
The Anti-Dengue Virus Peptide DV2 Inhibits Zika Virus Both In Vitro and In Vivo

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Lines and Virus
2.3. Peptide Synthesis
2.4. Cytotoxicity Tests
2.5. In Silico Docking of the DV2 Peptide
2.6. Virus Inhibition Assays
2.7. Flow Cytometry Analyses of ZIKV-Infected Cells
2.8. Mice Infection
2.9. Statistical Analysis
3. Results
3.1. In-Silico Docking Analyses of DV2 with the ZIKV E Protein
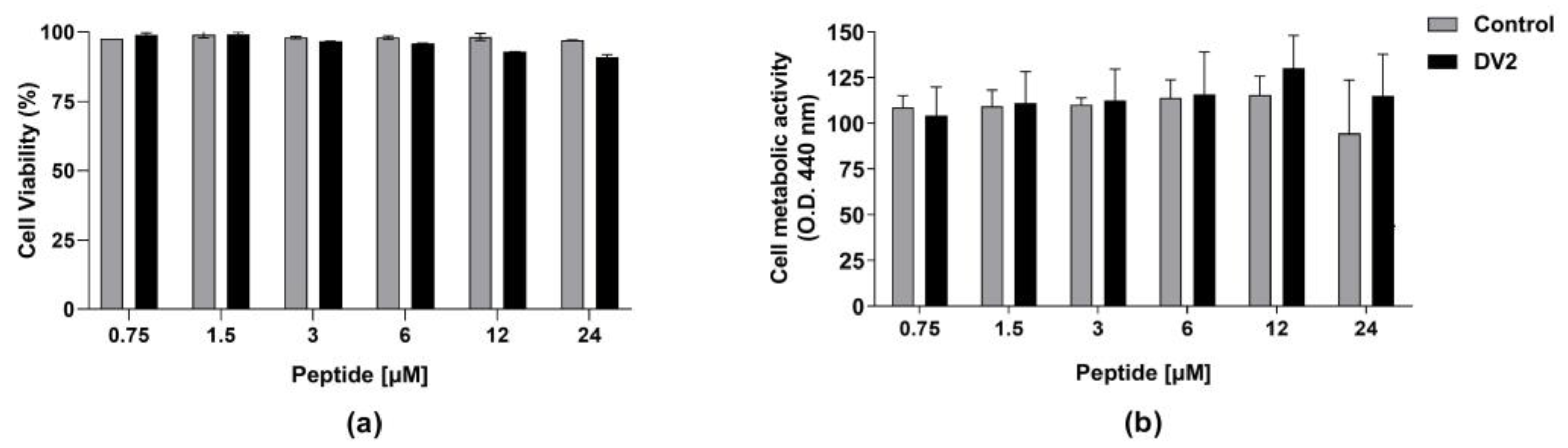
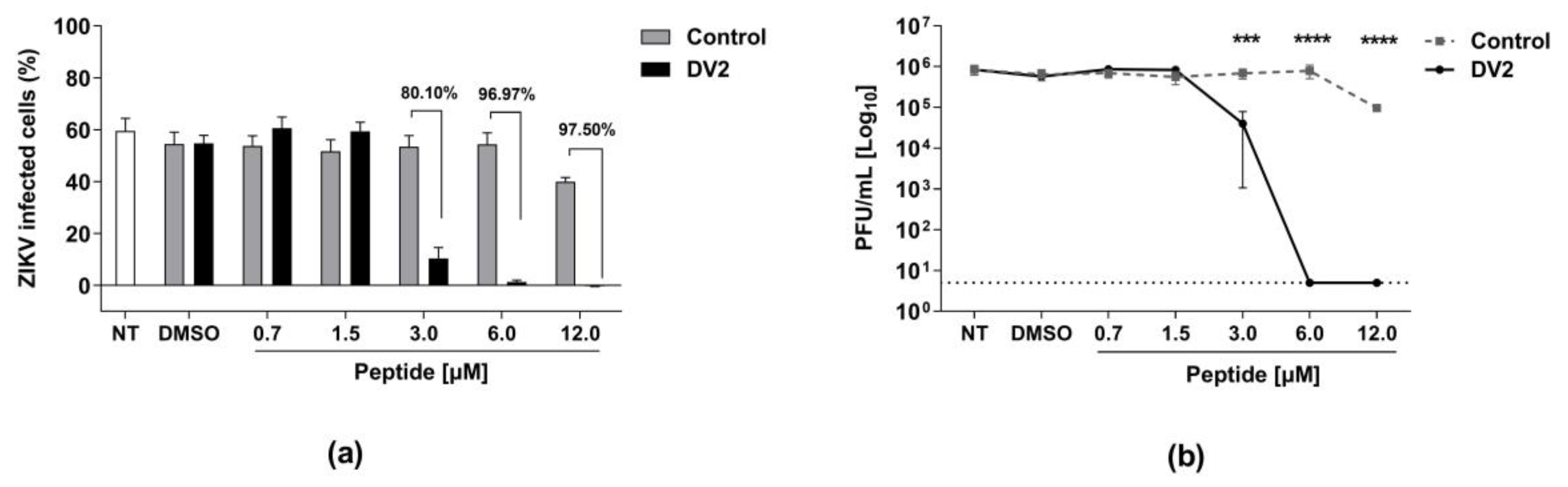
3.2. In Vitro Cytotoxicity and Anti-ZIKV Effects of the DV2 Peptide
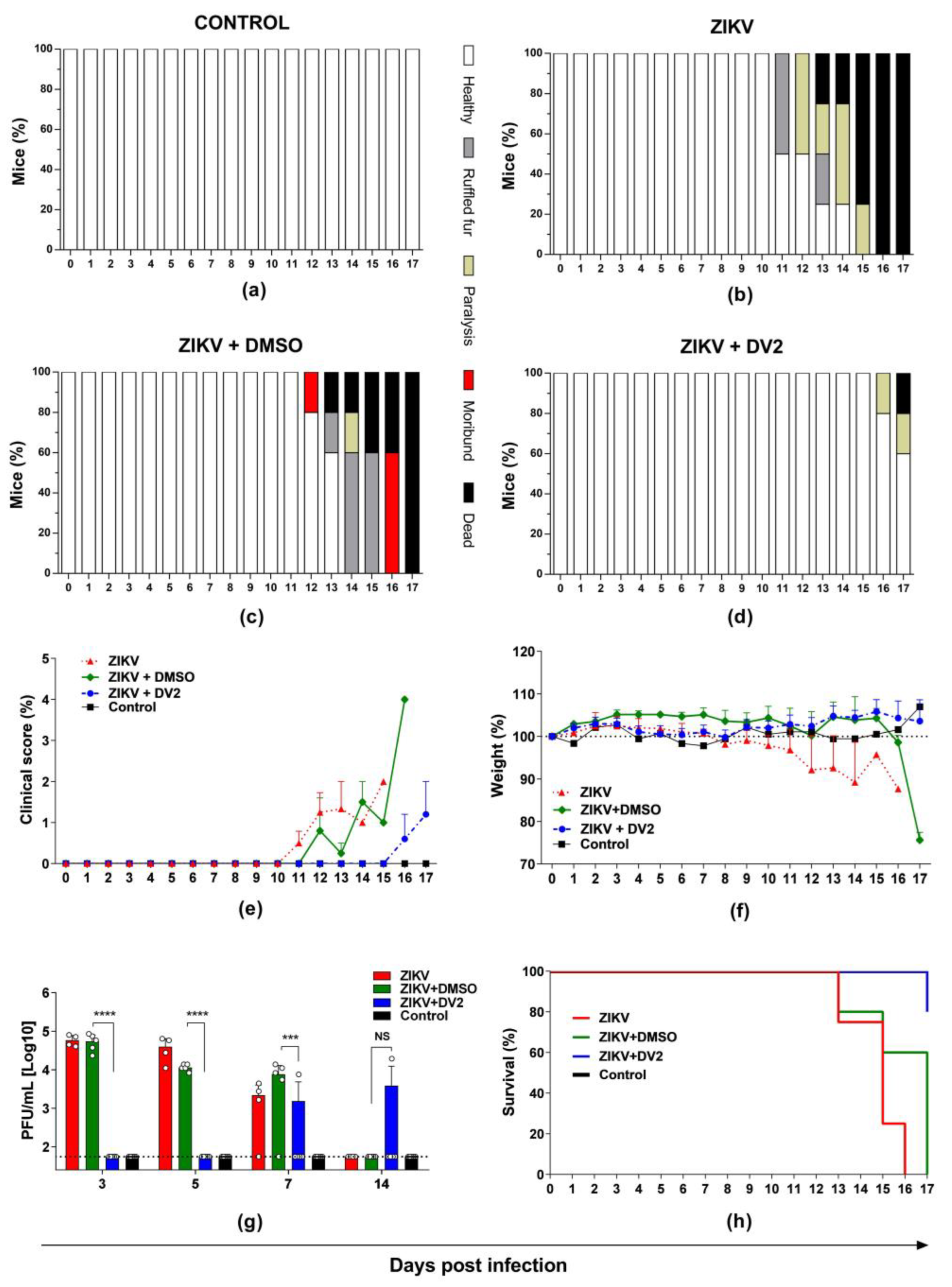
3.3. Protective Effects of the DV2 Peptide in AG129 Mice Challenged with ZIKV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus Infection—A Review of Immunopathogenesis, Immunological Response, and Immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef]
- Daep, C.A.; Muñoz-Jordán, J.L.; Eugenin, E.A. Flaviviruses, an Expanding Threat in Public Health: Focus on Dengue, West Nile, and Japanese Encephalitis Virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.K.; Bhattacharjee, S. Dengue Virus: Epidemiology, Biology, and Disease Aetiology. Can. J. Microbiol. 2021, 67, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The Global Economic Burden of Dengue: A Systematic Analysis. Lancet Infect Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Pielnaa, P.; Al-Saadawe, M.; Saro, A.; Dama, M.F.; Zhou, M.; Huang, Y.; Huang, J.; Xia, Z. Zika Virus-Spread, Epidemiology, Genome, Transmission Cycle, Clinical Manifestation, Associated Challenges, Vaccine and Antiviral Drug Development. Virology 2020, 543, 34–42. [Google Scholar] [CrossRef]
- Faria, N.R.; Quick, J.; Claro, I.M.; Thézé, J.; de Jesus, J.G.; Giovanetti, M.; Kraemer, M.U.G.; Hill, S.C.; Black, A.; da Costa, A.C.; et al. Establishment and Cryptic Transmission of Zika Virus in Brazil and the Americas. Nature 2017, 546, 406–410. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, W.K.; de França, G.V.A.; Carmo, E.H.; Duncan, B.B.; de Souza Kuchenbecker, R.; Schmidt, M.I. Infection-Related Microcephaly after the 2015 and 2016 Zika Virus Outbreaks in Brazil: A Surveillance-Based Analysis. Lancet 2017, 390, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An Update on Zika Virus Infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef] [Green Version]
- Miner, J.J.; Diamond, M.S. Zika Virus Pathogenesis and Tissue Tropism. Cell Host Microbe 2017, 21, 134–142. [Google Scholar] [CrossRef] [Green Version]
- França, G.V.A.D.; Pedi, V.D.; Garcia, M.H.D.O.; Carmo, G.M.I.D.; Leal, M.B.; Garcia, L.P. Síndrome congênita associada à infecção pelo vírus Zika em nascidos vivos no Brasil: Descrição da distribuição dos casos notificados e confirmados em 2015-2016. Epidemiol. Serv. Saúde 2018, 27, e2017473. [Google Scholar] [CrossRef] [Green Version]
- Richner, J.M.; Diamond, M.S. Zika Virus Vaccines: Immune Response, Current Status, and Future Challenges. Curr. Opin. Immunol. 2018, 53, 130–136. [Google Scholar] [CrossRef]
- Pinheiro-Michelsen, J.R.; Souza, R. da S.O.; Santana, I.V.R.; da Silva, P. de S.; Mendez, E.C.; Luiz, W.B.; Amorim, J.H. Anti-Dengue Vaccines: From Development to Clinical Trials. Front. Immunol. 2020, 11, 1252. [Google Scholar] [CrossRef]
- Torres-Flores, J.M.; Reyes-Sandoval, A.; Salazar, M.I. Dengue Vaccines: An Update. BioDrugs 2022, 36, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Modjarrad, K.; Lin, L.; George, S.L.; Stephenson, K.E.; Eckels, K.H.; De La Barrera, R.A.; Jarman, R.G.; Sondergaard, E.; Tennant, J.; Ansel, J.L.; et al. Preliminary Aggregate Safety and Immunogenicity Results from Three Trials of a Purified Inactivated Zika Virus Vaccine Candidate: Phase 1, Randomised, Double-Blind, Placebo-Controlled Clinical Trials. Lancet 2018, 391, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Tebas, P.; Roberts, C.C.; Muthumani, K.; Reuschel, E.L.; Kudchodkar, S.B.; Zaidi, F.I.; White, S.; Khan, A.S.; Racine, T.; Choi, H.; et al. Safety and Immunogenicity of an Anti–Zika Virus DNA Vaccine. N. Engl. J. Med. 2021, 385, e35. [Google Scholar] [CrossRef]
- Gaudinski, M.R.; Houser, K.V.; Morabito, K.M.; Hu, Z.; Yamshchikov, G.; Rothwell, R.S.; Berkowitz, N.; Mendoza, F.; Saunders, J.G.; Novik, L.; et al. Safety, Tolerability, and Immunogenicity of Two Zika Virus DNA Vaccine Candidates in Healthy Adults: Randomised, Open-Label, Phase 1 Clinical Trials. Lancet 2018, 391, 552–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, K.E.; Tan, C.S.; Walsh, S.R.; Hale, A.; Ansel, J.L.; Kanjilal, D.G.; Jaegle, K.; Peter, L.; Borducchi, E.N.; Nkolola, J.P.; et al. Safety and Immunogenicity of a Zika Purified Inactivated Virus Vaccine given via Standard, Accelerated, or Shortened Schedules: A Single-Centre, Double-Blind, Sequential-Group, Randomised, Placebo-Controlled, Phase 1 Trial. Lancet Infect. Dis. 2020, 20, 1061–1070. [Google Scholar] [CrossRef]
- Han, H.-H.; Diaz, C.; Acosta, C.J.; Liu, M.; Borkowski, A. Safety and Immunogenicity of a Purified Inactivated Zika Virus Vaccine Candidate in Healthy Adults: An Observer-Blind, Randomised, Phase 1 Trial. Lancet Infect. Dis. 2021, 21, 1282–1292. [Google Scholar] [CrossRef]
- Essink, B.; Chu, L.; Seger, W.; Barranco, E.; Le Cam, N.; Bennett, H.; Faughnan, V.; Pajon, R.; Paila, Y.D.; Bollman, B.; et al. The Safety and Immunogenicity of Two Zika Virus MRNA Vaccine Candidates in Healthy Flavivirus Baseline Seropositive and Seronegative Adults: The Results of Two Randomised, Placebo-Controlled, Dose-Ranging, Phase 1 Clinical Trials. Lancet Infect. Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.Z.; Cheng, P.G.; Abdulrahman, A.Y.; Teoh, T.C. The Identification of Active Compounds in Ganoderma Lucidum Var. Antler Extract Inhibiting Dengue Virus Serine Protease and Its Computational Studies. J. Biomol. Struct. Dyn. 2020, 38, 4273–4288. [Google Scholar] [CrossRef] [PubMed]
- Norshidah, H.; Vignesh, R.; Lai, N.S. Updates on Dengue Vaccine and Antiviral: Where Are We Heading? Molecules 2021, 26, 6768. [Google Scholar] [CrossRef] [PubMed]
- Diosa-Toro, M.; Troost, B.; van de Pol, D.; Heberle, A.M.; Urcuqui-Inchima, S.; Thedieck, K.; Smit, J.M. Tomatidine, a Novel Antiviral Compound towards Dengue Virus. Antiviral Res. 2019, 161, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Quintana, V.M.; Selisko, B.; Brunetti, J.E.; Eydoux, C.; Guillemot, J.C.; Canard, B.; Damonte, E.B.; Julander, J.G.; Castilla, V. Antiviral Activity of the Natural Alkaloid Anisomycin against Dengue and Zika Viruses. Antiviral Res. 2020, 176, 104749. [Google Scholar] [CrossRef]
- Ruan, J.; Rothan, H.A.; Zhong, Y.; Yan, W.; Henderson, M.J.; Chen, F.; Fang, S. A Small Molecule Inhibitor of ER-to-Cytosol Protein Dislocation Exhibits Anti-Dengue and Anti-Zika Virus Activity. Sci. Rep. 2019, 9, 10901. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Kim, E.H.; O’neal, J.T.; Dale, G.; Holthausen, D.J.; Bowen, J.R.; Quicke, K.M.; Skountzou, I.; Gopal, S.; George, S.; et al. The Amphibian Peptide Yodha Is Virucidal for Zika and Dengue Viruses. Sci. Rep. 2021, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.C.; Ricardo, R.C.; Santos, N.C. Dengue, West Nile, and Zika Viruses: Potential Novel Antiviral Biologics Drugs Currently at Discovery and Preclinical Development Stages. Pharmaceutics 2022, 14, 2535. [Google Scholar] [CrossRef] [PubMed]
- Low, J.G.H.; Ooi, E.E.; Vasudevan, S.G. Current Status of Dengue Therapeutics Research and Development. J. Infect. Dis. 2017, 215, S96–S102. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, G.; Gabrani, R. Antiviral Peptides: Identification and Validation. Int. J. Pept. Res. Ther. 2021, 27, 149–168. [Google Scholar] [CrossRef]
- Lok, S.-M.; Costin, J.M.; Hrobowski, Y.M.; Hoffmann, A.R.; Rowe, D.K.; Kukkaro, P.; Holdaway, H.; Chipman, P.; Fontaine, K.A.; Holbrook, M.R.; et al. Release of Dengue Virus Genome Induced by a Peptide Inhibitor. PLoS ONE 2012, 7, e50995. [Google Scholar] [CrossRef]
- Chew, M.-F.; Poh, K.-S.; Poh, C.-L. Peptides as Therapeutic Agents for Dengue Virus. Int. J. Med. Sci. 2017, 14, 1342–1359. [Google Scholar] [CrossRef] [Green Version]
- Panya, A.; Bangphoomi, K.; Choowongkomon, K.; Yenchitsomanus, P. Peptide Inhibitors Against Dengue Virus Infection. Chem. Biol. Drug Des. 2014, 84, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.G.; Yang, P.L.; Harrison, S.C. Peptide Inhibitors of Flavivirus Entry Derived from the E Protein Stem. J. Virol. 2010, 84, 12549–12554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.G.; Yang, P.L.; Harrison, S.C. Peptide Inhibitors of Dengue-Virus Entry Target a Late-Stage Fusion Intermediate. PLoS Pathog. 2010, 6, e1000851. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, Y.; Wang, S.; Sun, J.; Wang, P.; Xin, Q.; Zhang, L.; Xiao, G.; Wang, W. Antiviral Activity of Peptide Inhibitors Derived from the Protein E Stem against Japanese Encephalitis and Zika Viruses. Antiviral Res. 2017, 141, 140–149. [Google Scholar] [CrossRef]
- Bai, F.; Town, T.; Pradhan, D.; Cox, J.; Ashish; Ledizet, M.; Anderson, J.F.; Flavell, R.A.; Krueger, J.K.; Koski, R.A.; et al. Antiviral Peptides Targeting the West Nile Virus Envelope Protein. J. Virol. 2007, 81, 2047–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Deng, Y.-Q.; Zou, P.; Wang, Q.; Dai, Y.; Yu, F.; Du, L.; Zhang, N.-N.; Tian, M.; Hao, J.-N.; et al. A Peptide-Based Viral Inactivator Inhibits Zika Virus Infection in Pregnant Mice and Fetuses. Nat. Commun. 2017, 8, 15672. [Google Scholar] [CrossRef]
- Faria, N.R.; Azevedo, R.D.S.D.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika Virus in the Americas: Early Epidemiological and Genetic Findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.R.; Alves, R.P.D.S.; Sales, N.S.; Andreata-Santos, R.; Venceslau-Carvalho, A.A.; Pereira, S.S.; Castro-Amarante, M.F.; Rodrigues-Jesus, M.J.; Favaro, M.T.D.P.; Chura-Chambi, R.M.; et al. Enhanced Immune Responses and Protective Immunity to Zika Virus Induced by a DNA Vaccine Encoding a Chimeric NS1 Fused with Type 1 Herpes Virus GD Protein. Front Med. Technol. 2020, 2, 604160. [Google Scholar] [CrossRef]
- Andreata-Santos, R.; Alves, R.P.D.S.; Pereira, S.A.; Pereira, L.R.; Freitas, C.L.D.; Pereira, S.S.; Venceslau-Carvalho, A.A.; Castro-Amarante, M.F.; Favaro, M.T.P.; Mathias-Santos, C.; et al. Transcutaneous Administration of Dengue Vaccines. Viruses 2020, 12, 514. [Google Scholar] [CrossRef]
- Šali, A.; Blundell, T.L. Comparative Protein Modelling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Kurcinski, M.; Pawel Ciemny, M.; Oleniecki, T.; Kuriata, A.; Badaczewska-Dawid, A.E.; Kolinski, A.; Kmiecik, S. CABS-Dock Standalone: A Toolbox for Flexible Protein–Peptide Docking. Bioinformatics 2019, 35, 4170–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, B.L.; Kelley, L.A.; Barber, J.; Murray, J.W.; MacDonald, J.T. High-Quality Protein Backbone Reconstruction from Alpha Carbons Using Gaussian Mixture Models. J. Comput. Chem. 2013, 34, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully Automated Protein–Ligand Interaction Profiler. Nucleic Acids Res 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Hameed, M.; Wang, X.; Zhang, J.; Guo, S.; Anwar, M.N.; Pang, L.; Liu, K.; Li, B.; Shao, D.; et al. Antiviral Activity of Phage Display-Selected Peptides against Japanese Encephalitis Virus Infection in Vitro and in Vivo. Antiviral Res. 2020, 174, 104673. [Google Scholar] [CrossRef]
- Mertinková, P.; Mochnáčová, E.; Bhide, K.; Kulkarni, A.; Tkáčová, Z.; Hruškovicová, J.; Bhide, M. Development of Peptides Targeting Receptor Binding Site of the Envelope Glycoprotein to Contain the West Nile Virus Infection. Sci. Rep. 2021, 11, 20131. [Google Scholar] [CrossRef]
- Zoladek, J.; Burlaud-Gaillard, J.; Chazal, M.; Desgraupes, S.; Jeannin, P.; Gessain, A.; Pardigon, N.; Hubert, M.; Roingeard, P.; Jouvenet, N.; et al. Human Claudin-Derived Peptides Block the Membrane Fusion Process of Zika Virus and Are Broad Flavivirus Inhibitors. Microbiol. Spectr. 2022, 10, e0298922. [Google Scholar] [CrossRef]
- Stiasny, K.; Kiermayr, S.; Bernhart, A.; Heinz, F.X. The Membrane-Proximal “Stem” Region Increases the Stability of the Flavivirus E Protein Postfusion Trimer and Modulates Its Structure. J. Virol. 2013, 87, 9933–9938. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Wu, Z.; Wu, S.; Chen, S.; Cheng, A. The Key Amino Acids of E Protein Involved in Early Flavivirus Infection: Viral Entry. Virol. J. 2021, 18, 136. [Google Scholar] [CrossRef]
- Aliota, M.T.; Caine, E.A.; Walker, E.C.; Larkin, K.E.; Camacho, E.; Osorio, J.E. Characterization of Lethal Zika Virus Infection in AG129 Mice. PLoS Negl. Trop. Dis. 2016, 10, e0004682. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (μM) | IC90 (μM) | |
|---|---|---|
| Flow Cytometry 1 | 2.647 | 3.228 |
| Plaque assay 2 | 1.976 | 2.696 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Amarante, M.F.d.; Pereira, S.S.; Pereira, L.R.; Santos, L.S.; Venceslau-Carvalho, A.A.; Martins, E.G.; Balan, A.; Souza Ferreira, L.C.d. The Anti-Dengue Virus Peptide DV2 Inhibits Zika Virus Both In Vitro and In Vivo. Viruses 2023, 15, 839. https://doi.org/10.3390/v15040839
Castro-Amarante MFd, Pereira SS, Pereira LR, Santos LS, Venceslau-Carvalho AA, Martins EG, Balan A, Souza Ferreira LCd. The Anti-Dengue Virus Peptide DV2 Inhibits Zika Virus Both In Vitro and In Vivo. Viruses. 2023; 15(4):839. https://doi.org/10.3390/v15040839
Chicago/Turabian StyleCastro-Amarante, Maria Fernanda de, Samuel Santos Pereira, Lennon Ramos Pereira, Lucas Souza Santos, Alexia Adrianne Venceslau-Carvalho, Eduardo Gimenes Martins, Andrea Balan, and Luís Carlos de Souza Ferreira. 2023. "The Anti-Dengue Virus Peptide DV2 Inhibits Zika Virus Both In Vitro and In Vivo" Viruses 15, no. 4: 839. https://doi.org/10.3390/v15040839