
An Update on Eukaryotic Viruses Revived from Ancient Permafrost
 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Permafrost Sampling
2.2. Sample Preparation for Culturing
2.3. Detection of Virus Infection
2.4. Preliminary Identification of Infecting Viruses
2.5. Nomenclature of New Isolates
2.6. Further Characterization of New Virus Isolates
2.7. Viral Genome Sequencing
2.8. Deposition of Sequences
- Pandoravirus talik (strain Y4): OQ413801
- Pandoravirus lena (strain DY0): OQ411594-OQ411599
- Pandoravirus mammoth (strain Mm38): OQ411600-OQ411601
- Megavirus mammoth (strain Yana14): OQ411602
- Pithovirus mammoth (strain Yana14): OQ413582
- Cedratvirus duvanny (strain DY1): OQ413581
- Cedratvirus lena (strain DY0): OQ413577-OQ413579, OQ41358
- Pacmanvirus lupus (strain Tums2): OQ411603
2.9. Design of Virus-Specific PCR Primers
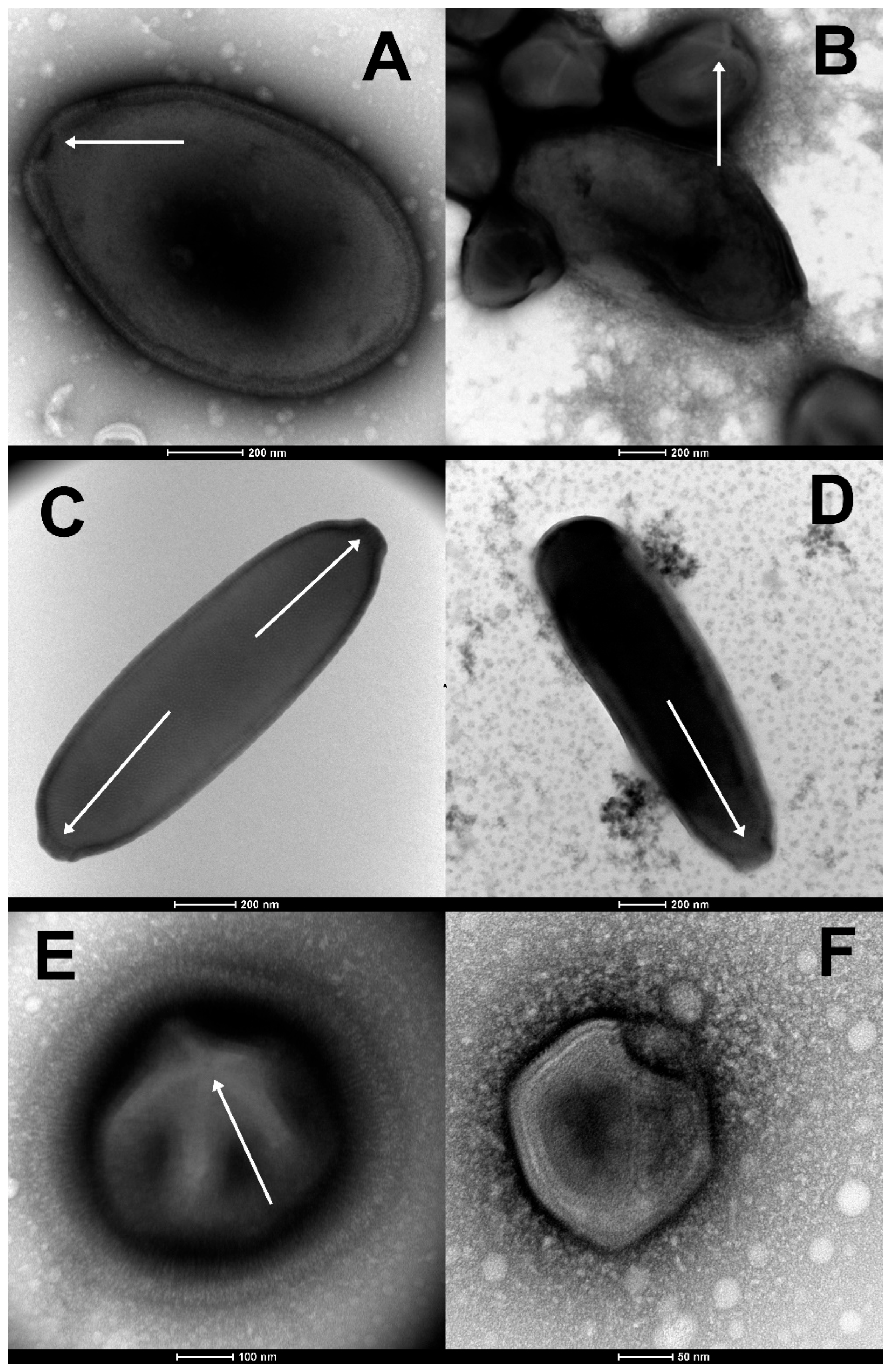
3. Results
3.1. Pandoraviruses
3.2. Cedratvirus and Pithovirus Isolates
3.3. Megavirus mammoth
3.4. Pacmanvirus lupus
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. 2022: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., Okem, A., Rama, B., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; 3056p. [Google Scholar] [CrossRef]
- Palmer, T.; Stevens, B. The scientific challenge of understanding and estimating climate change. Proc. Natl. Acad. Sci. USA 2019, 116, 24390–24395. [Google Scholar] [CrossRef] [Green Version]
- Rantanen, M.; Karpechko, A.Y.; Lipponen, A.; Nordling, K.; Hyvärinen, O.; Ruosteenoja, K.; Vihma, T.; Laaksonen, A. The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 2022, 3, 168. [Google Scholar] [CrossRef]
- Biskaborn, B.K.; Smith, S.L.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.A.; Schoeneich, P.; Romanovsky, V.E.; Lewkowicz, A.G.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264. [Google Scholar] [CrossRef] [Green Version]
- Turetsky, M.R.; Abbott, B.W.; Jones, M.C.; Walter Anthony, K.; Olefeldt, D.; Schuur, E.A.; Koven, C.; McGuire, A.D.; Grosse, G.; Kuhry, P.; et al. Permafrost collapse is accelerating carbon release. Nature 2019, 569, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M.; Nitze, I.; Strauss, J.; Günther, F.; Wetterich, S.; Kizyakov, A.; Fritz, M.; Opel, T.; Grigoriev, M.N.; Maksimov, G.T.; et al. Rapid fluvio-thermal erosion of a yedoma permafrost cliff in the Lena River Delta. Front. Earth Sci. 2020, 8, 336. [Google Scholar] [CrossRef]
- Günther, F.; Overduin, P.P.; Sandakov, A.V.; Grosse, G.; Grigoriev, M.N. Short- and long-term thermo-erosion of ice-rich permafrost coasts in the Laptev Sea region. Biogeosciences 2013, 10, 4297–4318. [Google Scholar] [CrossRef] [Green Version]
- Murton, J.B.; Opel, T.; Toms, P.; Blinov, A.; Fuchs, M.; Wood, J.; Gartner, A.; Merchel, S.; Rugel, G.; Savvinov, G.; et al. A multimethod dating study of ancient permafrost, Batagay megaslump, East Siberia. Quat. Res. 2022, 105, 1–22. [Google Scholar] [CrossRef]
- Kizyakov, A.I.; Wetterich, S.; Günther, F.; Opel, T.; Jongejans, L.L.; Courtin, J.; Meyer, H.; Shepelev, A.G.; Syromyatnikov, I.I.; Fedorov, A.N.; et al. Landforms and Degradation Pattern of the Batagay Thaw Slump, Northeastern Siberia. Geomorphology 2023, 420, 108501. [Google Scholar] [CrossRef]
- Gilichinsky, D.; Wagener, S. Microbial life in permafrost: A historical review. Permafr. Periglac. Process. 1995, 6, 243–250. [Google Scholar] [CrossRef]
- Burkert, A.; Douglas, T.A.; Waldrop, M.P.; Mackelprang, R. Changes in the Active, Dead, and Dormant Microbial Community Structure across a Pleistocene Permafrost Chronosequence. Appl. Environ. Microbiol. 2019, 85, e02646-18. [Google Scholar] [CrossRef] [Green Version]
- Hultman, J.; Waldrop, M.; Mackelprang, R.; David, M.M.; McFarland, J.; Blazewicz, S.J.; Harden, J.; Turetsky, M.R.; McGuire, A.D.; Shah, M.B.; et al. Multi-omics of permafrost, active layer and thermokarst bog soil microbiomes. Nature 2015, 521, 208–212. [Google Scholar] [CrossRef]
- Malavin, S.; Shmakova, L.; Claverie, J.-M.; Rivkina, E. Frozen Zoo: A collection of permafrost samples containing viable protists and their viruses. Biodivers. Data J. 2020, 8, e51586. [Google Scholar] [CrossRef]
- Lydolph, M.C.; Jacobsen, J.; Arctander, P.; Gilbert, M.T.; Gilichinsky, D.A.; Hansen, A.J.; Willerslev, E.; Lange, L. Beringian paleoecology inferred from permafrost-preserved fungal DNA. Appl. Environ. Microbiol. 2005, 71, 1012–1017. [Google Scholar] [CrossRef] [Green Version]
- Mishra, U.; Hugelius, G.; Shelef, E.; Yang, Y.; Strauss, J.; Lupachev, A.; Harden, J.W.; Jastrow, J.D.; Ping, C.L.; Riley, W.J.; et al. Spatial heterogeneity and environmental predictors of permafrost region soil organic carbon stocks. Sci. Adv. 2021, 7, eaaz5236. [Google Scholar] [CrossRef]
- Walter, K.M.; Zimov, S.A.; Chanton, J.P.; Verbyla, D.; Chapin, F.S., 3rd. Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming. Nature 2006, 443, 71–75. [Google Scholar] [CrossRef]
- Vishnivetskaya, T.A.; Petrova, M.A.; Urbance, J.; Ponder, M.; Moyer, C.L.; Gilichinsky, D.A.; Tiedje, J.M. Bacterial community in ancient Siberian permafrost as characterized by culture and culture-independent methods. Astrobiology 2006, 6, 400–414. [Google Scholar] [CrossRef]
- Johnson, S.S.; Hebsgaard, M.B.; Christensen, T.R.; Mastepanov, M.; Nielsen, R.; Munch, K.; Brand, T.; Gilbert, M.T.; Zuber, M.T.; Bunce, M.; et al. Ancient bacteria show evidence of DNA repair. Proc. Natl. Acad. Sci. USA 2007, 104, 14401–14405. [Google Scholar] [CrossRef] [Green Version]
- Revich, B.A.; Podolnaya, M.A. Thawing of permafrost may disturb historic cattle burial grounds in East Siberia. Glob. Health Action 2011, 4, 8482. [Google Scholar] [CrossRef]
- Hueffer, K.; Drown, D.; Romanovsky, V.; Hennessy, T. Factors contributing to anthrax outbreaks in the circumpolar north. Ecohealth 2020, 17, 174–180. [Google Scholar] [CrossRef]
- Timofeev, V.; Bahtejeva, I.; Mironova, R.; Titareva, G.; Lev, I.; Christiany, D.; Borzilov, A.; Bogun, A.; Vergnaud, G. Insights from Bacillus anthracis strains isolated from permafrost in the tundra zone of Russia. PLoS ONE 2019, 14, e0209140. [Google Scholar] [CrossRef] [Green Version]
- Legostaeva, Y.; Gololobova, A.; Popov, V. Geochemical Risks of Diamond Mining in Siberia. Environ. Sci. Proc. 2021, 5, 4. [Google Scholar] [CrossRef]
- Murton, J.B.; Goslar, T.; Edwards, M.E.; Bateman, M.D.; Danilov, P.P.; Savvinov, G.N.; Gubin, S.V.; Ghaleb, B.; Haile, J.; Kanevskiy, M.; et al. Palaeoenvironmental Interpretation of Yedoma Silt (Ice Complex) Deposition as Cold-Climate Loess, Duvanny Yar, Northeast Siberia. Permafr. Periglac. Process. 2015, 26, 208–288. [Google Scholar] [CrossRef]
- Mackelprang, R.; Waldrop, M.P.; DeAngelis, K.M.; David, M.M.; Chavarria, K.L.; Blazewicz, S.J.; Rubin, E.M.; Jansson, J.K. Metagenomic analysis of a permafrost microbial community reveals a rapid response to thaw. Nature 2011, 480, 368–371. [Google Scholar] [CrossRef] [Green Version]
- Krivushin, K.; Kondrashov, F.; Shmakova, L.; Tutukina, M.; Petrovskaya, L.; Rivkina, E. Two metagenomes from late pleistocene northeast siberian permafrost. Genome Announc. 2015, 3, e01380-14. [Google Scholar] [CrossRef] [Green Version]
- Perron, G.G.; Whyte, L.; Turnbaugh, P.J.; Goordial, J.; Hanage, W.P.; Dantas, G.; Desai, M.M. Functional characterization of bacteria isolated from ancient arctic soil exposes diverse resistance mechanisms to modern antibiotics. PLoS ONE 2015, 10, e0069533. [Google Scholar] [CrossRef]
- Mackelprang, R.; Burkert, A.; Haw, M.; Mahendrarajah, T.; Conaway, C.H.; Douglas, T.A.; Waldrop, M.P. Microbial survival strategies in ancient permafrost: Insights from metagenomics. ISME J. 2017, 11, 2305–2318. [Google Scholar] [CrossRef] [Green Version]
- Taş, N.; Prestat, E.; Wang, S.; Wu, Y.; Ulrich, C.; Kneafsey, T.; Tringe, S.G.; Torn, M.S.; Hubbard, S.S.; Jansson, J.K. Landscape topography structures the soil microbiome in arctic polygonal tundra. Nat. Commun. 2018, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Lau, M.; Vishnivetskaya, T.; Lloyd, K.G.; Wang, W.; Wiggins, J.; Miller, J.; Pfiffner, S.; Rivkina, E.M.; Onstott, T.C. Predominance of Anaerobic, Spore-Forming Bacteria in Metabolically Active Microbial Communities from Ancient Siberian Permafrost. Appl. Environ. Microbiol. 2019, 85, e00560-19. [Google Scholar] [CrossRef] [Green Version]
- Abramov, A.; Vishnivetskaya, T.; Rivkina, E. Are permafrost microorganisms as old as permafrost? FEMS Microbiol. Ecol. 2021, 97, fiaa260. [Google Scholar] [CrossRef]
- Rigou, S.; Christo-Foroux, E.; Santini, S.; Goncharov, A.; Strauss, J.; Grosse, G.; Fedorov, A.N.; Labadie, K.; Abergel, C.; Claverie, J.M. Metagenomic survey of the microbiome of ancient Siberian permafrost and modern Kamchatkan cryosols. microLife 2022, 3, uqac003. [Google Scholar] [CrossRef]
- Walsh, C.; Wencewicz, T. Antibiotics: Challenges, Mechanisms, Opportunities, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Haan, T.J.; Drown, D.M. Unearthing antibiotic resistance associated with disturbance-induced permafrost thaw in interior Alaska. Microorganisms 2021, 9, 116. [Google Scholar] [CrossRef]
- De Cock, K.M.; Jaffe, H.W.; Curran, J.W. Reflections on 40 Years of AIDS. Emerg. Infect. Dis. 2021, 27, 1553–1560. [Google Scholar] [CrossRef]
- Miranda, M.N.S.; Pingarilho, M.; Pimentel, V.; Torneri, A.; Seabra, S.G.; Libin, P.J.K.; Abecasis, A.B. A Tale of Three Recent Pandemics: Influenza, HIV and SARS-CoV-2. Front. Microbiol. 2022, 13, 889643. [Google Scholar] [CrossRef]
- Nasir, A.; Romero-Severson, E.; Claverie, J.-M. Investigating the Concept and Origin of Viruses. Trends Microbiol. 2020, 28, 959–967. [Google Scholar] [CrossRef]
- Claverie, J.-M. Fundamental Difficulties Prevent the Reconstruction of the Deep Phylogeny of Viruses. Viruses 2020, 12, 1130. [Google Scholar] [CrossRef]
- Emerson, J.B.; Roux, S.; Brum, J.R.; Bolduc, B.; Woodcroft, B.J.; Jang, H.B.; Singleton, C.M.; Solden, L.M.; Naas, A.E.; Boyd, J.A.; et al. Host-linked soil viral ecology along a permafrost thaw gradient. Nat. Microbiol. 2018, 3, 870–880. [Google Scholar] [CrossRef] [Green Version]
- Reid, A.H.; Fanning, T.G.; Hultin, J.V.; Taubenberger, J.K. Origin and evolution of the 1918 "Spanish" influenza virus hemagglutinin gene. Proc. Natl. Acad. Sci. USA 1999, 96, 1651–1656. [Google Scholar] [CrossRef] [Green Version]
- Biagini, P.; Thèves, C.; Balaresque, P.; Géraut, A.; Cannet, C.; Keyser, C.; Nikolaeva, D.; Gérard, P.; Duchesne, S.; Orlando, L.; et al. Variola virus in a 300-year-old Siberian mummy. N. Engl. J. Med. 2012, 367, 2056–2058. [Google Scholar] [CrossRef] [Green Version]
- Legendre, M.; Bartoli, J.; Shmakova, L.; Jeudy, S.; Labadie, K.; Adrait, A.; Lescot, M.; Poirot, O.; Bertaux, L.; Bruley, C.; et al. Thirty-thousand-year-old distant relative of giant icosahedral DNA viruses with a Pandoravirus morphology. Proc. Natl. Acad. Sci. USA 2014, 111, 4274–4279. [Google Scholar] [CrossRef] [Green Version]
- Legendre, M.; Lartigue, A.; Bertaux, L.; Jeudy, S.; Bartoli, J.; Lescot, M.; Alempic, J.-M.; Ramus, C.; Bruley, C.; Labadie, K.; et al. In-depth study of Mollivirus sibericum, a new 30,000-y-old giant virus infecting Acanthamoeba. Proc. Natl. Acad. Sci. USA 2015, 112, E5327–E5335. [Google Scholar] [CrossRef] [Green Version]
- Jongejans, L.L.; Grosse, G.; Ulrich, M.; Fedorov, A.N.; Konstantinov, P.; Strauss, J. Radiocarbon ages of talik sediments of an alas lake and a yedoma lake in the Yukechi Alas, Siberia. PANGAEA 2019. [Google Scholar] [CrossRef]
- Shmelev, D.; Cherbunina, M.; Rogov, V.; Opfergelt, S.; Monhonval, A.; Strauss, J. Reconstructing Permafrost Sedimentological Characteristics and Post-depositional Processes of the Yedoma Stratotype Duvanny Yar, Siberia. Front. Earth Sci. 2021, 9, 727315. [Google Scholar] [CrossRef]
- Grigoriev, S.E.; Fisher, D.C.; Obadă, T.; Shirley, E.A.; Rountrey, A.N.; Savvinov, G.N.; Darima, K.; Garmaeva, D.K.; Novgorodov, G.P.; Cheprasov, M.Y.; et al. A woolly mammoth (Mammuthus primigenius) carcass from maly Lyakhovsky island (new siberian islands, Russian Federation). Quat. Int. 2017, 445, 89–103. [Google Scholar] [CrossRef]
- Pitulko, V.V.; Nikolsky, P.A.; Girya, E.Y.; Basilyan, A.E.; Tumskoy, V.E.; Koulakov, S.A.; Astakhov, S.N.; Pavlova, E.Y.; Anisimov, M.A. The Yana RHS site: Humans in the Arctic before the last glacial maximum. Science 2004, 303, 52–56. [Google Scholar] [CrossRef]
- Basilyan, A.E.; Anisimov, M.A.; Nikolskiy, P.A.; Pitulko, V.V. Wooly mammoth mass accumulation next to the Paleolithic Yana RHS site, Arctic Siberia: Its geology, age, and relation to past human activity. J. Archaeol. Sci. 2011, 38, 2461–2474. [Google Scholar] [CrossRef]
- Guglielmini, J.; Woo, A.C.; Krupovic, M.; Forterre, P.; Gaia, M. Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes. Proc. Natl. Acad. Sci. USA 2019, 116, 19585–19592. [Google Scholar] [CrossRef] [Green Version]
- Christo-Foroux, E.; Alempic, J.-M.; Lartigue, A.; Santini, S.; Labadie, K.; Legendre, M.; Abergel, C.; Claverie, J.-M. Characterization of Mollivirus kamchatka, the First Modern Representative of the Proposed Molliviridae Family of Giant Viruses. J. Virol. 2020, 94, e01997-19. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [Green Version]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S. show statistics for a multiple alignment file. Available online: http://pwet.fr/man/linux/commandes/alistat/ (accessed on 30 September 2020).
- Philippe, N.; Legendre, M.; Doutre, G.; Couté, Y.; Poirot, O.; Lescot, M.; Arslan, D.; Seltzer, V.; Bertaux, L.; Bruley, C.; et al. Pandoraviruses: Amoeba viruses with genomes up to 2.5 Mb reaching that of parasitic eukaryotes. Science 2013, 341, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Legendre, M.; Alempic, J.M.; Philippe, N.; Lartigue, A.; Jeudy, S.; Poirot, O.; Ta, N.T.; Nin, S.; Couté, Y.; Abergel, C.; et al. Pandoravirus Celtis Illustrates the Microevolution Processes at Work in the Giant Pandoraviridae Genomes. Front. Microbiol. 2019, 10, 430. [Google Scholar] [CrossRef] [Green Version]
- Andreani, J.; Aherfi, S.; Bou Khalil, J.Y.; Di Pinto, F.; Bitam, I.; Raoult, D.; Colson, P.; La Scola, B. Cedratvirus, a Double-Cork Structured Giant Virus, is a Distant Relative of Pithoviruses. Viruses 2016, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.; Mueller, L.; Thomas, V.; Pillonel, T.; Jacquier, N.; Greub, G. Cedratvirus lausannensis - digging into Pithoviridae diversity. Environ. Microbiol. 2017, 19, 4022–4034. [Google Scholar] [CrossRef] [Green Version]
- Rigou, S.; Santini, S.; Abergel, C.; Claverie, J.-M.; Legendre, M. Past and present giant viruses diversity explored through permafrost metagenomics. Nat. Commun. 2022, 13, 5853. [Google Scholar] [CrossRef]
- Raoult, D.; Audic, S.; Robert, C.; Abergel, C.; Renesto, P.; Ogata, H.; La Scola, B.; Suzan, M.; Claverie, J.-M. The 1.2-megabase genome sequence of Mimivirus. Science 2004, 306, 1344–1350. [Google Scholar] [CrossRef]
- Arslan, D.; Legendre, M.; Seltzer, V.; Abergel, C.; Claverie, J.-M. Distant Mimivirus relative with a larger genome highlights the fundamental features of Megaviridae. Proc. Natl. Acad. Sci. USA 2011, 108, 17486–17491. [Google Scholar] [CrossRef] [Green Version]
- Claverie, J.-M.; Abergel, C. Mimiviridae: An Expanding Family of Highly Diverse Large dsDNA Viruses Infecting a Wide Phylogenetic Range of Aquatic Eukaryotes. Viruses 2018, 10, 506. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Kuznetsov, Y.G.; Sun, S.; Hafenstein, S.L.; Kostyuchenko, V.A.; Chipman, P.R.; Suzan-Monti, M.; Raoult, D.; McPherson, A.; Rossmann, M.G. Structural studies of the giant mimivirus. PLoS Biol. 2009, 7, e92. [Google Scholar] [CrossRef]
- Notaro, A.; Poirot, O.; Garcin, E.D.; Nin, S.; Molinaro, A.; Tonetti, M.; De Castro, C.; Abergel, C. Giant viruses of the Megavirinae subfamily possess biosynthetic pathways to produce rare bacterial-like sugars in a clade-specific manner. microLife 2022, 3, uqac002. [Google Scholar] [CrossRef]
- Andreani, J.; Khalil, J.Y.B.; Sevvana, M.; Benamar, S.; Di Pinto, F.; Bitam, I.; Colson, P.; Klose, T.; Rossmann, M.G.; Raoult, D.; et al. Pacmanvirus, a New Giant Icosahedral Virus at the Crossroads between Asfarviridae and Faustoviruses. J. Virol. 2017, 91, e00212-17. [Google Scholar] [CrossRef] [Green Version]
- Lemieux, A.; Colby, G.A.; Poulain, A.J.; Aris-Brosou, S. Viral spillover risk increases with climate change in High Arctic lake sediments. Proc. Biol. Sci. 2022, 289, 20221073. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; Nerad, T.A.; Visvesvara, G.S. Characterization of a new pathogenic Acanthamoeba Species, A. byersi n. sp., isolated from a human with fatal amoebic encephalitis. J. Eukaryot. Microbiol. 2013, 60, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Nature World News. Available online: https://www.natureworldnews.com/articles/45267/20210222/prehistoric-animal-viruses-permafrost-caught-interest-russian-scientists.htm (accessed on 22 February 2021).




{kind=link}
{kind=link}
{kind=link}
| Sample # | GPS Coordinates | Description | Isolated Virus |
|---|---|---|---|
| 1 | 55°06′54″ N 159°57′48″ E | Surface soil, Shapina river bank, Kamchatka Modern | Cedratvirus kamchatka (strain P5) |
| 2 | 62°06′23;92″ N 129°48′35″ E | Lena river, Yakutsk Modern | Cedratvirus lena (strain DY0) Pandoravirus lena (strain DY0) |
| 3 | 61°45′39.1″ N E 130°28′28.78″ | Talik, −6.5 m below a lake, Yukechi Alas [43] Isolation: >53 y BP | Pandoravirus talik (strain Y4) |
| 4 | 68°38′21.1″ N 159°3′20.67″ E | Melting ice wedge Duvanny yar [23,44] Mixed ages | Cedratvirus duvanny (strain DY1) Pandoravirus duvanny (strain DY1) |
| 5 | 61°45′39.1″ N 130°28′28.78″ E | −16 m below a lake, Yukechi Alas [43] Isolation: >48,500 y BP | Pandoravirus yedoma (strain Y2) |
| 6 | 74°13′00″ N 141°03′48″ E | Woolly mammoth stomach content, Maly Lyakhovsky Island [45] Isolation: >28,600 y BP | Pandoravirus mammoth (strain Mm38) |
| 7 | 70°43′25″ N 135°25′47″ E | Soil with mammoth wool RHS paleolithic site, Yana river left bank [46,47] Isolation: >27,000 BP | Megavirus mammoth (strain Yana14) Pithovirus mammoth (strain Yana14) Pandoravirus mammoth (strain Yana14) |
| 8 * | 70°43′25″ N 135°25′47″ E | Fossil wolf (Canis lupus) intestinal content, RHS paleolithic site [46,47] Isolation: >27,000 y BP | Pandoravirus lupus (strain Tums1) |
| 9 * | 70°43′25″ N 135°25′47″ E | Fossil wolf (Canis lupus) intestinal content, RHS paleolithic site [46,47] Isolation: >27,000 y BP | Pacmanvirus lupus (strain Tums2) |
| Virus Family or Subgroup | Primer Sequences (Forward & Reverse) | Prototype Gene |
|---|---|---|
| Pandoraviridae | F: TCGTGGATCGACATTGGCGTGCAGTT R: CTGGTAGGTGACGGCAAAGTT | Pandoravirus. salinus (NC_022098) CDS_1260 Putative oxidoreductase |
| Cedratvirus | F: AAACCTAGGTTGCTAACTGTAGATCCTTG R: GGAACCAGCGTTACCGAGTGCATCTTC | Cedratvirus A11 (NC_032108) BQ3484_149 Hypothetical protein |
| Pithoviridae | F: GTGGTCCAAAACTGGAAGAACTA R: GCGTCAAGCTCAACATCAAGTTC | Pithovirus sibericum (NC_023423) pv_393 DNA/RNA helicase |
| Megavirinae (A, B, C lineages) | F: TGGAATAATGGTGATGGTATTGATGT R: ACTGGTACCTAATCCTTTGTAATATTT | M. chilensis (NC_016072) mg403 Topoisomerase 2 |
| Pacmanvirus | F: GTCTCAATGGGCCACTTGAGCTG R: CCCGCTCTTGACCTCTGGGTTCC | Pacmanvirus A23 (LT706986) PACV_217 Major Capsid Protein |
| Virus | Accession | Base Pairs (Contigs) | Amplicon Identity * |
|---|---|---|---|
| Pandoravirus salinus (prototype) | NC_022098 | 2,473,870 | 100% (1203/1203) |
| Pandoravirus celtis | MK174290 | 2,028,440 | 93% (1128/1203) |
| Pandoravirus quercus | NC_037667 | 2,077,288 | 93% (1125/1203) |
| Pandoravirus inopinatum | NC_026440 | 2,243,109 | 93% (1133/1203) |
| Pandoravirus dulcis | NC_021858 | 1,908,524 | 92% (1122/1203) |
| Pandoravirus neocaledonia | NC_037666 | 2,003,191 | 86% (1045/1203) |
| Pandoravirus macleodensis | NC_037665 | 1,838,258 | 86% (1039/1203) |
| Pandoravirus lena (strain DY0) | OQ411594-OQ411599 | 2,030,260 (6) | 93% (1131/1203) |
| Pandoravirus duvanny (strain DY1) | - | unassembled | 92% (1114/1203) |
| Pandoravirus talik (strain Y4) | OQ413801 | 1,817,546 (1) | 92% (1114/1203) |
| Pandoravirus mammoth (strain Yana14) | - | not yet sequenced | 91% (1104/1203) |
| Pandoravirus yedoma (strain Y2) | - | not yet sequenced | 91% (1095/1203) |
| Pandoravirus lupus (strain Tums1) | - | not yet sequenced | 91% (1092/1203) |
| Pandoravirus mammoth (strain Mm38) | OQ411600-OQ411601 | 1,776,082 (2) | 86% (1040/1203) |
| Virus | Accession # | Base Pairs | Amplicon Identity * |
|---|---|---|---|
| Cedratvirus A11 (prototype) | NC_032108 | 589,068 | 100% (1239/1239) |
| Cedratvirus lausannensis | LT907979 | 575,161 | 94% (1176/1239) |
| Cedratvirus kamchatka | MN873693 | 466,767 | 88% (1091/1239) |
| Cedratvirus lena (strain DY0) | OQ413577-OQ413579, OQ41358 | 465,544 | 87% (1090/1239) |
| Cedratvirus duvanny (strain DY1) | OQ413581 | 472,117 | 87% (1087/1239) |
| Pithovirus sibericum (P1084-T) | NC_023423 | 610,033 | 100% (593/593) |
| Pithovirus mammoth (strain Yana14) | OQ413582 | 610,309 | 97% (581/593) |
| Virus | Accession # | Base Pairs | Amplicon Identity * |
|---|---|---|---|
| Megavirus chilensis | NC_016072 | 1,259,197 | 100% (1497/1497) |
| Megavirus vitis | MG807319 | 1,242,360 | 99% (1493/1497) |
| Megavirus mammoth (strain Yana14) | OQ411602 | 1,260,651 | 99% (1493/1497) |
| Megavirus powai lake | KU877344 | 1,208,707 | 93% (1396/1497) |
| Megavirus baoshan | MH046811 | 1,224,839 | 92% (1379/1497) |
| Moumouvirus | NC_020104 | 1,021,348 | 83% (1248/1497) |
| Moumouvirus australiensis | MG807320 | 1,098,002 | 82% (1244/1497) |
| Mimivirus | NC_014649 | 1,181,549 | 77% (1158/1497) |
| Virus | Accession # | Base Pairs | Amplicon Identity * |
|---|---|---|---|
| Pacmanvirus A23 | NC_034383 | 395,405 | 100% (470/470) |
| Pacmanvirus S19 | MZ440852 | 418,588 | 93% (439/470) |
| Pacmanvirus lupus (strain Tums2) | OQ411603 | 407,705 | <85% (399/468) + two large insertions |
| Pacmanvirus lupus Predicted Protein | Pacmanvirus A23 NC_034383 %ID (aa) | Pacmanvirus S19 MZ440852 %ID (aa) | Faustovirus KJ614390 %ID (aa) | Kaumoebavirus NC_034249 %ID (aa) | Asfarviruses NC_044958 %ID (aa) |
|---|---|---|---|---|---|
| RNA polymerase (RPB1) ORF 302 | 79% (1124/1415) | 80% (1130/1415) | 49% (707/1434) | 42% (598/1429) | 41% (596/1457) |
| RNA polymerase (RPB2) ORF 33 | 85% (1093/1289) | 85% (1104/1301) | 55% (681/1241) | 44% (528/1211) | 43% (526/1228) |
| DNA polymerase (PolB) ORF 265 | 65% (1036/1591) | 65% (1032/1591) | 37% (513/1385) | 27.5% (344/1250) | 33% (382/1163) |
| Genome size | |||||
| 407,705 bp | 395,405 bp | 418,588 bp | 457–491 kb | 351–363 kb | 172–191 kb |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alempic, J.-M.; Lartigue, A.; Goncharov, A.E.; Grosse, G.; Strauss, J.; Tikhonov, A.N.; Fedorov, A.N.; Poirot, O.; Legendre, M.; Santini, S.; et al. An Update on Eukaryotic Viruses Revived from Ancient Permafrost. Viruses 2023, 15, 564. https://doi.org/10.3390/v15020564
Alempic J-M, Lartigue A, Goncharov AE, Grosse G, Strauss J, Tikhonov AN, Fedorov AN, Poirot O, Legendre M, Santini S, et al. An Update on Eukaryotic Viruses Revived from Ancient Permafrost. Viruses. 2023; 15(2):564. https://doi.org/10.3390/v15020564
Chicago/Turabian StyleAlempic, Jean-Marie, Audrey Lartigue, Artemiy E. Goncharov, Guido Grosse, Jens Strauss, Alexey N. Tikhonov, Alexander N. Fedorov, Olivier Poirot, Matthieu Legendre, Sébastien Santini, and et al. 2023. "An Update on Eukaryotic Viruses Revived from Ancient Permafrost" Viruses 15, no. 2: 564. https://doi.org/10.3390/v15020564