
The RL13 Temperance Factor Represses Replication of the Highly Cell Culture-Adapted Towne Strain of Human Cytomegalovirus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses
2.3. Focus Size Determinations
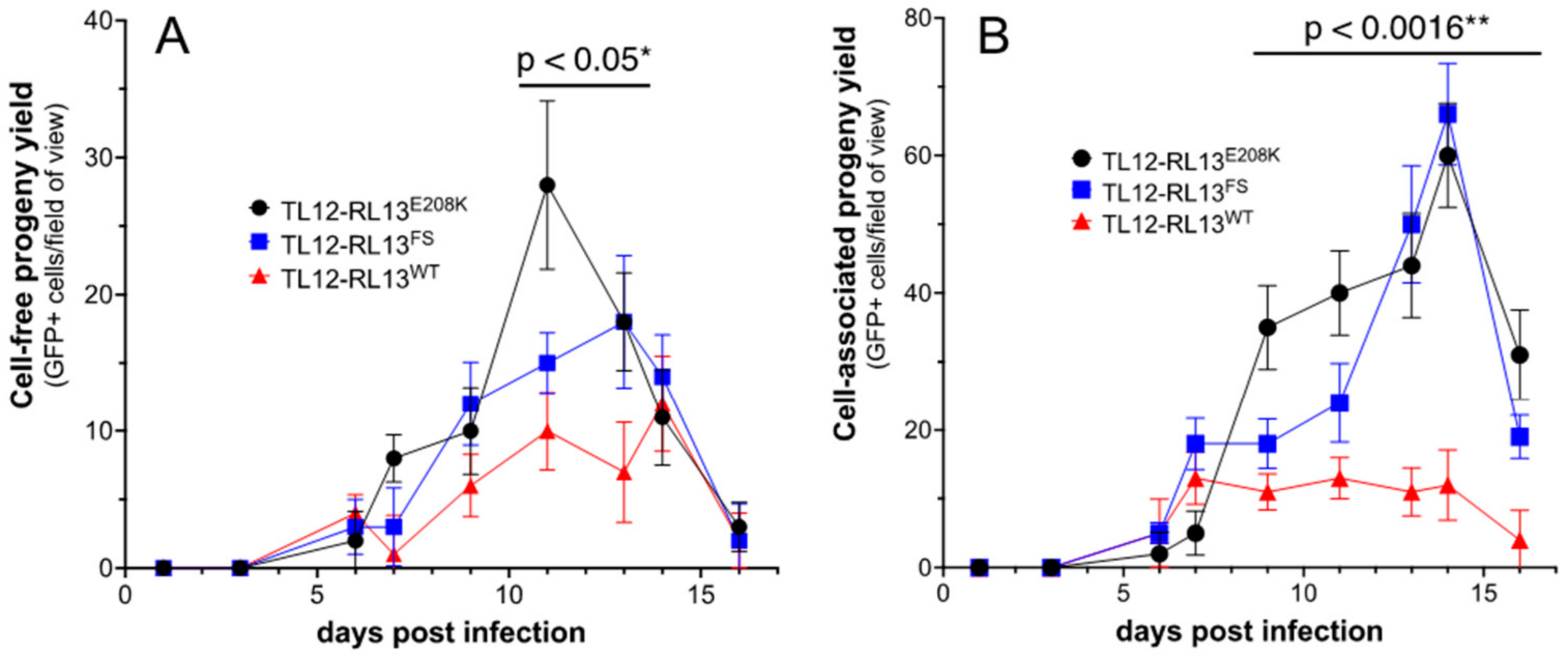
2.4. Growth Curves
2.5. Immunofluorescence
2.6. Western Immunoblotting
2.7. Sequencing, Sequence Analyses, Alignments, and Statistical Methods
3. Results
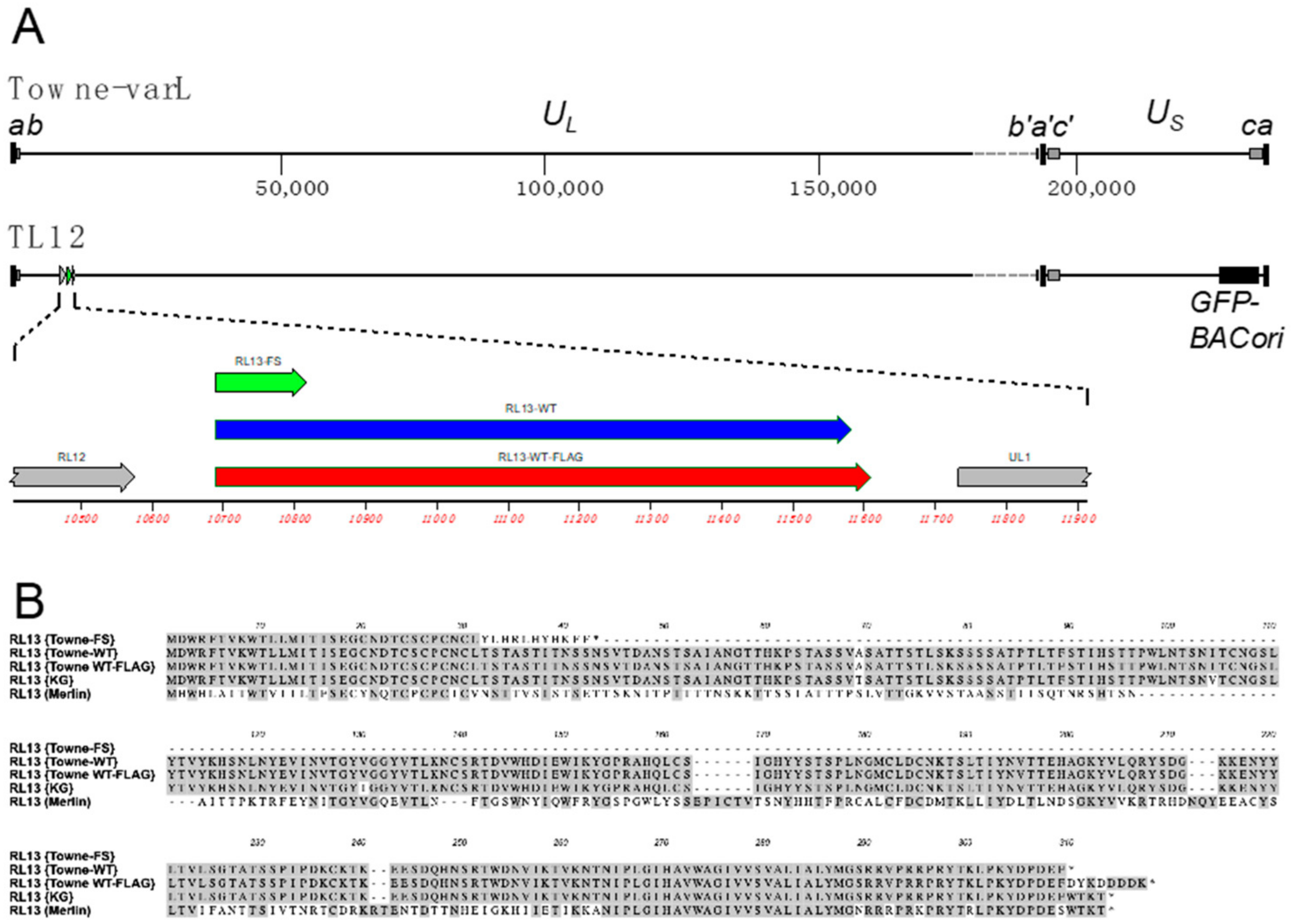
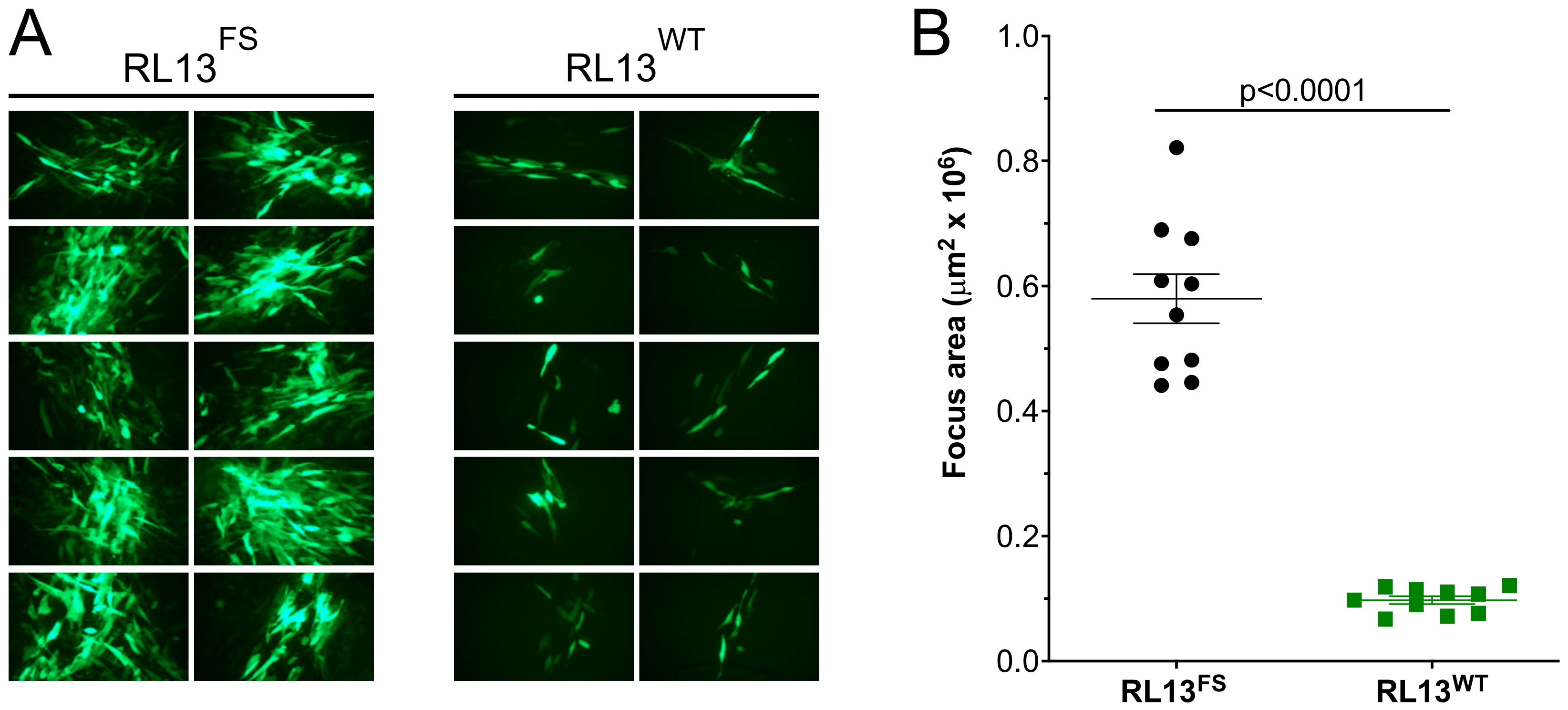
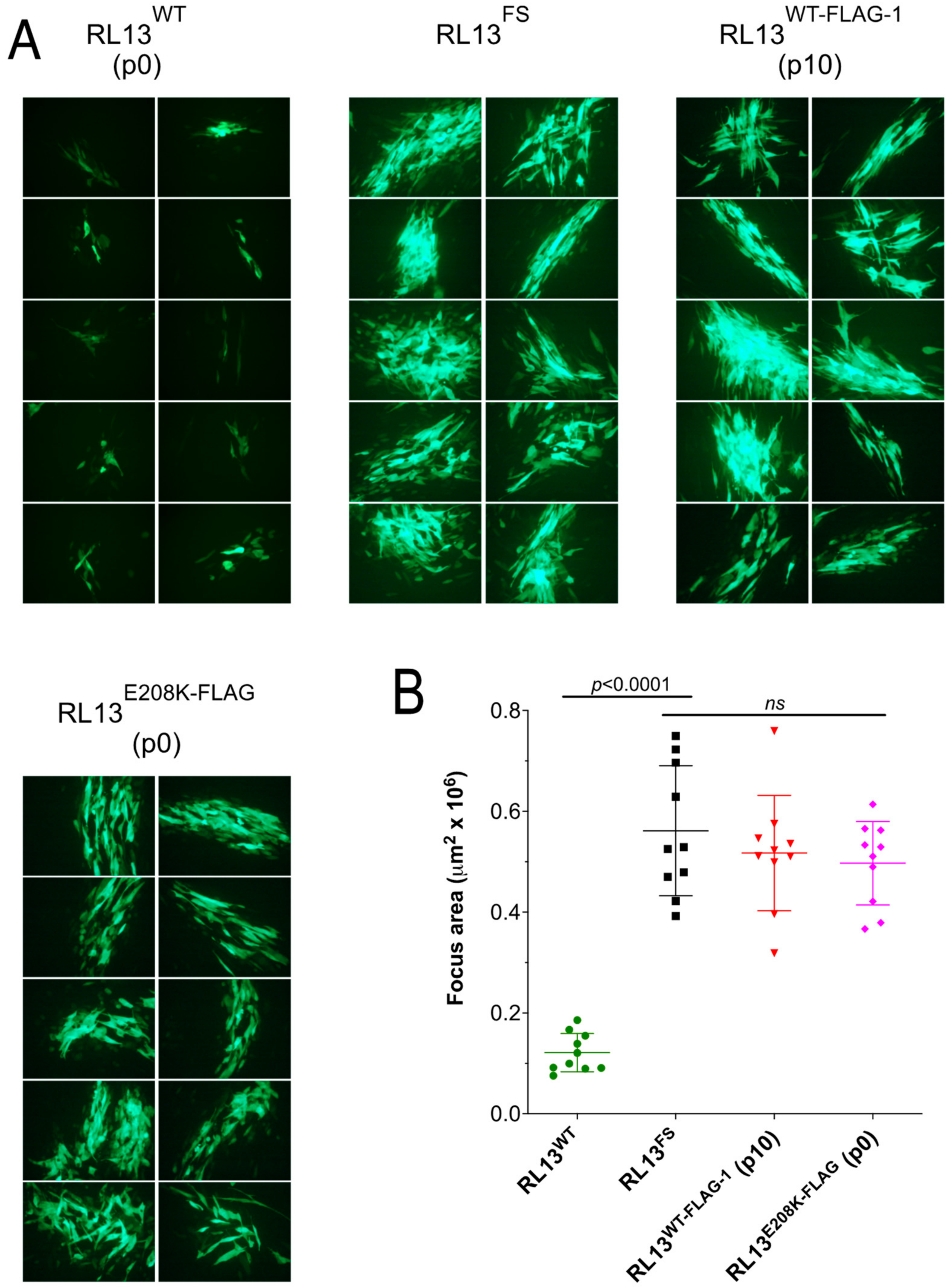
3.1. Repair of the RL13 Mutation in TL12 Results in Reduced Focus Size
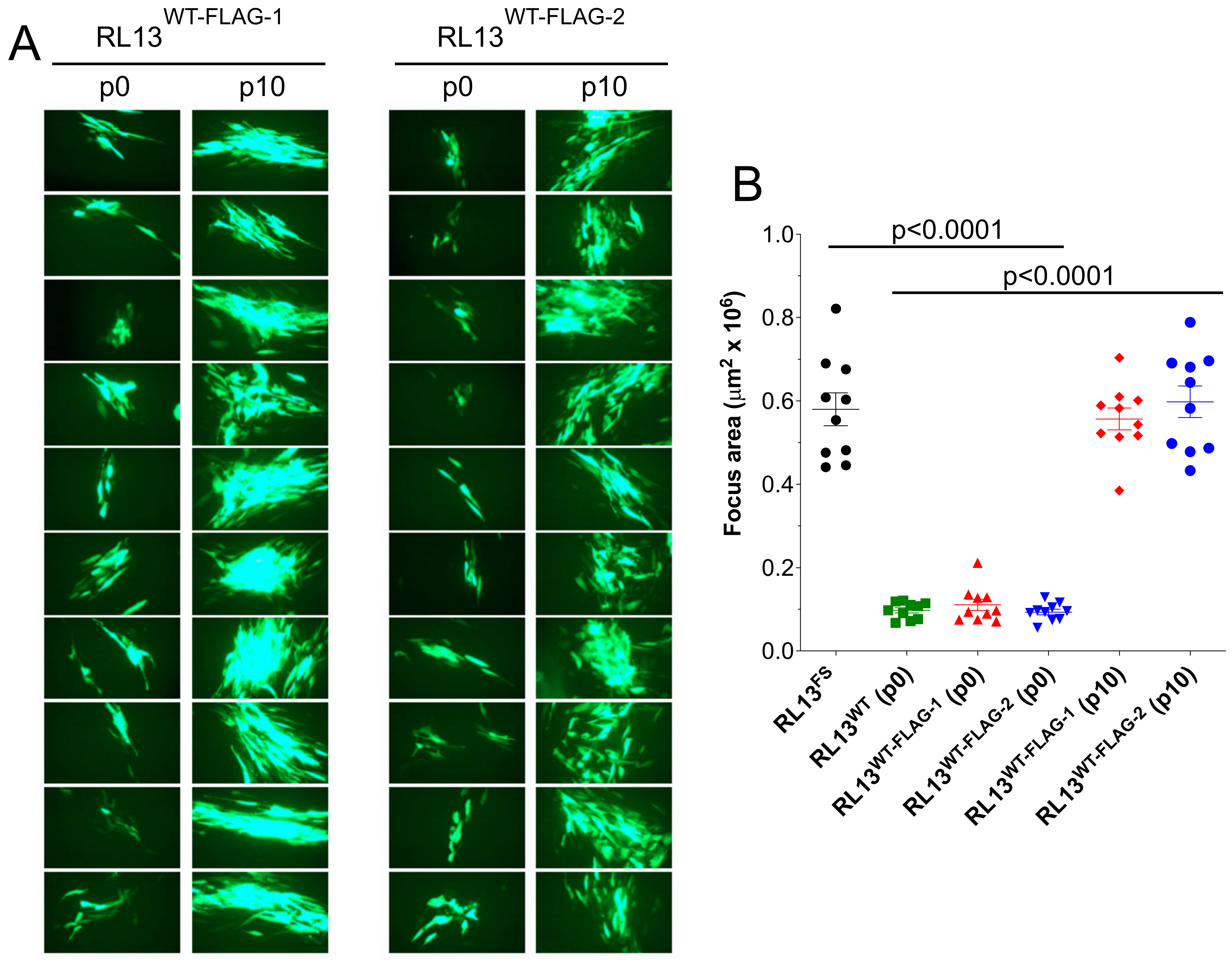
3.2. RL13 in TL12-RL13WT Mutates after a Few Passages
3.3. RL13-FLAG Is Functional
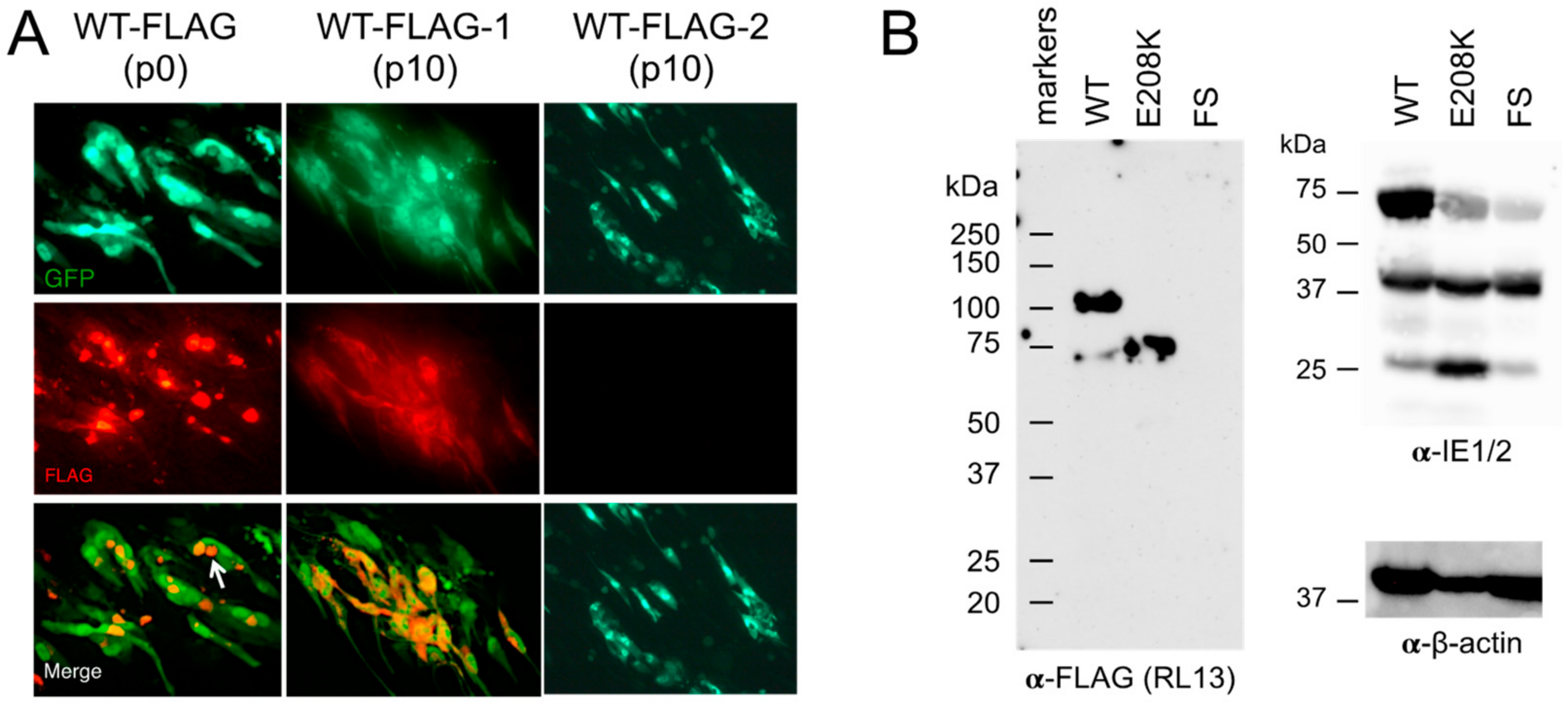
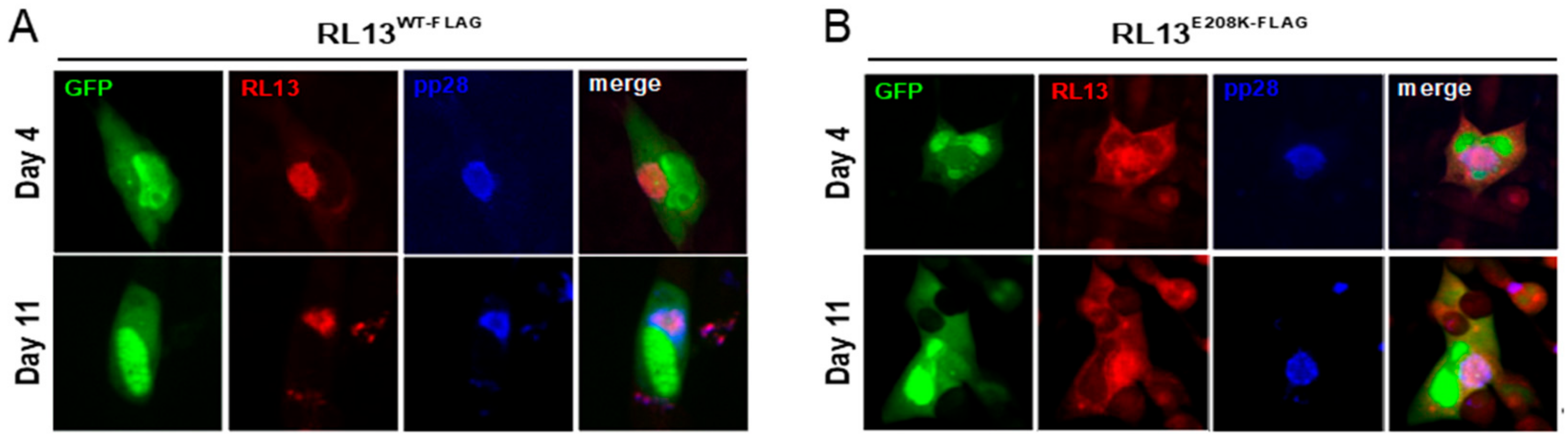
3.4. Detection of RL13WT-FLAG Using Immunofluorescence and Western Blotting
3.5. The E208K Mutation Disrupts RL13 Localization and Function
3.6. Emergence of RL13 Mutations Can Be Quantitated by Immunofluorescence
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional profiling of a human cytomegalovirus genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef] [PubMed]
- Stanton, R.J.; Baluchova, K.; Dargan, D.J.; Cunningham, C.; Sheehy, O.; Seirafian, S.; McSharry, B.P.; Neale, M.L.; Davies, J.A.; Tomasec, P.; et al. Reconstruction of the complete human cytomegalovirus genome in a BAC reveals RL13 to be a potent inhibitor of replication. J. Clin. Investig. 2010, 120, 3191–3208. [Google Scholar] [CrossRef] [PubMed]
- Murrell, I.; Wilkie, G.S.; Davison, A.J.; Statkute, E.; Fielding, C.A.; Tomasec, P.; Wilkinson, G.W.; Stanton, R.J. Genetic Stability of Bacterial Artificial Chromosome-Derived Human Cytomegalovirus during Culture In Vitro. J. Virol. 2016, 90, 3929–3943. [Google Scholar] [CrossRef] [PubMed]
- Dargan, D.J.; Douglas, E.; Cunningham, C.; Jamieson, F.; Stanton, R.J.; Baluchova, K.; McSharry, B.P.; Tomasec, P.; Emery, V.C.; Percivalle, E.; et al. Sequential mutations associated with adaptation of human cytomegalovirus to growth in cell culture. J. Gen. Virol. 2010, 91(6), 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Cortese, M.; Calo, S.; D’Aurizio, R.; Lilja, A.; Pacchiani, N.; Merola, M. Recombinant human cytomegalovirus (HCMV) RL13 binds human immunoglobulin G Fc. PLoS ONE 2012, 7, e50166. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Adler, S.P.; Davison, A.J.; Smith, L.; Habib, S.E.; McVoy, M.A. Bacterial artificial chromosome clones of viruses comprising the towne cytomegalovirus vaccine. J. Biomed. Biotechnol. 2012, 2012, 428498. [Google Scholar] [CrossRef]
- Warming, S.; Costantino, N.; Court, D.L.; Jenkins, N.A.; Copeland, N.G. Simple and highly efficient BAC recombineering using galK selection. Nucleic Acids Res. 2005, 33, e36. [Google Scholar] [CrossRef]
- McVoy, M.A.; Wang, J.B.; Dittmer, D.P.; Bierle, C.J.; Swanson, E.C.; Fernandez-Alarcon, C.; Hernandez-Alvarado, N.; Zabeli, J.C.; Schleiss, M.R. Repair of a Mutation Disrupting the Guinea Pig Cytomegalovirus Pentameric Complex Acquired during Fibroblast Passage Restores Pathogenesis in Immune-Suppressed Guinea Pigs and in the Context of Congenital Infection. J. Virol. 2016, 90, 7715–7727. [Google Scholar] [CrossRef]
- Schleiss, M.R.; Bierle, C.J.; Swanson, E.C.; McVoy, M.A.; Wang, J.B.; Al-Mahdi, Z.; Geballe, A.P. Vaccination with a Live Attenuated Cytomegalovirus Devoid of a Protein Kinase R Inhibitory Gene Results in Reduced Maternal Viremia and Improved Pregnancy Outcome in a Guinea Pig Congenital Infection Model. J. Virol. 2015, 89, 9727–9738. [Google Scholar] [CrossRef]
- Cui, X.; McGregor, A.; Schleiss, M.R.; McVoy, M.A. Cloning the complete guinea pig cytomegalovirus genome as an infectious bacterial artificial chromosome with excisable origin of replication. J. Virol. Methods 2008, 149, 231–239. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Furukawa, T.; Zygraich, N.; Huygelen, C. Candidate cytomegalovirus strain for human vaccination. Infect. Immun. 1975, 12, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Cha, T.A.; Tom, E.; Kemble, G.W.; Duke, G.M.; Mocarski, E.S.; Spaete, R.R. Human cytomegalovirus clinical isolates carry at least 19 genes not found in laboratory strains. J. Virol. 1996, 70, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.; Rose, D.; Wagner, M.; Rhiel, S.; McVoy, M.A. Cloning of the genomes of human cytomegalovirus strains Toledo, TownevarRIT3, and Towne long as BACs and site-directed mutagenesis using a PCR-based technique. Virology 2003, 307, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Prichard, M.N.; Penfold, M.E.; Duke, G.M.; Spaete, R.R.; Kemble, G.W. A review of genetic differences between limited and extensively passaged human cytomegalovirus strains. Rev. Med. Virol. 2001, 11, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Bradley, A.J.; Lurain, N.S.; Ghazal, P.; Trivedi, U.; Cunningham, C.; Baluchova, K.; Gatherer, D.; Wilkinson, G.W.; Dargan, D.J.; Davison, A.J. High-throughput sequence analysis of variants of human cytomegalovirus strains Towne and AD169. J. Gen. Virol. 2009, 90(10), 2375. [Google Scholar] [CrossRef]
- Das, S.; Vasanji, A.; Pellett, P.E. Three-dimensional structure of the human cytomegalovirus cytoplasmic virion assembly complex includes a reoriented secretory apparatus. J. Virol. 2007, 81, 11861–11869. [Google Scholar] [CrossRef]
- Al Qaffas, A.; Camiolo, S.; Nichols, J.; Davison, A.J.; Ourahmane, A.; Cui, X.; Schleiss, M.R.; Hertel, L.; Dittmer, D.P.; McVoy, M.A. Genome Sequence of Human Cytomegalovirus Ig-KG-H2, a Variant of Strain KG Propagated in the Presence of Neutralizing Antibodies. Microbiol. Resour. Announc. 2020, 9, e00063-20. [Google Scholar] [CrossRef]
- Ourahmane, A.; Cui, X.; He, L.; Catron, M.; Dittmer, D.P.; Al Qaffasaa, A.; Schleiss, M.R.; Hertel, L.; McVoy, M.A. Inclusion of Antibodies to Cell Culture Media Preserves the Integrity of Genes Encoding RL13 and the Pentameric Complex Components During Fibroblast Passage of Human Cytomegalovirus. Viruses 2019, 11, 221. [Google Scholar] [CrossRef]
- Al Qaffas, A.; Nichols, J.; Davison, A.J.; Ourahmane, A.; Hertel, L.; McVoy, M.A.; Camiolo, S. LoReTTA, a user-friendly tool for assembling viral genomes from PacBio sequence data. Virus Evol. 2021, 7, veab042. [Google Scholar] [CrossRef]
- Dolan, A.; Cunningham, C.; Hector, R.D.; Hassan-Walker, A.F.; Lee, L.; Addison, C.; Dargan, D.J.; McGeoch, D.J.; Gatherer, D.; Emery, V.C.; et al. Genetic content of wild-type human cytomegalovirus. J. Gen. Virol. 2004, 85(5), 1301–1312. [Google Scholar] [CrossRef]
- Suarez, N.M.; Wilkie, G.S.; Hage, E.; Camiolo, S.; Holton, M.; Hughes, J.; Maabar, M.; Vattipally, S.B.; Dhingra, A.; Gompels, U.A.; et al. Human Cytomegalovirus Genomes Sequenced Directly from Clinical Material: Variation, Multiple-Strain Infection, Recombination, and Gene Loss. J. Infect Dis 2019, 220, 781–791. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modification | Step | Oligonucleotide | Sequence |
|---|---|---|---|
| RL13WT | 1 | RL13 (TL12)-galk-FW | GATATCTGAAGGTTGCAATGACACGTGCTCCTGTCCGTGCAATTGCCTTTACGACTCACTATAGGGCGAATTGG |
| RL13 (TL12)-galk-RV | GCATCGGTGACAGAGTTAGAAGAATTTGTGATAGTGGAGGCGGTGGAGGTGCTATGACCATGATTACGCCAAGC | ||
| 2 | TL12_RL13 (WT)-FWAN | TAACGTTATGGACTGGAGATTTACGGTTAAGTGGACGTTACTGATGATTACGATATCTGAAGGTTGCAATGACACGTGCTCCTGTCCGTGCAATTGCCTT | |
| TL12_RL13 (WT)-RVAN | GTTCCATTTGCGATAGCTGAAGTGCTGTTAGCATCGGTGACAGAGTTAGAAGAATTTGTGATAGTGGAGGCGGTGGAGGTAAGGCAATTGCACGGACAGG | ||
| RL13WT-FLAG | 1 | RL13 (TL12) FLAG-epitope-galk-FW | GTTATACAAAACTTCCCAAATACGACCCAGATGAATTTTAGACTAAAACCACGACTCACTATAGGGCGAATTGG |
| RL13 (TL12) FLAG-epitope-galk-RV | AGACATTATTGGCTAAAAATAAAAACAAAAGTTTATTGATGTGCATGTTAGCTATGACCATGATTACGCCAAGC | ||
| 2 | RL13 (TL12) FLAG-epitope-FWAN | CGTCCCCAGAAGACCGCGTTATACAAAACTTCCCAAATACGACCCAGATGAATTTTAGACTAAAACCGACTACAAGGACGACGATGACAAGGGACCTAAG | |
| RL13 (TL12) FLAG-epitope-RVAN | CCACAAAAACCACACGGAGACATTATTGGCTAAAAATAAAAACAAAAGTTTATTGATGTGCATGTTACTTAGGTCCCTTGTCATCGTCGTCCTTGTAGTC | ||
| RL13E208K | 1 | E208K-GalK-FW | CCGAACACGCTGGAAAATACGTTTTGCAACGTTACAGTGACGGTAAAAAGGACGACTCACTATAGGGCGAATTGG |
| E208K-GalK-RV | GTATAGGAGACGATGTTGCAGTTCCTGATAACACGGTTAAATAGTAGTTTTGCTATGACCATGATTACGCCAAGC | ||
| 2 | E208K-FWAN | ATATACAACGTAACTACCGAACACGCTGGAAAATACGTTTTGCAACGTTACAGTGACGGTAAAAAGAAAAACTAC | |
| E208K-RVAN | TACATTTATCAGGTATAGGAGACGATGTTGCAGTTCCTGATAACACGGTTAAATAGTAGTTTTTCTTTTTACCGT |
| Passage | ||||
|---|---|---|---|---|
| Lineage | 0 | 2 | 4 | 10 |
| RL13-WT-FLAG-1 | 0/9 a | 2/10 a | 7/9 a | 9/9 a |
| RL13-WT-FLAG-2 | 0/10 b | 2/10 b | 9/10 b | 10/10 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ourahmane, A.; Hertel, L.; McVoy, M.A. The RL13 Temperance Factor Represses Replication of the Highly Cell Culture-Adapted Towne Strain of Human Cytomegalovirus. Viruses 2023, 15, 1023. https://doi.org/10.3390/v15041023
Ourahmane A, Hertel L, McVoy MA. The RL13 Temperance Factor Represses Replication of the Highly Cell Culture-Adapted Towne Strain of Human Cytomegalovirus. Viruses. 2023; 15(4):1023. https://doi.org/10.3390/v15041023
Chicago/Turabian StyleOurahmane, Amine, Laura Hertel, and Michael A. McVoy. 2023. "The RL13 Temperance Factor Represses Replication of the Highly Cell Culture-Adapted Towne Strain of Human Cytomegalovirus" Viruses 15, no. 4: 1023. https://doi.org/10.3390/v15041023