management in Okra (Abelmoschus esculentus L.): Status and Perspectives. Front. Plant Sci. 2017, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Capsicum annuum and Capsicum frutescens (Bell pepper, Chilli, Pepper, Sweet pepper). In Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019. [Google Scholar] [CrossRef]
- Menike, G.D.N.; De Costa, D.M. Variation of field symptoms and molecular diversity of the virus isolates associated with chilli leaf curl complex in different agroecological regions of Sri Lanka. Trop. Agric. Res. 2017, 28, 144–161. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; Ghosh, A. Topical spray of dsRNA induces mortality and inhibits chilli leaf curl virus transmission by Bemisia tabaci Asia II 1. Cells 2022, 11, 833. [Google Scholar] [CrossRef]
- Roy, B.; Dubey, S.; Ghosh, A.; Shukla, S.; Mandal, B.; Sinha, P. Simulation of leaf curl disease dynamics in chilli for strategic management options. Sci. Rep. 2021, 11, 1010. [Google Scholar] [CrossRef]
- Zou, C.; Shu, Y.N.; Yang, J.J.; Pan, L.L.; Zhao, J.; Chen, N.; Liu, S.S.; Wang, X.W. Begomovirus-Associated Betasatellite Virulence Factor βC1 Attenuates Tobacco Defense to Whiteflies via Interacting With Plant SKP1. Front. Plant Sci. 2020, 11, 574557. [Google Scholar] [CrossRef]
- Pan, L.L.; Miao, H.Y.; Wang, Q.M.; Walling, L.L.; Liu, S.S. Virus-induced phytohormone dynamics and their effects on plant-insect interactions. New Phytol. 2021, 230, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhang, Y.; Ma, W. The probing and feeding process of the greenhouse whitefly, Trzaleurodes vaporarzorum Westwood. Entomol. Sin. 1991, 1, 67–76. [Google Scholar] [CrossRef]
- Zhang, X.S.; Colvin, H.J. A general model of plant-virus disease infection incorporating vector aggregation. Plant Pathol. 2001, 49, 435–444. [Google Scholar] [CrossRef]
- Cunniffe, N.J.; Taylor, N.P.; Hamelin, F.M.; Jeger, M.J. Epidemiological and ecological consequences of virus manipulation of host and vector in plant virus transmission. PLoS Comput. Biol. 2021, 17, e1009759. [Google Scholar] [CrossRef]
- Cohen, S. Epidemiology of Whitefly-Transmitted Viruses. In Whiteflies, Their Bionomics, Pest Status and Management; Gerling, D., Ed.; Intercept: Andover, UK, 1990; pp. 211–225. [Google Scholar]
- Cohen, S.; Antignus, Y. Tomato yellow leaf curl virus, a whitefly-borne geminivirus of tomatoes. Adv. Dis. Vector Res. 1994, 10, 259–288. [Google Scholar] [CrossRef]
- Lapidot, M.; Polston, J.E. Biology and Epidemiology of Bemisia-Vectored Viruses. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 227–231. [Google Scholar] [CrossRef]
- Thongsripong, P.; Hyman, J.M.; Kapan, D.D.; Bennett, S.N. Human-mosquito Contact: A Missing Link in Our Understanding of Mosquito-Borne Disease Transmission Dynamics. Ann. Entomol. Soc. Am. 2021, 114, 397–414. [Google Scholar] [CrossRef]
- Jones, R.A.C. Trends in plant virus epidemiology: Opportunities from new or improved technologies. Virus Res. 2014, 186, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Jeger, M.J.; Bosch, F.V.D.; Madden, L.V.; Holt, J.K. A model for analysing plant-virus transmission characteristics and epidemic development. IMA J. Math. Appl. Med. Biol. 1998, 15, 1–18. [Google Scholar] [CrossRef]
- Chitnis, N.; Hyman, J.M.; Cushing, J.M. Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model. Bull. Math. Biol. 2008, 70, 1272–1296. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.M.; Garcia, A.J.; Focks, D.A.; Morrison, A.C.; Scott, T.W. Parameterization and sensitivity analysis of a complex simulation model for mosquito population dynamics, dengue transmission, and their control. Am. J. Trop. Med. Hyg. 2011, 85, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Antonovics, J.; Iwasa, Y.; Hassell, M.P. A generalized model of parasitoid, venereal, and vector-based transmission processes. Am Nat. 1995, 145, 661–675. [Google Scholar] [CrossRef]
- Miller, E.; Dushoff, J.; Huppert, A. The risk of incomplete personal protection coverage in vector-borne disease. J. R. Soc. Interface 2016, 13, 20150666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, J.; Bewick, S.; Calabrese, J.; Fagan, W.F. Dynamic modelling of personal protection control strategies for vector-borne disease limits the role of diversity amplification. J. R. Soc. Interface 2018, 15, 20180166. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Nakasuji, R.; Miyai, S.; Kawamoto, H.; Kiritani, K. Mathematical epidemiology of rice dwarf virus transmitted by green rice leafhoppers: A differential equation model. J. Appl. Ecol. 1985, 22, 839–847. [Google Scholar] [CrossRef]
- Chan, M.; Jeger, M.J. An analytical model of plant virus disease dynamics with roguing and replanting. J. Appl. Ecol. 1994, 3, 413–427. [Google Scholar] [CrossRef]
- Holt, J.K.; Jeger, M.J.; Thresh, J.M.; Otim-Nape, G.W. An Epidemiological Model Incorporating Vector Population Dynamics Applied to African Cassava Mosaic Virus Disease. J. Appl. Ecol. 1997, 34, 793–806. [Google Scholar] [CrossRef]
- Holt, J.; Colvin, J.; Muniyappa, V. Identifying control strategies for tomato leaf curl virus disease using an epidemiological model. J. Appl. Ecol. 1999, 36, 625–633. [Google Scholar] [CrossRef]
- Kinene, T.; Luboobi, L.; Nannyonga, B.; Mwanga, G.D. A mathematical model for the dynamics and cost effectiveness of the current controls of cassava brown streak disease in Uganda. J. Math. Comput. Sci. 2015, 5, 567–600. [Google Scholar]
- Jeger, M.; Chen, Z.; Cunningham, E.; Martin, G.; Powell, G. Population biology and epidemiology of plant virus epidemics: From tripartite to tritrophic interactions. Eur. J. Plant Pathol. 2012, 133, 3–23. [Google Scholar] [CrossRef]
- Cox, D.R. Regression models and life tables (with discussion). J. R. Stat. Soc. 1972, 34, 187–202. [Google Scholar] [CrossRef]
- Hegde, D.M. Effect of soil moisture and nitrogen on plant water relations, mineral composition and productivity of bell pepper (Capsicum annuum). Indian J. Agron. 1989, 34, 30–34. [Google Scholar]
- Wankhede, B.N.; Morey, D.K. Effect of levels of irrigation, nitrogen and potash on the growth and yield of chilli. Punjab Rao Krishi Vidyapeeth Res. J. 1984, 8, 22–25. [Google Scholar]
- Wilcoxon, R.D.; Skovmand, B.; Atif, A.H. Evalaution of wheat cultivars for their ability to retard development of stem rust. Ann. Appl. Biol. 1975, 80, 275–281. [Google Scholar] [CrossRef]
- Bos, L. Crop losses caused by viruses. Crop Prot. 1982, 1, 263–282. [Google Scholar] [CrossRef]
- Hepworth, G. Exact Confidence Intervals for Proportions Estimated by Group Testing. Biometrics 1996, 52, 1134–1146. [Google Scholar] [CrossRef]
- Hughes, G.; Gottwald, T.R. Survey methods for assessment of citrus tristeza virus incidence. Phytopathology 1998, 88, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Chakraborty, P.; Ghosh, A. How many begomovirus copies are acquired and inoculated by its vector, whitefly (Bemisia tabaci) during feeding? PLoS ONE 2021, 16, e0258933. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.R.; Wilson, J.M.; Garrett, R.G.; Smith, P.R. ELISA indexing of commercial carnations for carnation mottle virus using a urease-antibody conjugate. Plant Pathol. 1985, 34, 467–471. [Google Scholar] [CrossRef]
- Walter, S.D.; Hildreth, S.W.; Beaty, B.J. estimation of infection rates in populations of organisms using pools of variable size. Am. J. Epidemiol. 1980, 112, 124–128. [Google Scholar] [CrossRef]
- Thomas, R.; Thieffry, D.; Kaufman, M. Dynamical behavior of biological regulatory networks. 1. Biological role of feedback loops and practical use of the concept of the loop-characteristic state. Bull. Math. Biol. 1995, 57, 247–276. [Google Scholar] [CrossRef]
- Crespi, B.J. Vicious circles: Positive feedback in major evolutionary and ecological transitions. Trends Ecol. Evol. 2004, 19, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Zamberletti, P.; Sabir, K.; Opitz, T.; Bonnefon, O.; Gabriel, E.; Papaix, J. More pests but less pesticide applications: Ambivalent effect of landscape complexity on conservation biological control. PLOS Comput. Biol. 2021, 17, e1009559. [Google Scholar] [CrossRef] [PubMed]
- Devi, O.P.; Devi, K.S. Viral diseases in king chilli: A brief report. AgriCos E-Newsl. 2020, 2, 60–62. [Google Scholar]
- Zehra, S.B.; Ahmad, A.; Sharma, A.; Sofi, S.; Lateef, A.; Bashir, Z.; Husain, M.; Rathore, J.P. Chilli leaf curl virus an emerging threat to chilli in India. Int. J. Pure Appl. Biosci. 2017, 5, 404–414. [Google Scholar] [CrossRef]
- Rabbinge, R.; Rijsdijk, F.H. Disease and crop physiology: A modeler’s point of view. In Effects of Disease on the Physiology of the Growing Plant; Ayres, P.G., Ed.; Cambridge University Press: Cambridge, UK, 1981; pp. 201–220. [Google Scholar]
- Boote, K.J.; Jones, J.W.; Mishoe, J.W.; Berger, R.D. Coupling pests to crop growth simulators to predict yield reductions. Phytopathology 1983, 73, 1581–1587. [Google Scholar] [CrossRef]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-virus interactions in a plant host and in a hemipteran vector: Implications for vector fitness and virus epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explanatory Variable (Groups of Treatments) | 2020 | 2021 | ||||||
|---|---|---|---|---|---|---|---|---|
| Hazard Ratio | Standard Error | 95% CI | p-Value | Hazard Ratio | Standard Error | 95% CI | p-Value | |
| Control (without any treatment) | ||||||||
| Insecticide spray (imidacloprid 0.01%) | 5.60 | 0.52 | 2.02–15.52 | 0.001 | 5.02 | 0.52 | 1.80–13.99 | 0.002 |
| Plant cover (interception for 14 days) | 3.72 | 0.53 | 1.31–10.60 | 0.014 | 3.58 | 0.53 | 1.26–10.19 | 0.017 |
| Plant cover (interception for 21 days) | 2.23 | 0.55 | 0.74–6.67 | 0.15 | 2.12 | 0.55 | 0.71–6.32 | 0.178 |
| Plant cover (interception for 30 days) | 1.55 | 0.58 | 0.49–4.91 | 0.44 | 1.78 | 0.57 | 0.58–5.44 | 0.312 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, B.; Venu, E.; Kumar, S.; Dubey, S.; Lakshman, D.; Mandal, B.; Sinha, P. Leaf Curl Epidemic Risk in Chilli as a Consequence of Vector Migration Rate and Contact Rate Dynamics: A Critical Guide to Management. Viruses 2023, 15, 854. https://doi.org/10.3390/v15040854
Roy B, Venu E, Kumar S, Dubey S, Lakshman D, Mandal B, Sinha P. Leaf Curl Epidemic Risk in Chilli as a Consequence of Vector Migration Rate and Contact Rate Dynamics: A Critical Guide to Management. Viruses. 2023; 15(4):854. https://doi.org/10.3390/v15040854
Chicago/Turabian StyleRoy, Buddhadeb, Emmadi Venu, Sathiyaseelan Kumar, Shailja Dubey, Dilip Lakshman, Bikash Mandal, and Parimal Sinha. 2023. "Leaf Curl Epidemic Risk in Chilli as a Consequence of Vector Migration Rate and Contact Rate Dynamics: A Critical Guide to Management" Viruses 15, no. 4: 854. https://doi.org/10.3390/v15040854