
The Role of the Leishmania infantum Infected Dogs as a Potential Reservoir Host for Toscana Virus in a Zoonotic Visceral Leishmaniasis Focus of Northern Tunisia

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Protocol
2.2. Seasonal Activity of Sandflies in the Exposition Site
2.3. Collection, Identification, and Infection Prevalence of Wild Sandflies with L. infantum and TOSV in the Exposure Site
2.4. Ethics Statement
2.5. Xenodiagnosis
2.5.1. Detection of L. infantum Infections in P. perniciosus Females Fed on Dogs
2.5.2. Detection of TOSV in P. perniciosus Fed on Dogs
2.5.3. Isolation of Phlebovirus from Infected P. perniciosus with TOSV following Engorgement on Dogs
2.6. Phylogenetic Analysis
3. Results
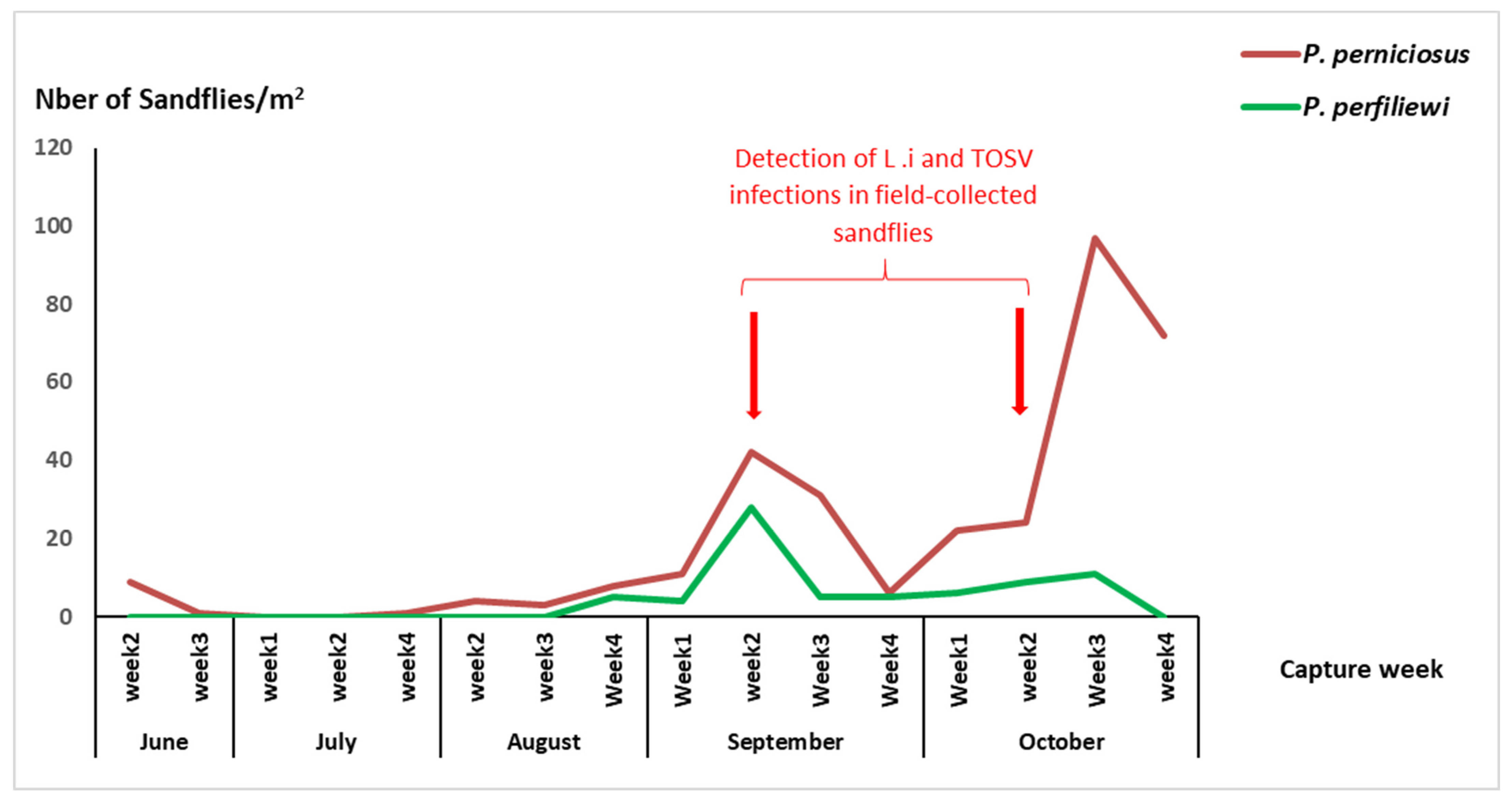
3.1. Abundance and Seasonal Activity of Sandflies in the Exposure Site
3.2. Detection of Leishmania infantum and TOSV in Field-Collected Sandflies from the Exposure Site
3.3. Infection of Phlebotomus perniciosus with Leishmania infantum by Xenodiagnosis
3.4. Infection of Phlebotomus perniciosus with Toscana Virus by Xenodiagnosis
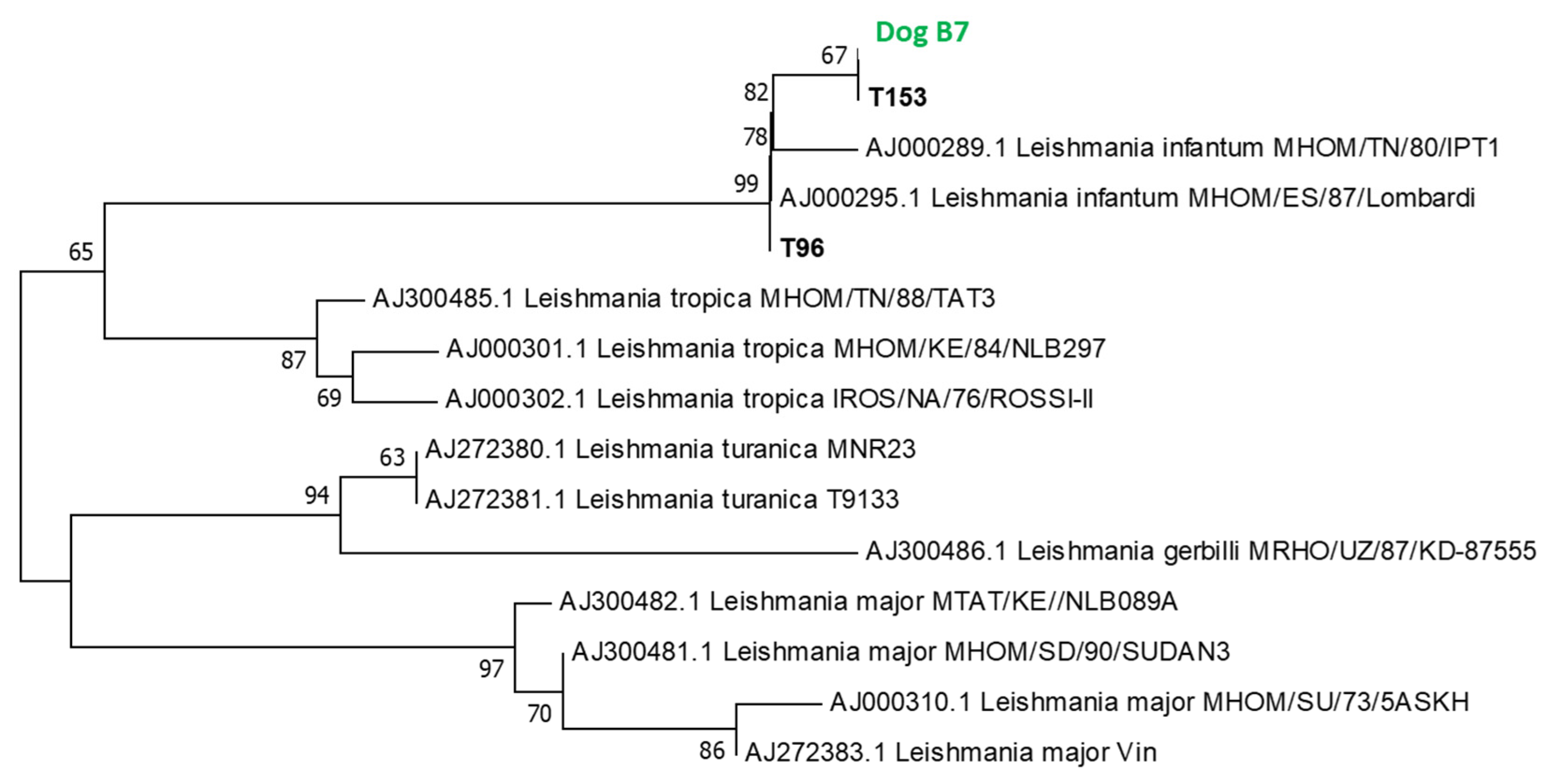
3.5. Phylogenetic Study
3.5.1. Phylogenetic Analysis of L. infantum
3.5.2. Homology, Genetic Distances and Phylogenetic Analysis of TOSV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chelbi, I.; Maghraoui, K.; Zhioua, S.; Cherni, S.; Labidi, I.; Satoskar, A.; Hamilton, J.G.C.; Zhioua, E. Enhanced attractiveness of sandfly vectors of Leishmania infantum to dogs infected with zoonotic visceral leishmaniasis. PLoS Negl. Trop. Dis. 2021, 15, e0009647. [Google Scholar]
- Ben Ismail, R. Incrimination de Phlebotomus perniciosus comme vecteur de Leishmania infantum. Arch. Inst. Past. Tunis 1993, 70, 91–110. [Google Scholar]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.; Amaducci, L.; Coluzzi, M.; et al. Ecology of viruses isolated from sandflies in Italy and characterized of a new Phlebovirus (Arabia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sanchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef]
- Bichaud, L.; Souris, M.; Mary, C.; Ninove, L.; Thirion, L.; Piarroux, R.P.; Piarroux, R.; de Lamballerie, X.; Charrel, R.N. Epidemiologic relationship between Toscana virus infection and Leishmania infnatum due to common exposure to Phlebotomus perniciosus sandfly vector. PLoS Negl. Trop. Dis. 2011, 5, e1328. [Google Scholar] [CrossRef]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Am. J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef]
- Bahri, O.; Fazaa, O.; Ben Alaya-Bouafif, N.; Bouloy, M.; Triki, H.; Bouattour, A. Role of Toscana virus in meningo-encephalitis in Tunisia. Pathol. Biol. 2011, 59, 125–127. [Google Scholar]
- Sakhria, S.; Bichaud, L.; Mensi, M.; Salez, N.; Dachraoui, K.; Thirion, L.; Cherni, S.; Chelbi, I.; de Lamballerie, X.; Zhioua, E.; et al. Co-Circulation of Toscana Virus and Punique Virus in Northern Tunisia: A microneutralisation-based seroprevalence study. PLoS Negl. Trop. Dis. 2013, 7, e2429. [Google Scholar] [CrossRef]
- Sakhria, S.; Alwassouf, S.; Fares, W.; Bichaud, L.; Dachraoui, K.; Alkan, C.; Zoghlami, Z.; de Lamballerie, X.; Zhioua, E.; Charrel, R.N. Presence of sand-fly phlebovirus of two antigenic complexes (Sandfly fever Naples virus and Sandfly fever Sicilian virus) in two different bio-geographical regions of Tunisia demonstrated by a microneutralization-based sero-prevalence study in dogs. Parasit. Vectors 2014, 7, 476. [Google Scholar]
- Zoghlami, Z.; Chouihi, E.; Barhoumi, W.; Dachraoui, K.; Massoudi, N.; Ben Hele, K.; Habboul, Z.; Hadhri, M.H.; Limam, S.; Mhadhbi, M.; et al. Interactions between canine and human visceral leishmaniases in a holoendemic focus of Central Tunisia. Acta Trop. 2014, 139, 32–38. [Google Scholar] [CrossRef]
- Dachraoui, K.; Fares, W.; Bichaud, L.; Barhoumi, W.; Beier, J.C.; Derbali, M.; Cherni, S.; de Lamballerie, X.; Chelbi, I.; Charrel, R.N.; et al. Phleboviruses associated with sandflies in arid bio-geographical areas of Central Tunisia. Acta Trop. 2016, 158, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Aoun, K.; Diwani, M.F.; Benikhlef, R.; Bouratbine, A.; Ben Haj Ali, S.; Harrat, Z.; Benlkaïd, M.; Kilani, M.; Ben Ismail, R. Leishmania infantum MON-1: Seul zymodème isolé chez les chiens leishmaniens en Tunisie. Bull. Soc. Pathol. Exot. 2003, 96, 77–79. [Google Scholar]
- Ben Slimane, T.; Chouihi, E.; Ben Hadj Ahmed, S.; Chelbi, I.; Barhoumi, W.; Cherni, S.; Zoghlami, Z.; Gharbi, M.; Zhioua, E. An investigation on vertical transmission of Leishmania infantum in experimentally infected dogs and assessment of offspring’s infectiousness potential by xenodiagnosis. Vet. Parasitol. 2014, 15, 282–286. [Google Scholar] [CrossRef]
- Quinnell, R.J.; Courtenay, O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology 2009, 136, 1915–1934. [Google Scholar] [CrossRef]
- Fares, W.; Dachraoui, K.; Barhoumi, W.; Cherni, S.; Chelbi, I.; Zhioua, E. Co-circulation of Toscana virus and Leishmania infantum in a focus of zoonotic visceral leishmaniasis from Central Tunisia. Acta Trop. 2020, 204, 105342. [Google Scholar] [CrossRef] [PubMed]
- Zhioua, E.; Kaabi, B.; Chelbi, I. Entomological investigations following the spread of visceral leishmaniasis toward the center and the south of Tunisia. J. Vector Ecol. 2007, 32, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Chelbi, I.; Derbali, M.; Al-Ahmadi, Z.; Zaafouri, B.; El Fahem, A.; Zhioua, E. Phenology of Phlebotomus papatasi (Diptera: Psychodidae) relative to the seasonal prevalence of zoonotic cutaneous leishmaniasis in central Tunisia. J. Med. Entomol. 2007, 44, 385–388. [Google Scholar] [CrossRef]
- Dachraoui, K.; Chelbi, I.; Ben Said, M.; Ben Osman, R.; Cherni, S.; Charrel, R.; Zhioua, E. Transmission dynamic of Punique virus in Tunisia. Viruses 2022, 14, 904. [Google Scholar] [CrossRef]
- Croset, H.; Rioux, J.A.; Master, M.; Bayar, N. Les phlébotomes de la Tunisie (Diptera, Phlebotominae). Mise au point systématique, chorologique et éthologique. Ann. Parasitol. Hum. Comp. 1978, 53, 711–749. [Google Scholar] [CrossRef]
- Leger, N.; Pesson, B.; Madulo-Leblond, G.; Abonnenc, E. Sur la différenciation des femelles du sous-genre Larroussius, Nitzulescu, 1931 (Diptera-Phlebotominae) de la région méditerrannéenne. Ann. Parasitol. Hum. Comp. 1983, 58, 611–623. [Google Scholar] [CrossRef]
- Pesson, B.; Ready, J.S.; Benabdennbi, I.; Martín-Sánchez, J.; Esseghir, S.; Cadi-Soussi, M.; Morillas-Marquez, F.; Ready, P.D. Sandflies of the Phlebotomus perniciosus complex: Mitochondrial introgression and a new sibling species of Phlebotomus longicuspis in the Moroccan Rif. Med. Vet. Entomol. 2004, 18, 25–37. [Google Scholar] [CrossRef]
- Boussaa, S.; Boumezzough, A.; Remy, P.E.; Glasser, N.; Pesson, B. Morphological and isoenzymatic differentitation of Phlebotomus pernicious and Phlebotomus longicuspis (Diptera: Psychodidae) in Southern Morocco. Acta Trop. 2008, 106, 184–189. [Google Scholar] [CrossRef]
- Parvizi, P.; Mauricio, I.; Aransay, A.M.; Miles, M.A.; Ready, P.D. First detection of Leishmania major in peridomestic Phlebotomus papatasi from Isfahan province, Iran: Comparison of nested PCR of nuclear ITS ribosomal DNA and semi-nested PCR of minicircle kinetoplast DNA. Acta Trop. 2005, 93, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Ready, P.D. Nested PCRs and sequencing of nuclear ITS-rDNA fragments detect three Leishmania species of gerbils in sandflies from Iranian foci of zoonotic cutaneous leishamniasis. Trop. Med. Int. Health 2008, 13, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Moradi, G.; Akbari, G.; Farhmand, M.; Ready, P.; Piazak, N.; Assmar, M.; Amirkhani, A. PCR detection and sequencing of parasite ITS-rDNA gene from reservoir host of zoonotic cutaneous leishmaniasis in central Iran. Parasitol. Res. 2008, 103, 1273–1278. [Google Scholar] [CrossRef]
- Sanchez-Seco, M.P.; Echevarria, J.M.; Hernandez, L.; Estevez, D.; Navarro-Mari, J.M.; Tenorio, A. Detection and identification of Toscana and other phleboviruses by RT-nested-PCR assays 453 with degenerated primers. J. Med. Virol. 2003, 71, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Zhioua, E.; Moureau, G.; Chelbi, I.; Ninove, L.; Salez, N.; Derbali, M.; Champs, M.; Cherni, S.; Lamballerie, X.; Charrel, R.N. Punique virus, a novel phlebovirus, related to Sandfly fever Naples virus, isolated from sandflies collected in Tunisia. J. Gen. Virol. 2010, 91, 1275–1283. [Google Scholar] [CrossRef]
- Maroli, M.; Fiorentino, S.; Guandalini, E. Biology of a laboratory colony of Phlebotomus perniciosus (Diptera: Psychodidae). J. Med. Entomol. 1987, 24, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Chelbi, I.; Zhioua, E. Biology of Phlebotomus papatasi Scopoli (Diptera: Psychodidae) in the laboratory. J. Med. Entomol. 2007, 44, 597–600. [Google Scholar] [CrossRef]
- Courtenay, O.; Carson, C.; Calvo-Bado, L.; Garcez, L.M.; Quinnell, R.J. Heterogeneities in Leishmania infantum infection: Using skin parasite burdens to identify highly infectious dogs. PLoS Negl. Trop. Dis. 2014, 8, e2583. [Google Scholar] [CrossRef] [PubMed]
- Chengfeng, L.; Jian, Y.; Jia, H.; Xiulian, S. On the calculation of TCID50 for quantitation of virus infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetic Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Piorkowski, G.; Chelbi, I.; Moureau, G.; Cherni, S.; de Lamballerie, X.; Sakhria, S.; Charrel, R.N.; Zhioua, E. Isolation of Toscana virus from sandfliessandflies, Tunisia. Emerg. Infect. Dis. 2013, 19, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, W.; Fares, W.; Cherni, S.; Derbali, M.; Dachraoui, K.; Chelbi, I.; Ramalho-Ortigao, M.; Beier, J.C.; Zhioua, E. Changes of sandfly populations and Leishmania infantum infection rates in an irrigated village located in arid Central Tunisia. Int. J. Environ. Res. Public Health 2016, 13, 329. [Google Scholar] [CrossRef]
- Remoli, M.E.; Jiménez, M.; Fortuna, C.; Benedetti, E.; Marchi, A.; Genovese, D.; Gramiccia, M.; Molina, R.; Ciufolini, M.G. Phleboviruses detection in Phlebotomus perniciosus from a human leishmaniasis focus in South-West Madrid region, Spain. Parasit. Vectors 2016, 9, 205. [Google Scholar] [CrossRef]
- Faucher, B.; Bichaud, L.; Charrel, R.; Mary, C.; Izri, A.; de Lamballerie, X.; Piarroux, R. Presence of sandflies infected with Leishmania infantum and Massilia virus in the Marseille urban area. Clin. Microbiol. Infect. 2014, 20, 340–343. [Google Scholar] [CrossRef]
- Es-Sette, N.; Ajouad, M.; Bichaud, L.; Hamdi, S.; Mellouki, F.; Charrel, R.N.; Lemrani, M. Phlebotomus sergenti a common vector of Leishmania tropica and Toscana virus in Morocco. J. Vector Borne Dis. 2014, 51, 86–90. [Google Scholar]
- Ergunay, K.; Kasap, O.E.; Orsten, S.; Oter, K.; Gunay, F.; Yoldar, A.Z.A.; Dincer, E.; Alten, B.; Ozkul, A. Phlebovirus and Leishmania detection in sandflies from eastern Thrace and northern Cyprus. Parasit. Vectors 2014, 7, 575. [Google Scholar] [CrossRef]
- Tesh, R.B.; Modi, G.B. Maintenance of Toscana virus in Phlebotomus perniciosus by vertical transmission. Am. J. Trop. Med. Hyg. 1987, 36, 189–193. [Google Scholar] [CrossRef]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae: Phlebovirus) in its natural sandfly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef]
- Maroli, M.; Ciufolini, M.G.; Verani, P. Vertical transmission of Toscana virus in the sandfly, Phlebotomus perniciosus, via the second gonotrophic cycle. Med. Vet. Entomol. 1993, 7, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; López-Roig, M.; Monastiri, A.; Charrel, R.N.; Serra-Cobo, J. Seroprevalence of Toscana Virus and Sandfly Fever Sicilian Virus in European bat colonies measured using a neutralization test. Viruses 2021, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Ayhan, N.; Ortuño, M.; Ortiz, J.; Gould, E.A.; Maia, C.; Berriatua, E.; Charrel, R.N. Experimental infection of dogs with Toscana Virus and Sandfly Fever Sicilian Virus to determine their potential as possible vertebrate hosts. Microorganisms 2020, 8, 596. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Duboise, S.M. Viremia and immune response with sequential Phlebovirus infections. Am. J. Trop. Med. Hyg. 1987, 36, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Magalhães-Junior, J.T.; Mota, T.F.; Porfirio-Passos, G.; Larangeira, D.F.; Franke, C.R.; Barrouin-Melo, S.M. Xenodiagnosis on dogs with visceral leishmaniasis: Canine and sandfly aspects related to the parasite transmission. Vet. Parasitol. 2017, 223, 120–126. [Google Scholar] [CrossRef]
- Ben Helel, K.; Ben Rejeb, M.; Habboul, Z.; Khattat, N.; Mejaouel, H.; Said-Latiri, H.; Kaabi, B.; Zhioua, E. Risk factors for mortality of children with zoonotic visceral leishmaniasis in Central Tunisia. PLoS ONE 2017, 12, e0189725. [Google Scholar] [CrossRef]
- Alvar, J.; Aparicio, P.; Aseffa, A.; Den Boer, M.; Cañavate, C.; Dedet, J.P.; Gradoni, L.; ter Horst, R.; López-Vélez, R.; Moreno, J. The relationship between leishaniasis and AIDS: The second 10 years. Clin. Microbiol. Rev. 2008, 21, 334–359. [Google Scholar] [CrossRef]
- Rossi, M.; Castiglioni, P.; Hartley, M.A.; Eren, R.O.; Prével, F.; Desponds, C.; Utzschneider, D.T.; Zehnc, D.; Cusi, M.G.; Kuhlman, F.M.; et al. Type I interferons induced by endogenous or exogenous viral infections promote metastasis and relapse of leishmaniasis. Proc. Natl. Acad. Sci. USA 2017, 114, 4987–4992. [Google Scholar] [CrossRef]
- Rath, C.T.; Schnellrath, L.C.; Damaso, C.R.; de Arruda, L.B.; da Costa Vasconcelos, P.F.; Gomes, C.; Marcia Laurenti, D.; Silva, T.C.C.; de Carvalho Vivarini, A.; Fasel, N.; et al. Amazonian phlebovirus (Bunyaviridae) potentiates the infection of Leishmania (Leishmania) amazonensis: Role of the PKR/IFN/IL-10 axis. PLoS Trop. Negl. Dis. 2019, 13, e0007500. [Google Scholar] [CrossRef]
- Heirwegh, E.; MacLean, E.; He, J.; Kamhawi, S.; Sagan, S.M.; Olivier, M. Sandfly fever Sicilian virus-Leismania major co-infection modulates innate imflammatory response favoring myeloid cell infections and skin hyperinflammation. PLoS Negl. Trop. Dis. 2021, 15, e0009638. [Google Scholar] [CrossRef]
- dos Santos, J.V.; Freixi, P.F.; Vivarino, A.; de C Medina, J.M.; Caldas, L.A.; Attias, M.; Teixeira, K.L.D.; Silva, T.C.; Lopes, U.G. Endoplasmic stress affects the coinfection of Leishmania amazonensis and the Phlebovirus (Bunyaviridae) Icoaraci. Viruses 2022, 14, 1948. [Google Scholar] [CrossRef] [PubMed]
- Kaabi, B.; Zhioua, E. Modeling and comparative study of the spread of zoonotic visceral leishmaniasis from Northern to Central Tunisia. Acta Trop. 2018, 178, 19–26. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Number of Sandflies | Total (%) | |
|---|---|---|---|
| Male | Female | ||
| P. perniciosus | 221 | 120 | 341 (41.94) |
| P. perfiliewi | 135 | 120 | 255 (31.36) |
| S. minuta parotti | 164 | 49 | 213 (26.19) |
| P. papatasi | 3 | 6 | 3 (0.36) |
| P. longicuspis | 1 | 0 | 1 (0.12) |
| Total | 524 | 289 | 813 (100) |
| Date of Collection | Positive Pools | Nbr of Sandflies per Pool (Sex) | PCR Leishmania ITS-rDNA | RT-PCR, Nphlebo/A TOSV 2- |
|---|---|---|---|---|
| 7 September | T96 | 30 (F) | + (L. infantum) | - |
| T99 | 30 (F) | - | + (TOSV) | |
| T114 | 30 (M) | - | + (TOSV) | |
| T117 | 24 (F) | - | + (TOSV) | |
| 21 September | T139 | 30 (F) | - | + (TOSV) |
| T145 | 30 (M) | - | + (TOSV) | |
| 31 September | T153 | 30 (F) | + (L. infantum) | - |
| 8 October | T173 | 12 (F) | - | + (TOSV) |
| Date | Dog A | Dog B | Dog C | Healthy Dog | |||||
|---|---|---|---|---|---|---|---|---|---|
| Start of dogs exposition | 22 July 2020 | L. i | TOSV | L. i | TOSV | L. i | TOSV | L. i | TOSV |
| End of dogs exposition | 19 November 2020 | ||||||||
| Day 0 post-blood feeding | 20 November 2020 | - | - | + | - | - | + | - | - |
| Day 7 post-blood feeding | 26 November 2020 | - | - | + | - | - | + | - | - |
| 1 Dog C7 | 2 T99 | 3 T114 | 4 T117 | 5 T139 | 6 T145 | 7 T173 | ||
|---|---|---|---|---|---|---|---|---|
| 1 | Dog C7 | 100/0 | 0.005 | 0.000 | 0.009 | 0.000 | 0.000 | 0.000 |
| 2 | T99 | 99.5 | 100/0 | 0.005 | 0.014 | 0.005 | 0.005 | 0.005 |
| 3 | T114 | 100 | 99.5 | 100/0 | 0.009 | 0.000 | 0.000 | 0.000 |
| 4 | T117 | 99.1 | 98.6 | 99.1 | 100/0 | 0.009 | 0.009 | 0.009 |
| 5 | T139 | 100 | 99.5 | 100 | 99.1 | 100/0 | 0.000 | 0.000 |
| 6 | T145 | 100 | 99.5 | 100 | 99.1 | 100 | 100/0 | 0.000 |
| 7 | T173 | 100 | 99.5 | 100 | 99.1 | 100 | 100 | 100/0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dachraoui, K.; Chelbi, I.; Labidi, I.; Ben Osman, R.; Sayadi, A.; Ben Said, M.; Cherni, S.; Abbas, M.A.S.; Charrel, R.; Zhioua, E. The Role of the Leishmania infantum Infected Dogs as a Potential Reservoir Host for Toscana Virus in a Zoonotic Visceral Leishmaniasis Focus of Northern Tunisia. Viruses 2023, 15, 1012. https://doi.org/10.3390/v15041012
Dachraoui K, Chelbi I, Labidi I, Ben Osman R, Sayadi A, Ben Said M, Cherni S, Abbas MAS, Charrel R, Zhioua E. The Role of the Leishmania infantum Infected Dogs as a Potential Reservoir Host for Toscana Virus in a Zoonotic Visceral Leishmaniasis Focus of Northern Tunisia. Viruses. 2023; 15(4):1012. https://doi.org/10.3390/v15041012
Chicago/Turabian StyleDachraoui, Khalil, Ifhem Chelbi, Imen Labidi, Raja Ben Osman, Aida Sayadi, Mourad Ben Said, Saifedine Cherni, Mohammed Abdo Saghir Abbas, Rémi Charrel, and Elyes Zhioua. 2023. "The Role of the Leishmania infantum Infected Dogs as a Potential Reservoir Host for Toscana Virus in a Zoonotic Visceral Leishmaniasis Focus of Northern Tunisia" Viruses 15, no. 4: 1012. https://doi.org/10.3390/v15041012