
Human Cytomegalovirus UL23 Antagonizes the Antiviral Effect of Interferon-γ by Restraining the Expression of Specific IFN-Stimulated Genes
 and
and 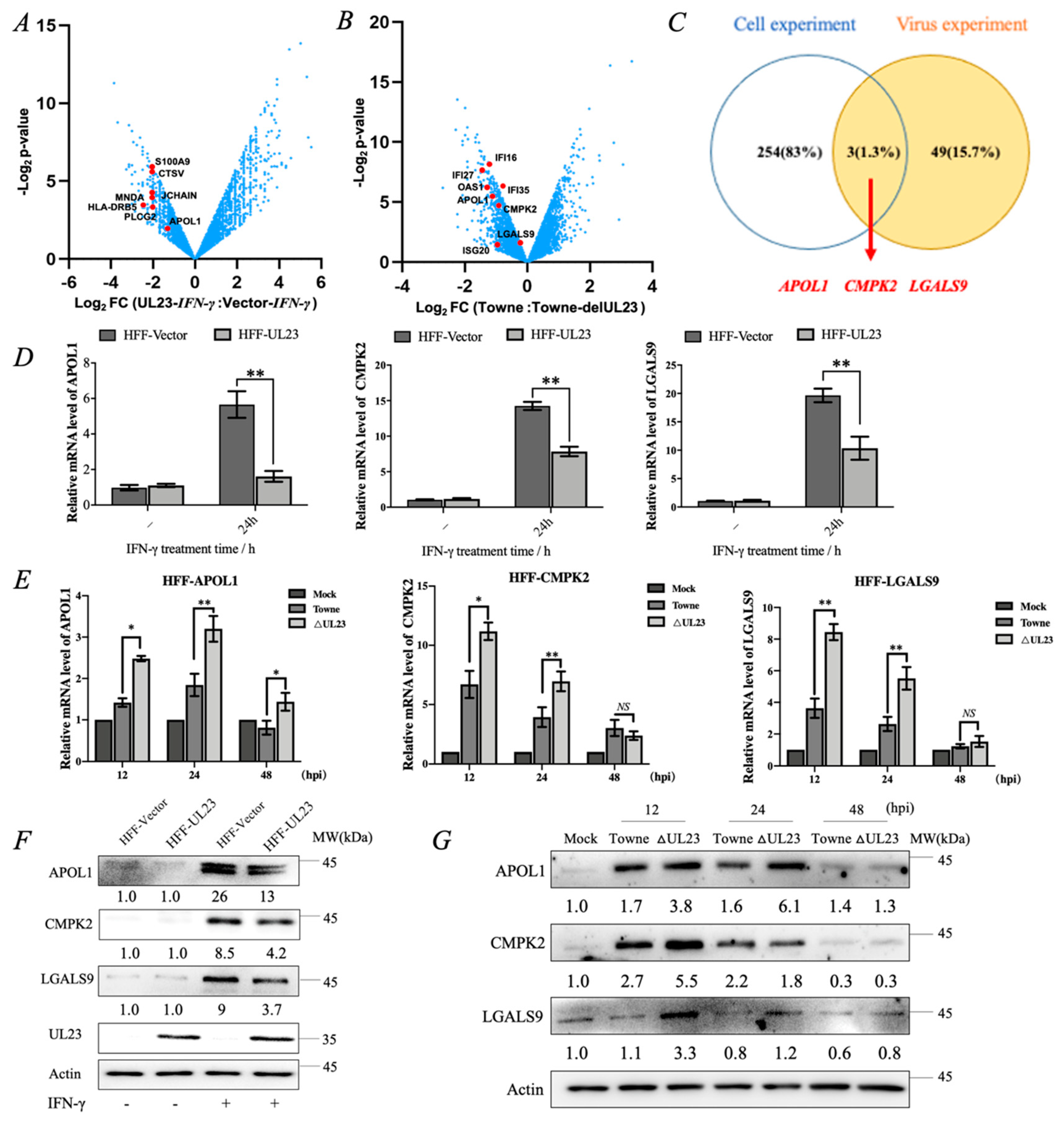{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
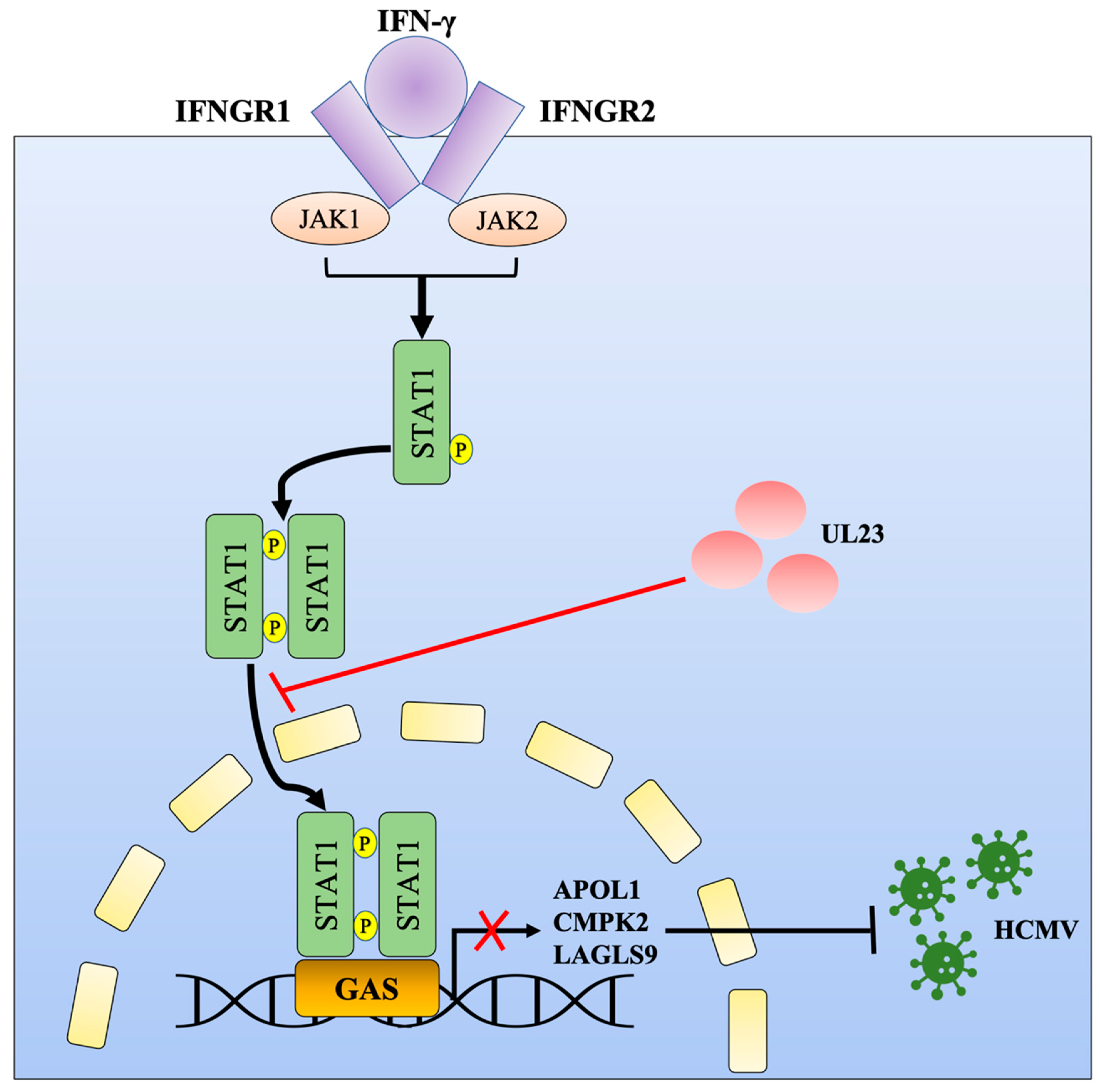
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cell Culture
2.2. Reagents and Antibodies
2.3. RNA-Seq and Data Analysis
2.4. Virus Infection and Growth Analysis
2.5. RNA or DNA Isolation and Quantitative Reverse Transcription-PCR (RT-qPCR)
2.6. Western Blotting Assay
2.7. RNA Interference
2.8. Stable Cell Lines
2.9. Statistical Analysis
3. Results
3.1. RNA-Seq Analysis Revealed That HCMV-Encoded UL23 Protein Inhibited the Expression of IFN-Inducible Genes APOL1, CMPK2, and LGALS9
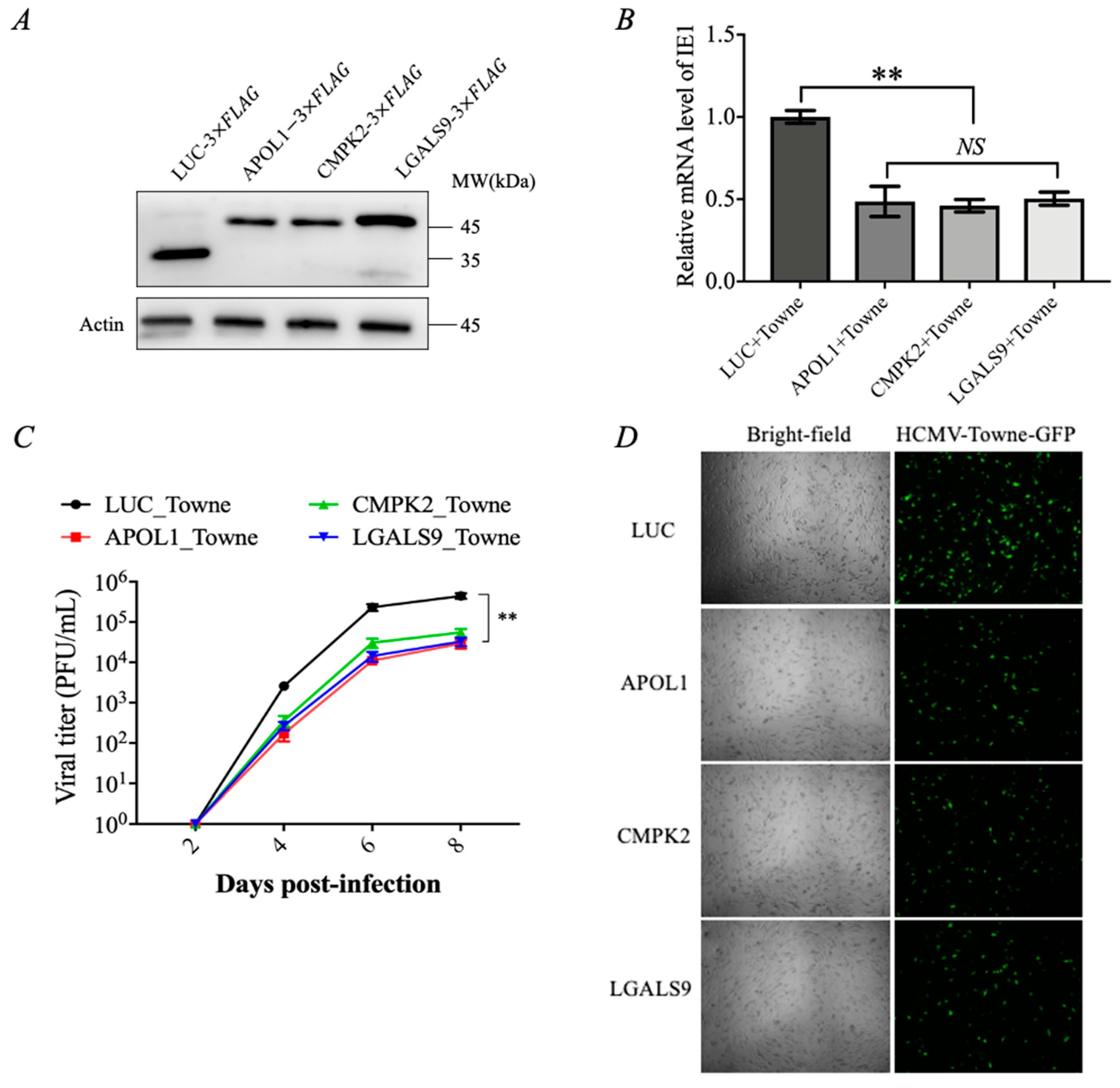
3.2. IFN-γ-Inducible Genes APOL1, CMPK2, and LGALS9 Inhibit HCMV Replication
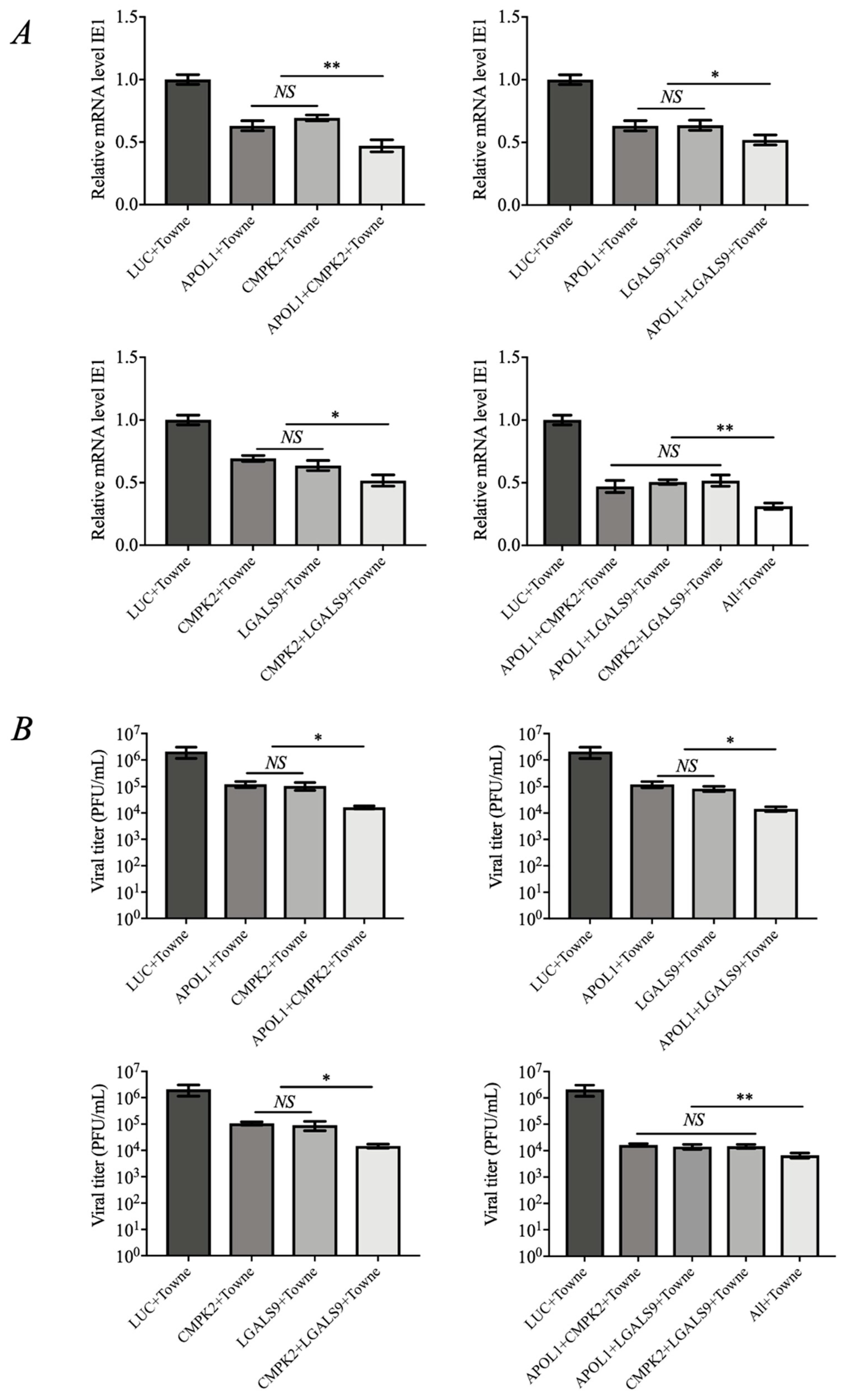
3.3. The Combination of APOL1, CMPK2, and LGALS9 Exhibits Synergistic Anti-HCMV Effects
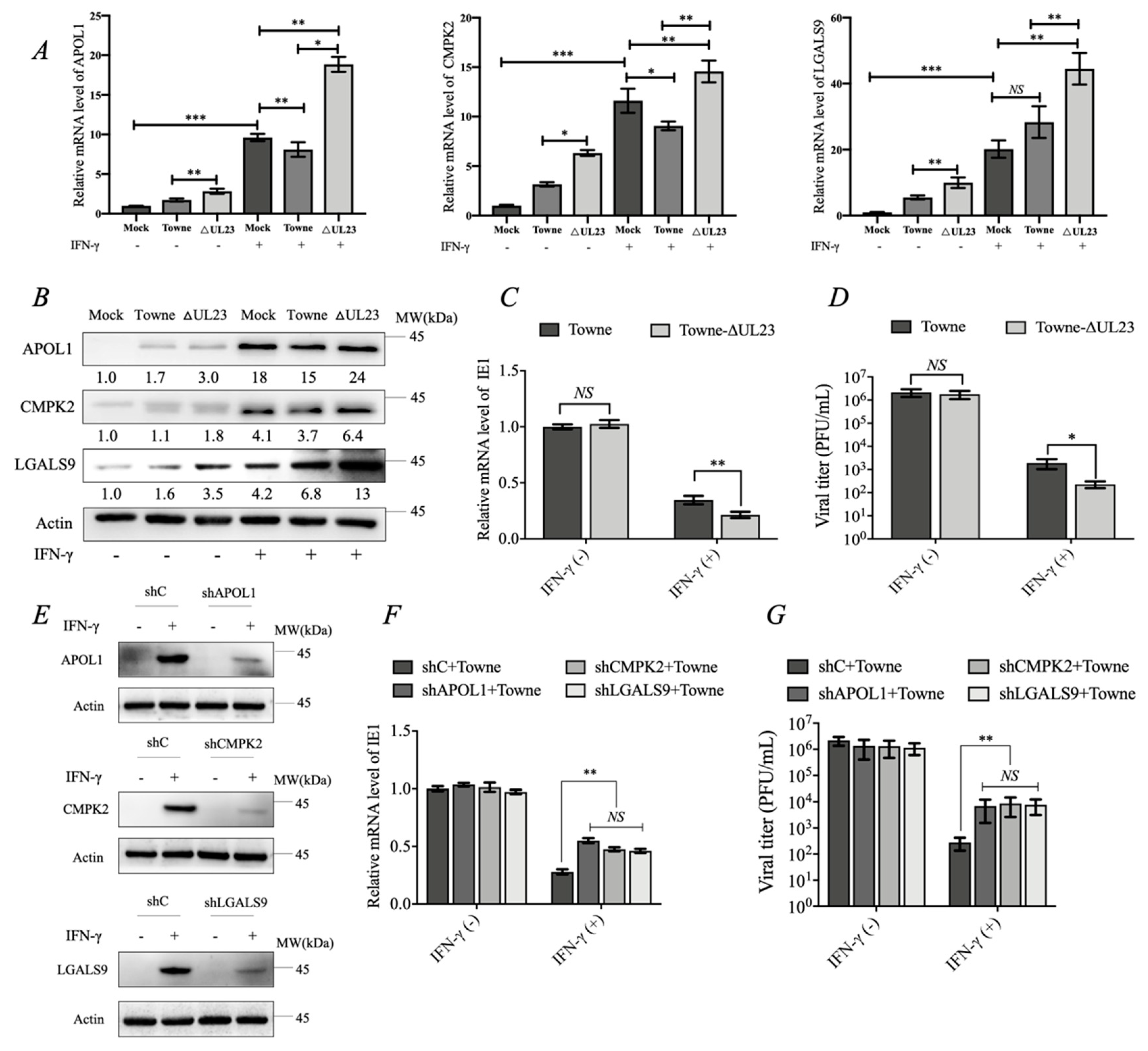
3.4. UL23 Limited the Antiviral Effect of IFN-γ by Downregulating the Expression of APOL1, CMPK2, and LGALS9
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crough, T.; Khanna, R. Immunobiology of human cytomegalovirus: From bench to bedside. Clin. Microbiol. Rev. 2009, 22, 76–98. [Google Scholar] [CrossRef]
- Yasuoka, K.; Fujikawa, H.; Shibazaki, S. Cytomegalovirus enteritis: A surprising cause of severe diarrhoea and protein-losing gastroenteropathy in an intensive care patient. BMJ Case Rep. 2022, 15, e249258. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A.P.; Kirby, K.A.; Rubenfeld, G.D.; Leisenring, W.M.; Bulger, E.M.; Neff, M.J.; Gibran, N.S.; Huang, M.L.; Santo Hayes, T.K.; Corey, L.; et al. Cytomegalovirus reactivation in critically ill immunocompetent patients. JAMA 2008, 300, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Sweet, C. The pathogenicity of cytomegalovirus. FEMS Microbiol. Rev. 1999, 23, 457–482. [Google Scholar] [CrossRef]
- Noriega, V.; Redmann, V.; Gardner, T.; Tortorella, D. Diverse immune evasion strategies by human cytomegalovirus. Immunol. Res. 2012, 54, 140–151. [Google Scholar] [CrossRef]
- Jarvis, M.A.; Nelson, J.A. Mechanisms of human cytomegalovirus persistence and latency. Front. Biosci. 2002, 7, d1575–d1582. [Google Scholar] [CrossRef]
- Goodrum, F.; Caviness, K.; Zagallo, P. Human cytomegalovirus persistence. Cell Microbiol. 2012, 14, 644–655. [Google Scholar] [CrossRef]
- Trilling, M.; Le, V.T.; Hengel, H. Interplay between CMVs and interferon signaling: Implications for pathogenesis and therapeutic intervention. Future Microbiol. 2012, 7, 1269–1282. [Google Scholar] [CrossRef]
- Goodwin, C.M.; Ciesla, J.H.; Munger, J. Who’s Driving? Human Cytomegalovirus, Interferon, and NFkappaB Signaling. Viruses 2018, 10, 447. [Google Scholar] [CrossRef] [PubMed]
- Browne, E.P.; Shenk, T. Human cytomegalovirus UL83-coded pp65 virion protein inhibits antiviral gene expression in infected cells. Proc. Natl. Acad. Sci. USA 2003, 100, 11439–11444. [Google Scholar] [CrossRef]
- Kim, Y.E.; Ahn, J.H. Positive role of promyelocytic leukemia protein in type I interferon response and its regulation by human cytomegalovirus. PLoS Pathog. 2015, 11, e1004785. [Google Scholar] [CrossRef] [PubMed]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Stark, G.R. How cells respond to interferons revisited: From early history to current complexity. Cytokine Growth Factor Rev. 2007, 18, 419–423. [Google Scholar] [CrossRef]
- Der, S.D.; Zhou, A.; Williams, B.R.; Silverman, R.H. Identification of genes differentially regulated by interferon alpha, beta, or gamma using oligonucleotide arrays. Proc. Natl. Acad. Sci. USA 1998, 95, 15623–15628. [Google Scholar] [CrossRef]
- Dell’Oste, V.; Biolatti, M.; Galitska, G.; Griffante, G.; Gugliesi, F.; Pasquero, S.; Zingoni, A.; Cerboni, C.; De Andrea, M. Tuning the Orchestra: HCMV vs. Innate Immunity. Front. Microbiol. 2020, 11, 661. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, J.; Cristea, I.M. Human cytomegalovirus tegument protein pUL83 inhibits IFI16-mediated DNA sensing for immune evasion. Cell Host Microbe 2013, 14, 591–599. [Google Scholar] [CrossRef]
- Fu, Y.Z.; Guo, Y.; Zou, H.M.; Su, S.; Wang, S.Y.; Yang, Q.; Luo, M.H.; Wang, Y.Y. Human cytomegalovirus protein UL42 antagonizes cGAS/MITA-mediated innate antiviral response. PLoS Pathog. 2019, 15, e1007691. [Google Scholar] [CrossRef]
- Choi, H.J.; Park, A.; Kang, S.; Lee, E.; Lee, T.A.; Ra, E.A.; Lee, J.; Lee, S.; Park, B. Human cytomegalovirus-encoded US9 targets MAVS and STING signaling to evade type I interferon immune responses. Nat. Commun. 2018, 9, 125. [Google Scholar] [CrossRef]
- Park, A.; Ra, E.A.; Lee, T.A.; Choi, H.J.; Lee, E.; Kang, S.; Seo, J.Y.; Lee, S.; Park, B. HCMV-encoded US7 and US8 act as antagonists of innate immunity by distinctively targeting TLR-signaling pathways. Nat. Commun. 2019, 10, 4670. [Google Scholar] [CrossRef]
- Huang, Z.F.; Zou, H.M.; Liao, B.W.; Zhang, H.Y.; Yang, Y.; Fu, Y.Z.; Wang, S.Y.; Luo, M.H.; Wang, Y.Y. Human Cytomegalovirus Protein UL31 Inhibits DNA Sensing of cGAS to Mediate Immune Evasion. Cell Host Microbe 2018, 24, 69–80.e4. [Google Scholar] [CrossRef]
- Chen, J.; Xia, S.; Yang, X.; Chen, H.; Li, F.; Liu, F.; Chen, Z. Human Cytomegalovirus Encoded miR-US25-1-5p Attenuates CD147/EMMPRIN-Mediated Early Antiviral Response. Viruses 2017, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Sheng, J.; Vu, G.P.; Liu, Y.; Foo, C.; Wu, S.; Trang, P.; Paliza-Carre, M.; Ran, Y.; Yang, X.; et al. Human cytomegalovirus UL23 inhibits transcription of interferon-gamma stimulated genes and blocks antiviral interferon-gamma responses by interacting with human N-myc interactor protein. PLoS Pathog. 2018, 14, e1006867. [Google Scholar] [CrossRef]
- Feng, L.; Li, W.; Wu, X.; Li, X.; Yang, X.; Ran, Y.; Wu, J.; Li, H. Human Cytomegalovirus UL23 Attenuates Signal Transducer and Activator of Transcription 1 Phosphorylation and Type I Interferon Response. Front. Microbiol. 2021, 12, 692515. [Google Scholar] [CrossRef]
- Marchini, A.; Liu, H.; Zhu, H. Human cytomegalovirus with IE-2 (UL122) deleted fails to express early lytic genes. J. Virol. 2001, 75, 1870–1878. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional profiling of a human cytomegalovirus genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef]
- Boldogh, I.; Bui, T.K.; Szaniszlo, P.; Bresnahan, W.A.; Albrecht, T.; Hughes, T.K. Novel activation of gamma-interferon in nonimmune cells during human cytomegalovirus replication. Proc. Soc. Exp. Biol. Med. 1997, 215, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.E.; Khatua, A.K.; Popik, W. The innate immune factor apolipoprotein L1 restricts HIV-1 infection. J. Virol. 2014, 88, 592–603. [Google Scholar] [CrossRef]
- El-Diwany, R.; Soliman, M.; Sugawara, S.; Breitwieser, F.; Skaist, A.; Coggiano, C.; Sangal, N.; Chattergoon, M.; Bailey, J.R.; Siliciano, R.F.; et al. CMPK2 and BCL-G are associated with type 1 interferon-induced HIV restriction in humans. Sci. Adv. 2018, 4, eaat0843. [Google Scholar] [CrossRef]
- Miyakawa, K.; Nishi, M.; Ogawa, M.; Matsunaga, S.; Sugiyama, M.; Nishitsuji, H.; Kimura, H.; Ohnishi, M.; Watashi, K.; Shimotohno, K.; et al. Galectin-9 restricts hepatitis B virus replication via p62/SQSTM1-mediated selective autophagy of viral core proteins. Nat. Commun. 2022, 13, 531. [Google Scholar] [CrossRef] [PubMed]
- Le, V.T.K.; Trilling, M.; Wilborn, M.; Hengel, H.; Zimmermann, A. Human cytomegalovirus interferes with signal transducer and activator of transcription (STAT) 2 protein stability and tyrosine phosphorylation. J. Gen. Virol. 2008, 89 Pt 10, 2416–2426. [Google Scholar] [CrossRef]
- Biolatti, M.; Dell’Oste, V.; Pautasso, S.; Gugliesi, F.; von Einem, J.; Krapp, C.; Jakobsen, M.R.; Borgogna, C.; Gariglio, M.; De Andrea, M.; et al. Human Cytomegalovirus Tegument Protein pp65 (pUL83) Dampens Type I Interferon Production by Inactivating the DNA Sensor cGAS without Affecting STING. J. Virol. 2018, 92, e01774-17. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Amsler, L.; Verweij, M.; DeFilippis, V.R. The tiers and dimensions of evasion of the type I interferon response by human cytomegalovirus. J. Mol. Biol. 2013, 425, 4857–4871. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, A.C.; Stempel, M.; Chan, B.; Brinkmann, M.M. One Step Ahead: Herpesviruses Light the Way to Understanding Interferon-Stimulated Genes (ISGs). Front. Microbiol. 2020, 11, 124. [Google Scholar] [CrossRef]
- Platanitis, E.; Demiroz, D.; Schneller, A.; Fischer, K.; Capelle, C.; Hartl, M.; Gossenreiter, T.; Muller, M.; Novatchkova, M.; Decker, T. A molecular switch from STAT2-IRF9 to ISGF3 underlies interferon-induced gene transcription. Nat. Commun. 2019, 10, 2921. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Lai, J.H.; Wu, D.W.; Wu, C.H.; Hung, L.F.; Huang, C.Y.; Ka, S.M.; Chen, A.; Chang, Z.F.; Ho, L.J. Mitochondrial CMPK2 mediates immunomodulatory and antiviral activities through IFN-dependent and IFN-independent pathways. iScience 2021, 24, 102498. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, K.; Jeremiah, S.S.; Ogawa, M.; Nishi, M.; Ohnishi, M.; Ryo, A. Crosstalk between the innate immune system and selective autophagy in hepatitis B virus infection. Autophagy 2022, 18, 2006–2007. [Google Scholar] [CrossRef]
- Moar, P.; Tandon, R. Galectin-9 as a biomarker of disease severity. Cell Immunol. 2021, 361, 104287. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Subbannayya, Y.; Humphries, F.; Skejsol, A.; Pinto, S.M.; Giambelluca, M.; Espevik, T.; Fitzgerald, K.A.; Kandasamy, R.K. UMP-CMP kinase 2 gene expression in macrophages is dependent on the IRF3-IFNAR signaling axis. PLoS ONE 2021, 16, e0258989. [Google Scholar] [CrossRef] [PubMed]
- Muehlig, A.K.; Gies, S.; Huber, T.B.; Braun, F. Collapsing Focal Segmental Glomerulosclerosis in Viral Infections. Front. Immunol. 2021, 12, 800074. [Google Scholar] [CrossRef] [PubMed]
- Machala, E.A.; Avdic, S.; Stern, L.; Zajonc, D.M.; Benedict, C.A.; Blyth, E.; Gottlieb, D.J.; Abendroth, A.; McSharry, B.P.; Slobedman, B. Restriction of Human Cytomegalovirus Infection by Galectin-9. J. Virol. 2019, 93, e01746-18. [Google Scholar] [CrossRef] [PubMed]
- Hubel, P.; Urban, C.; Bergant, V.; Schneider, W.M.; Knauer, B.; Stukalov, A.; Scaturro, P.; Mann, A.; Brunotte, L.; Hoffmann, H.H.; et al. A protein-interaction network of interferon-stimulated genes extends the innate immune system landscape. Nat. Immunol. 2019, 20, 493–502. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Peng, W.; Wang, J.; Zhang, C.; Zhao, W.; Ran, Y.; Yang, X.; Chen, J.; Li, H. Human Cytomegalovirus UL23 Antagonizes the Antiviral Effect of Interferon-γ by Restraining the Expression of Specific IFN-Stimulated Genes. Viruses 2023, 15, 1014. https://doi.org/10.3390/v15041014
Wang H, Peng W, Wang J, Zhang C, Zhao W, Ran Y, Yang X, Chen J, Li H. Human Cytomegalovirus UL23 Antagonizes the Antiviral Effect of Interferon-γ by Restraining the Expression of Specific IFN-Stimulated Genes. Viruses. 2023; 15(4):1014. https://doi.org/10.3390/v15041014
Chicago/Turabian StyleWang, Hankun, Weijian Peng, Jialin Wang, Chunling Zhang, Wangchun Zhao, Yanhong Ran, Xiaoping Yang, Jun Chen, and Hongjian Li. 2023. "Human Cytomegalovirus UL23 Antagonizes the Antiviral Effect of Interferon-γ by Restraining the Expression of Specific IFN-Stimulated Genes" Viruses 15, no. 4: 1014. https://doi.org/10.3390/v15041014