
Meta-Transcriptomic Analysis Reveals Novel RNA Viruses in Hippocampus erectus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Information
2.2. RNA Extraction and Sequencing
2.3. Virus Discovery and Confirmation
2.4. RT-PCR
2.5. Genome Annotation and Phylogenetic Analyses
3. Results
3.1. Overview of the Virome in H. erectus
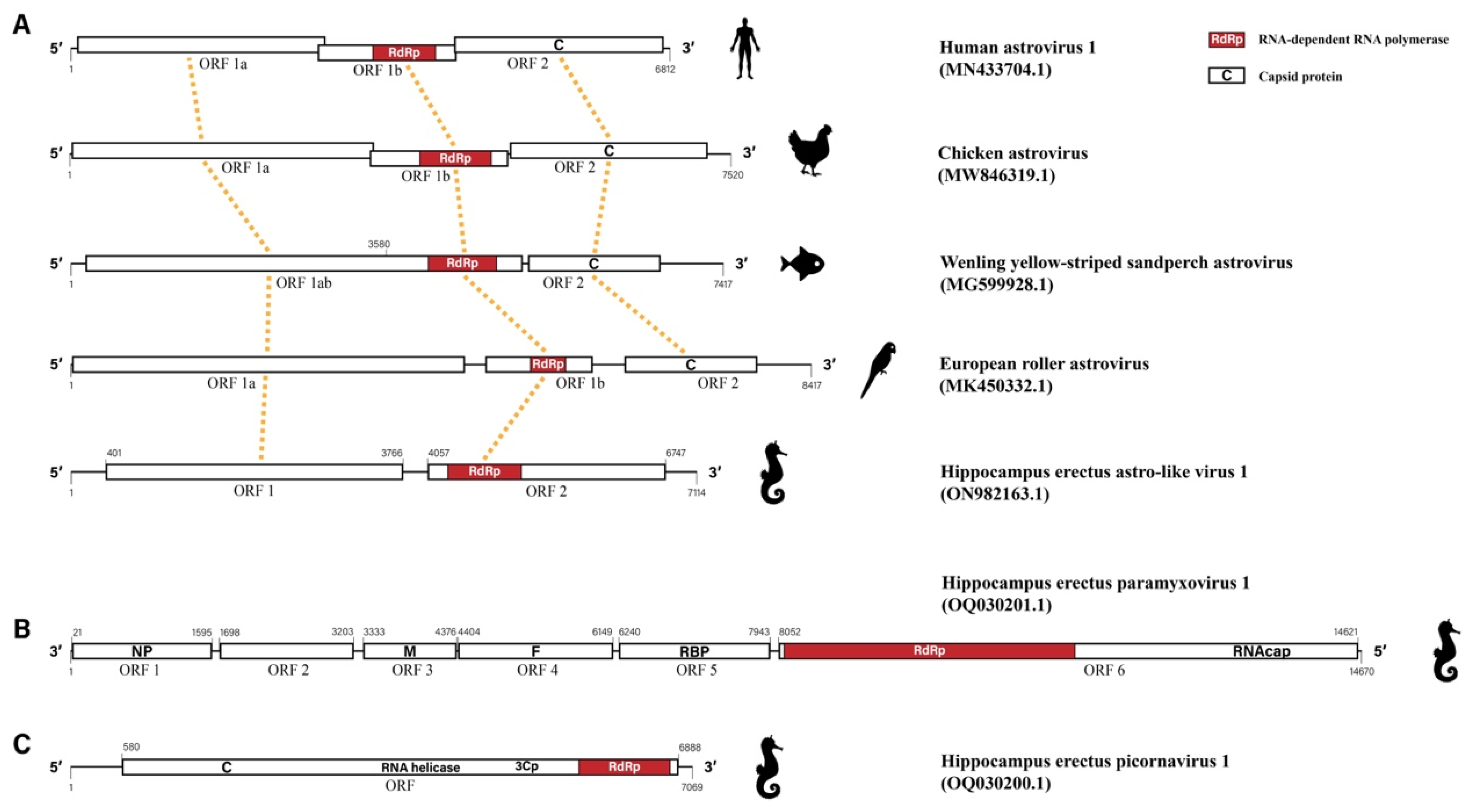
3.2. Hippocampus erectus Astro-like Virus 1
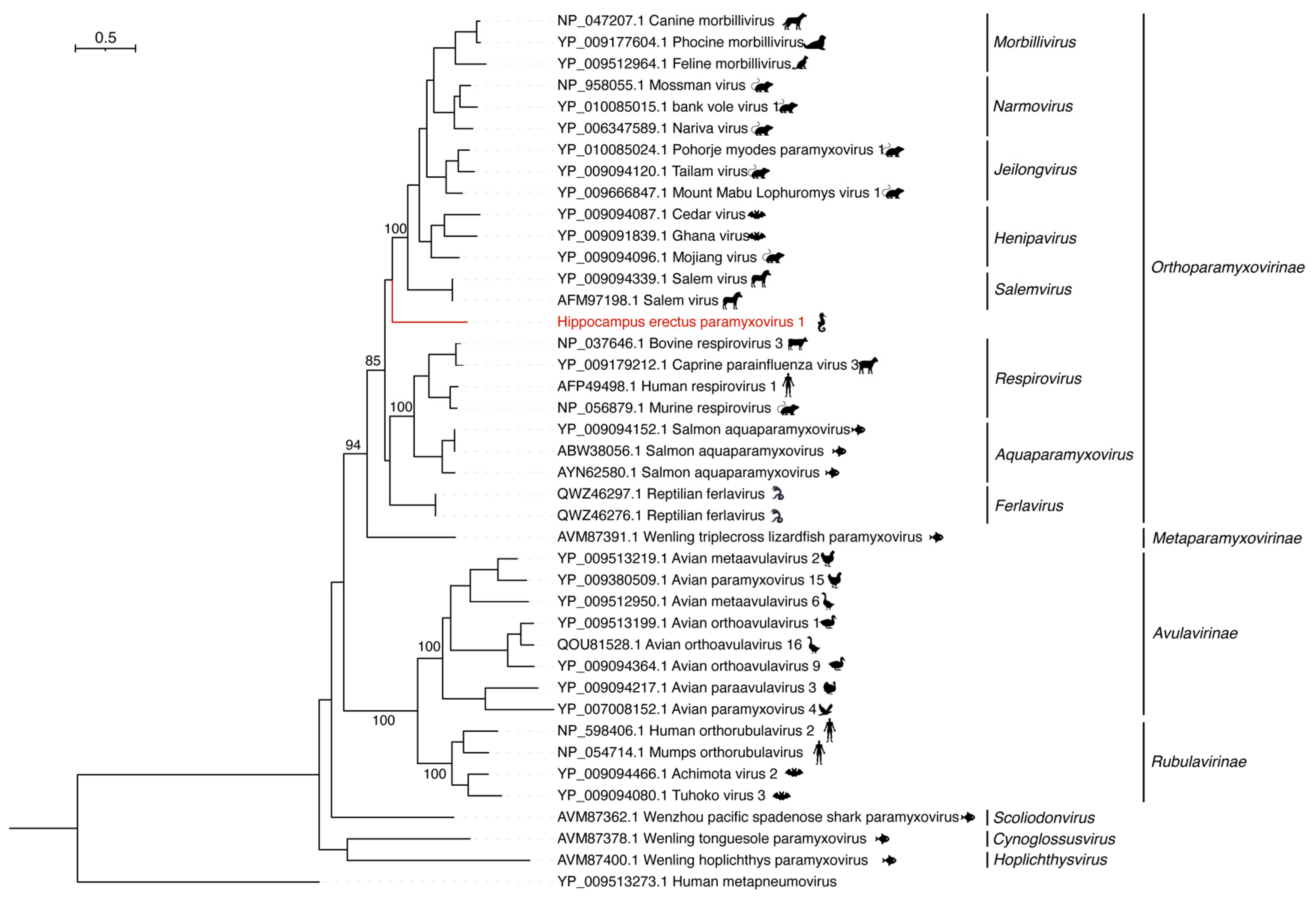
3.3. Hippocampus erectus Paramyxovirus 1
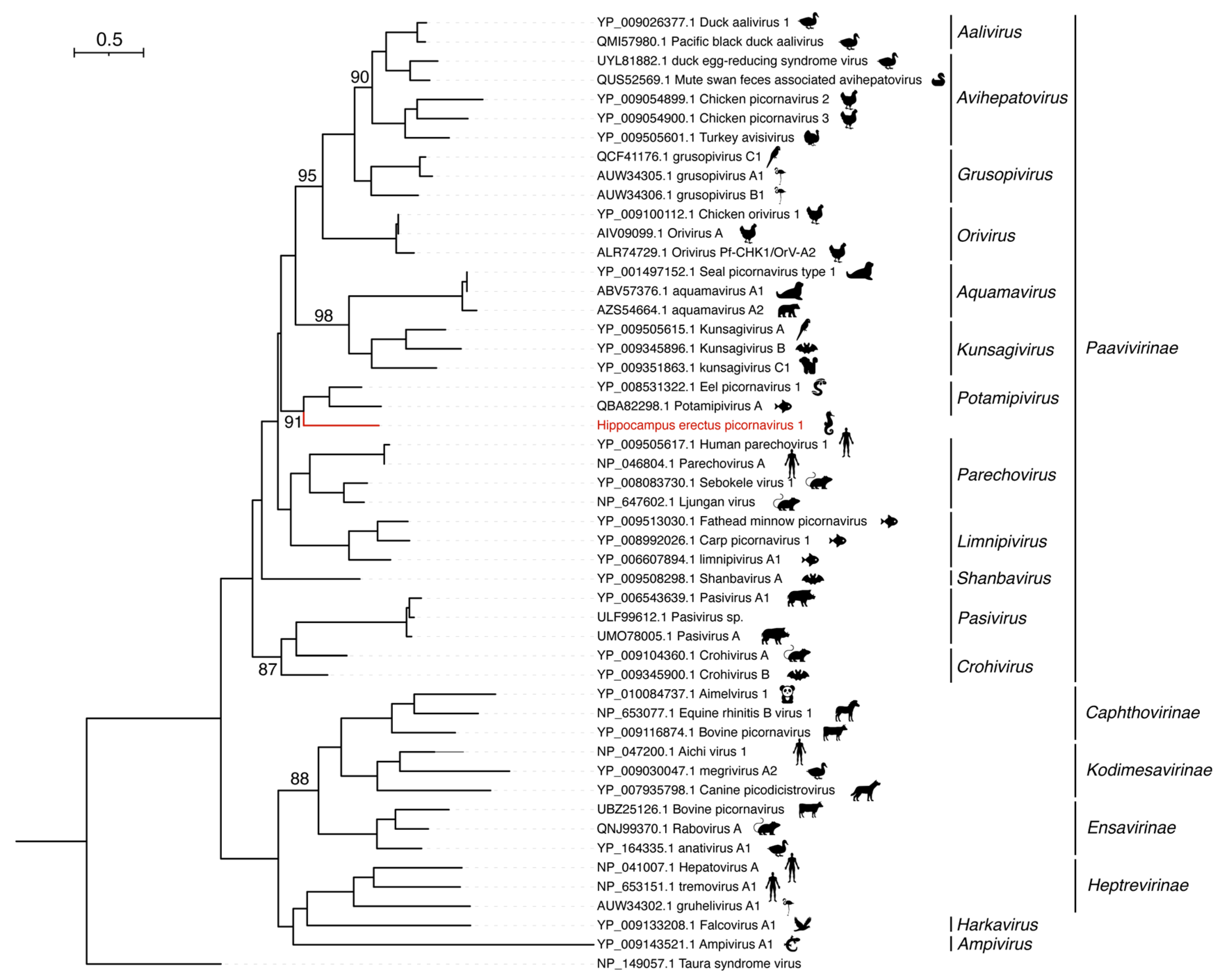
3.4. Hippocampus erectus Picornavirus 1
3.5. Nervous Necrosis Virus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.I.; Silas, S.; Wang, Y.; Wu, S.; Bocek, M.; Kazlauskas, D.; Krupovic, M.; Fire, A.; Dolja, V.V.; Koonin, E.V. Doubling of the known set of RNA viruses by metagenomic analysis of an aquatic virome. Nat. Microbiol. 2020, 5, 1262–1270. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liu, S.; Zhu, L.; Wan, X.; Liu, Q.; Qiu, L.; Zou, P.; Zhang, Q.; Huang, J. Complete genome sequence of an isolate of a novel genotype of yellow head virus from Fenneropenaeus chinensis indigenous in China. Arch. Virol. 2017, 162, 1149–1152. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Jang-Liaw, N.-H.; Lin, Y.-S.; Fang, Y.-C.; Shao, K.-T. Authenticating the use of dried seahorses in the traditional Chinese medicine market in Taiwan using molecular forensics. J. Food Drug Anal. 2013, 21, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Fan, S.; Zhang, Y.; Xu, M.; Zhang, H.; Yang, Y.; Lee, A.P.; Woltering, J.M.; Ravi, V.; Gunter, H.M.; et al. The seahorse genome and the evolution of its specialized morphology. Nature 2016, 540, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Goffredo, S.; Piccinetti, C.; Zaccanti, F. Volunteers in Marine Conservation Monitoring: A Study of the Distribution of Seahorses Carried Out in Collaboration with Recreational Scuba Divers. Conserv. Biol. 2004, 18, 1492–1503. [Google Scholar] [CrossRef] [Green Version]
- Boehm, J.T.; Waldman, J.; Robinson, J.D.; Hickerson, M.J. Population Genomics Reveals Seahorses (Hippocampus erectus) of the Western Mid-Atlantic Coast to Be Residents Rather than Vagrants. PLoS ONE 2015, 10, e0116219. [Google Scholar] [CrossRef] [Green Version]
- Pollom, R. Hippocampus erectus. In The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2017; Volume 2017, p. e.T10066A20191442. [Google Scholar]
- Huang, J.; Qin, G.; Zhang, B.; Tan, S.; Sun, J.; Lin, Q. Effects of food, salinity, and ammonia-nitrogen on the physiology of juvenile seahorse (Hippocampus erectus) in two typical culture models in China. Aquaculture 2020, 520, 734965. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, H.; Wang, X.; Wang, L.; Ji, D.; Liu, H.; Wang, K. Novel co-infection by ciliates and barentsia outbreak in large-scale cultured lined seahorse (Hippocampus erectus) in the northern China. Aquac. Res. 2020, 51, 867–872. [Google Scholar] [CrossRef]
- Dong, X.; Hu, T.; Liu, Q.; Li, C.; Sun, Y.; Wang, Y.; Shi, W.; Zhao, Q.; Huang, J. A Novel Hepe-Like Virus from Farmed Giant Freshwater Prawn Macrobrachium rosenbergii. Viruses 2020, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chen, X.; Qi, J.; He, L.; Luo, H.; Lin, J.; Qiu, F.; Wang, Q.; Zheng, L. Isolation and identification of a new strain of nervous necrosis virus from the big-belly seahorse Hippocampus abdominalis. Virol. J. 2022, 19, 109. [Google Scholar] [CrossRef]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Strother, K.O.; Mundt, E.; Zsak, L.; Day, J.M.; Spackman, E. Molecular characterization of avian astroviruses. Arch. Virol. 2011, 156, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, F.A.I.V.; San, S.; Hasan, M.K.; Wang, D.; Jones-Engel, L.; Schultz-Cherry, S. Non-Human Primates Harbor Diverse Mammalian and Avian Astroviruses Including Those Associated with Human Infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed]
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Lin, X.D.; Chen, X.; Tian, J.H.; Chen, L.J.; Li, K.; Wang, W.; Eden, J.S.; Shen, J.J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef]
- Noton, S.L.; Fearns, R. Initiation and regulation of paramyxovirus transcription and replication. Virology 2015, 479–480, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Jackson, T.; Belsham, G.J. Picornaviruses: A View from 3A. Viruses 2021, 13, 456. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, C.; Yang, M.; Huang, F.; Duan, X.; Wang, S.; Cheng, H.; Yang, H.; Zhao, H.; Qin, Q. Single-cell RNA-seq landscape midbrain cell responses to red spotted grouper nervous necrosis virus infection. PLoS Pathog. 2021, 17, e1009665. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Sample | Batch | Group | RPKM a |
|---|---|---|---|---|
| Hippocampus erectus astro-like virus 1 | FRRL190067409 | 201907 | Normal | 8.92 |
| FRRL190067410 | Normal | - b | ||
| FRRL190067412 | Unhealthy | 15.68 | ||
| FRRL190067411 | Unhealthy | 13.72 | ||
| FRRL192024264 | 201911 | Normal | 1.79 | |
| FRRL192024267 | Unhealthy | 4.09 | ||
| Hippocampus erectus paramyxovirus 1 | FRRL190067409 | 201907 | Normal | 16.94 |
| FRRL190067410 | Normal | - b | ||
| FRRL190067412 | Unhealthy | - b | ||
| FRRL190067411 | Unhealthy | 7.61 | ||
| FRRL192024264 | 201911 | Normal | - b | |
| FRRL192024267 | Unhealthy | - b | ||
| Hippocampus erectus picornavirus 1 | FRRL190067409 | 201907 | Normal | - b |
| FRRL190067410 | Normal | - b | ||
| FRRL190067412 | Unhealthy | - b | ||
| FRRL190067411 | Unhealthy | 166.02 | ||
| FRRL192024264 | 201911 | Normal | 17.79 | |
| FRRL192024267 | Unhealthy | 38.56 | ||
| nervous necrosis virus | FRRL190067409 | 201907 | Normal | - b |
| FRRL190067410 | Normal | - b | ||
| FRRL190067412 | Unhealthy | 6610.70 (RdRp c) 6125.40 (cp d) | ||
| FRRL190067411 | Unhealthy | - b | ||
| FRRL192024264 | 201911 | Normal | - b | |
| FRRL192024267 | Unhealthy | - b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Ren, Z.; Guo, X.; Wang, Y.; Meng, F.; Shi, W.; Wang, X.; Dong, X. Meta-Transcriptomic Analysis Reveals Novel RNA Viruses in Hippocampus erectus. Viruses 2023, 15, 772. https://doi.org/10.3390/v15030772
Zhang F, Ren Z, Guo X, Wang Y, Meng F, Shi W, Wang X, Dong X. Meta-Transcriptomic Analysis Reveals Novel RNA Viruses in Hippocampus erectus. Viruses. 2023; 15(3):772. https://doi.org/10.3390/v15030772
Chicago/Turabian StyleZhang, Fan, Zhihao Ren, Xiaomeng Guo, Yiting Wang, Fanzeng Meng, Weifeng Shi, Xinping Wang, and Xuan Dong. 2023. "Meta-Transcriptomic Analysis Reveals Novel RNA Viruses in Hippocampus erectus" Viruses 15, no. 3: 772. https://doi.org/10.3390/v15030772