
A Capsid Protein Fragment of a Fusagra-like Virus Found in Carica papaya Latex Interacts with the 50S Ribosomal Protein L17

 , , , , ,
, , , , ,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
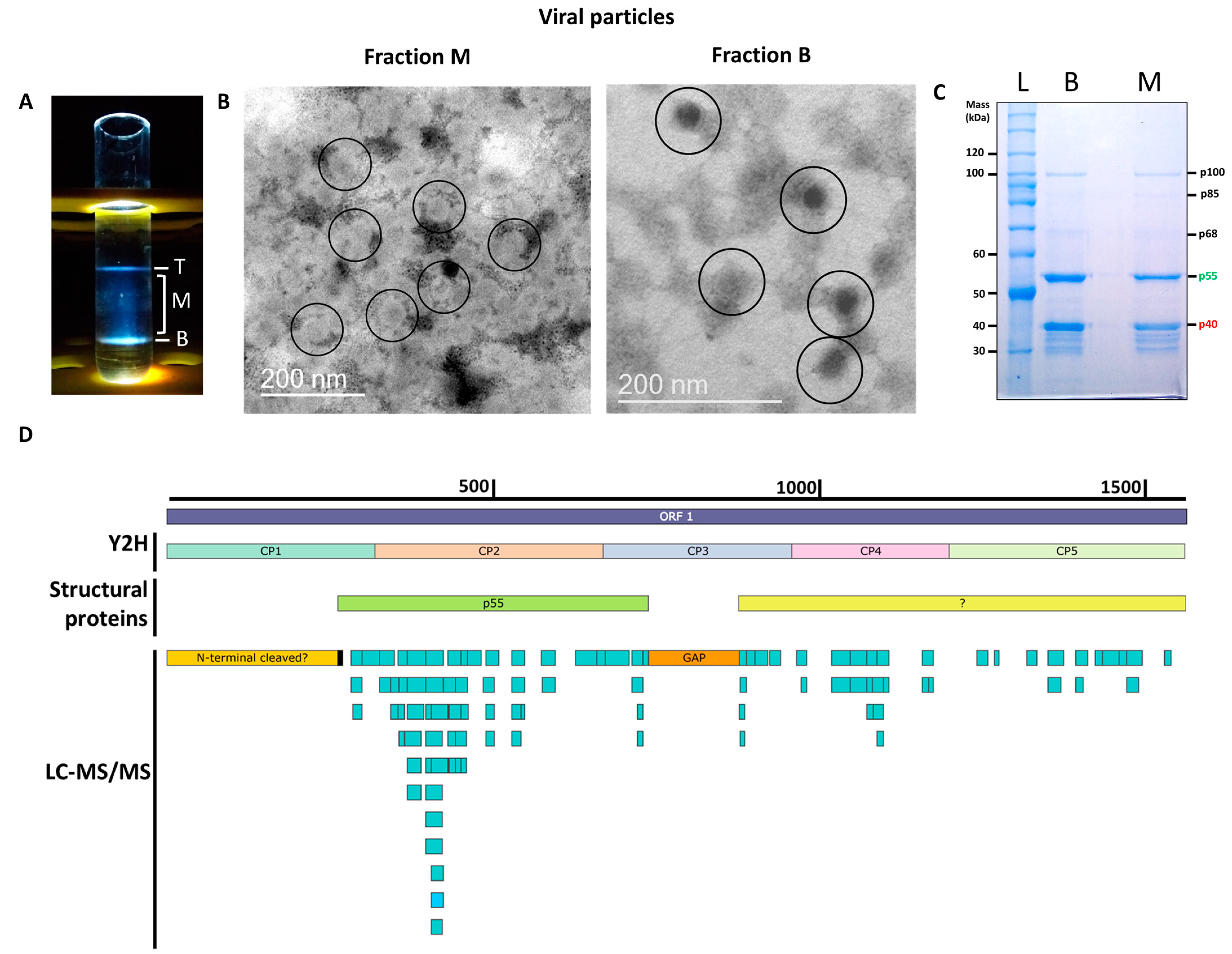
2.1. Virus Purification and Transmission Electron Microscopy Analysis
2.2. Protein Identification of Virion Preparations
2.2.1. Tandem Mass Spectrometry
2.2.2. Amino-Terminal Sequencing
2.3. P55 Structural Homology, Domain and CLEAVAGE Prediction
2.4. Cloning of PMeV ORF1 and AtRPL17
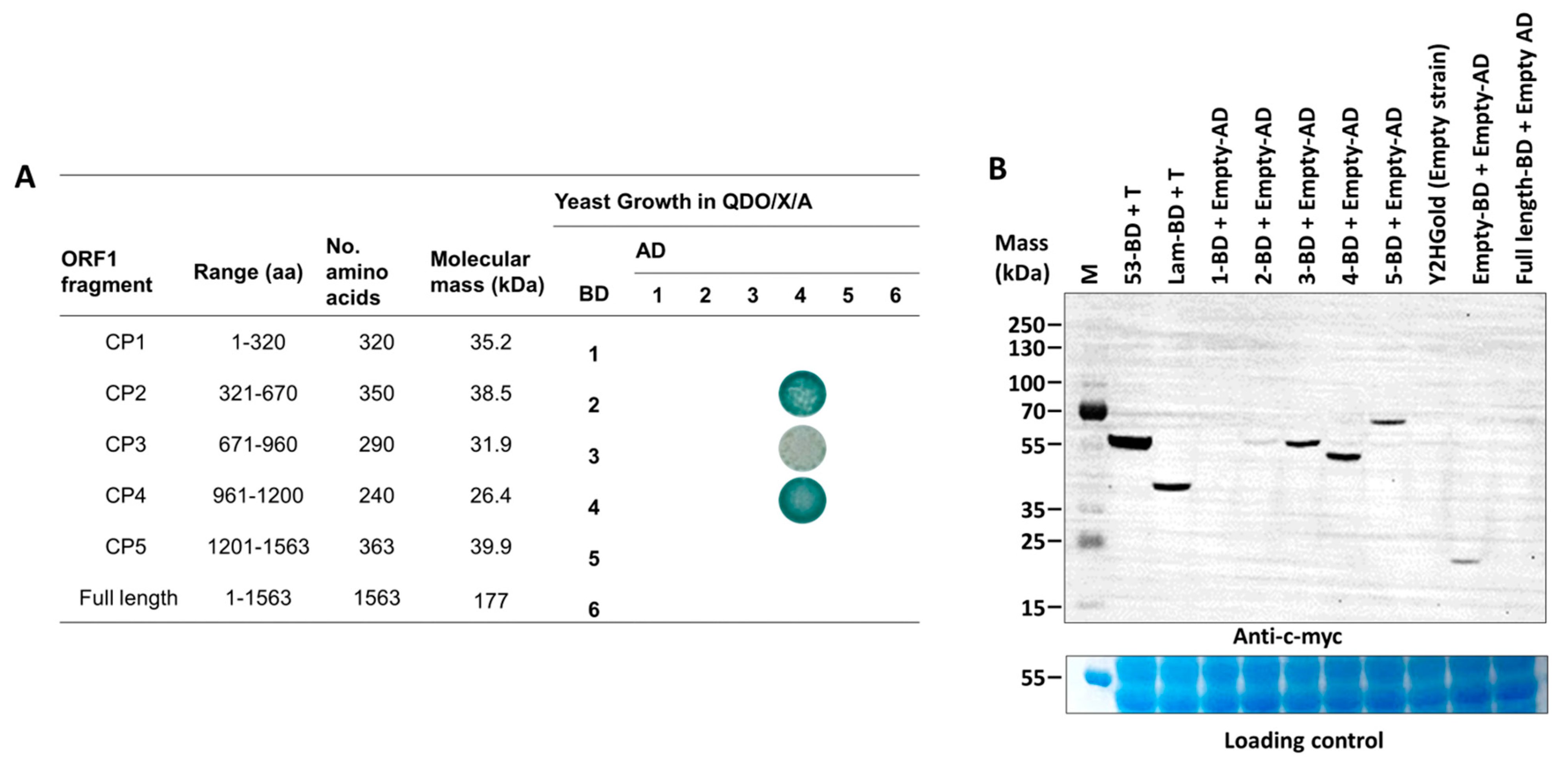
2.5. Binary Interactions and Library Screening Using Yeast Two-Hybrid Assay
2.6. Protein–Protein Interaction Network
2.7. Transient Expression and Detection of Protein Interaction in Nicotiana Benthamiana
3. Results
3.1. Identification of the PMeV Capsid-Protein Coding Region
3.2. PMeV p55 Share Conserved Domains with Mycovirus Capsid Proteins
3.3. A Fragment of p55 (CP2, aa 321–670) Interacts with a Viral Structural Protein Fragment, CP4 (aa 961–1200)
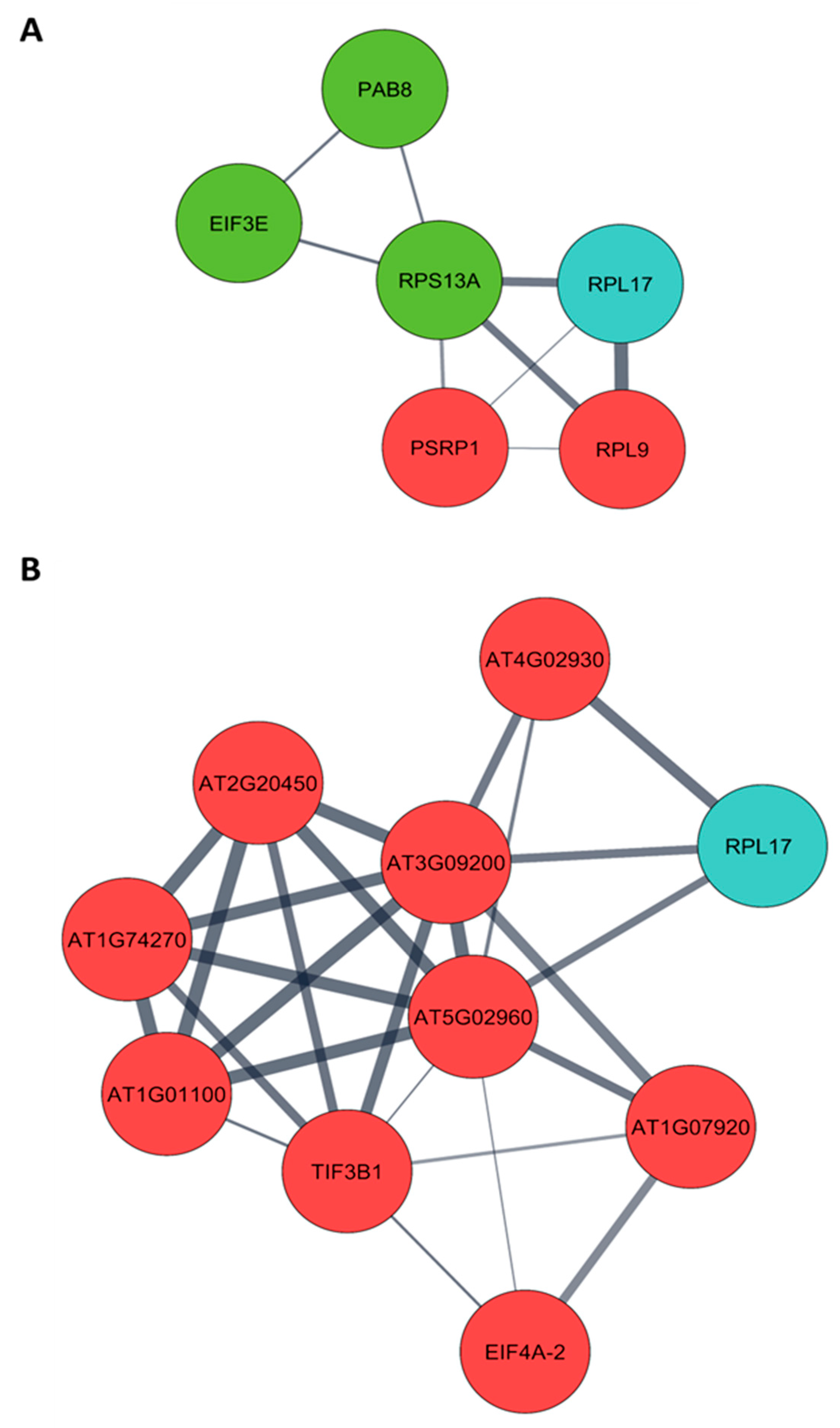
3.4. Translation and RNA-Silencing Complexes Co-Purify with the PMeV Complex
3.5. Arabidopsis Library Screening Using a C-Terminal PMeV ORF1 Fragment (aa 961–1200) Identified 28 Plant-Interacting Proteins
3.6. CP2 Putatively Associates with Translation-Related Proteins Differentially Expressed in Pre-Flowering C. papaya
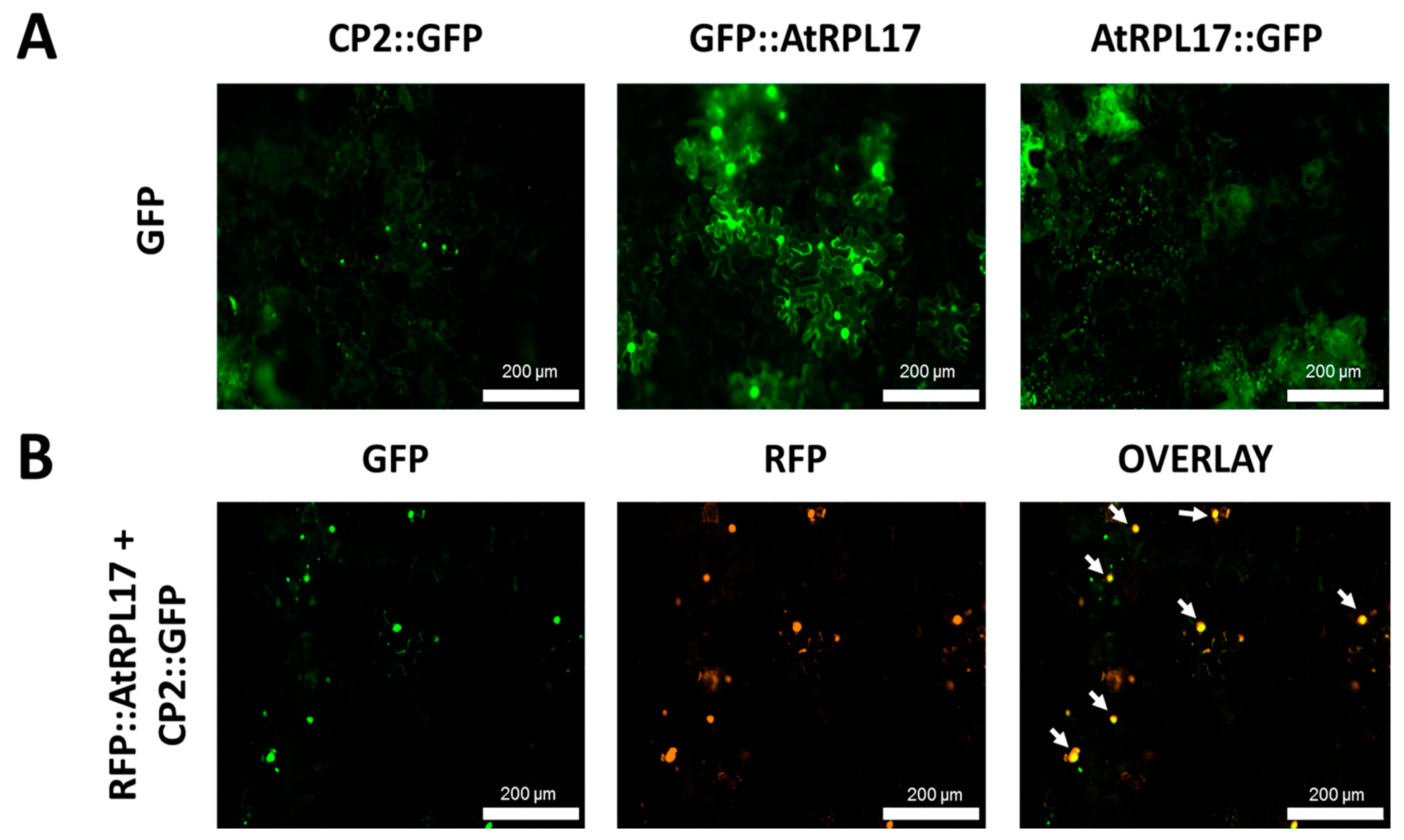
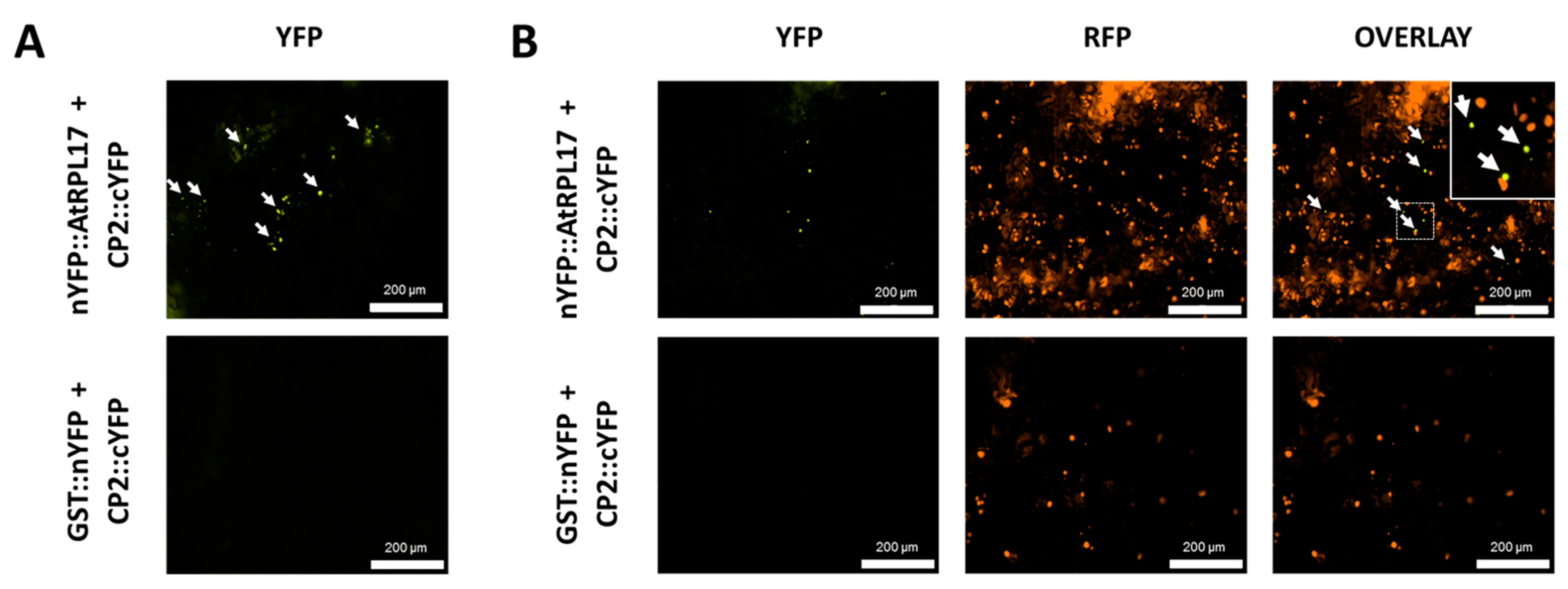
3.7. CP2 and AtRPL17 Co-Localize and Interact in Nicotiana Benthamiana Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kitajima, E.W.; Rodrigues, C.H.; Silveira, J.S.; Alves, F.; Ventura, J.A.; Aragão, F.J.L.; Oliveira, L.H.R. Association of isometric viruslike particles, restricted to laticifers, with “meleira” (sticky disease) of papaya (Carica papaya). Fitopatol. Bras. 1993, 18, 118. [Google Scholar]
- Maciel-Zambolim, E.; Kunieda-Alonso, S.; Matsuoka, K.; De Carvalho, M.; Zerbini, F. Purification and some properties of papaya meleira virus, a novel virus infecting papayas in Brazil. Plant Pathol. 2003, 52, 389–394. [Google Scholar] [CrossRef]
- Antunes, T.F.S.; Amaral, R.J.V.; Ventura, J.A.; Godinho, M.T.; Amaral, J.G.; Souza, F.O.; Zerbini, P.A.; Zerbini, F.M.; Fernandes, P.M.B. The dsRNA virus papaya meleira virus and an ssRNA virus are associated with papaya sticky disease. PLoS ONE 2016, 11, e0155240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, T.F.S.; Maurastoni, M.; Madroñero, L.J.; Fuentes, G.; Santamaría, J.M.; Ventura, J.A.; Abreu, E.F.; Fernandes, A.A.R.; Fernandes, P.M.B. Battle of three: The curious case of papaya sticky disease. Plant Dis. 2020, 104, 2754–2763. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Zhang, H.; Qiu, D.; Guo, L. Two novel relative double-stranded RNA mycoviruses infecting fusarium poae strain SX63. Int. J. Mol. Sci. 2016, 17, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Alam, M.M.; Zhang, R.; Hisano, S.; Suzuki, N. Proof of concept of the Yadokari nature: A capsidless replicase-encoding but replication-dependent positive-sense single-stranded RNA virus hosted by an unrelated double-stranded RNA Virus. J. Virol. 2021, 95, e00467–e00421. [Google Scholar] [CrossRef]
- Liu, J.; Carino, E.; Bera, S.; Gao, F.; May, J.P.; Simon, A.E. Structural analysis and whole genome mapping of a new type of plant virus subviral RNA: Umbravirus-like associated RNAs. Viruses 2021, 13, 646. [Google Scholar] [CrossRef]
- Evert, R.F. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; John Wiley & Sons: New York, NY, USA, 2006; pp. 489–490. [Google Scholar] [CrossRef]
- Rodrigues, S.P.; Da Cunha, M.; Ventura, J.A.; Fernandes, P.M.B. Effects of the papaya meleira virus on papaya latex structure and composition. Plant Cell Rep. 2009, 28, 861–871. [Google Scholar] [CrossRef]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef]
- Callaway, A.; Giesman-Cookmeyer, D.; Gillock, E.T.; Sit, T.L.; Lommel, S.A. The multifunctional capsid proteins of plant RNA viruses. Annu. Rev. Phytopathol. 2001, 39, 419–460. [Google Scholar] [CrossRef] [PubMed]
- Herranz, M.C.; Pallas, V.; Aparicio, F. Multifunctional roles for the N-terminal basic motif of alfalfa mosaic virus coat protein: Nucleolar/cytoplasmic shuttling, modulation of RNA-binding activity, and virion formation. Mol. Plant-Microbe Interact. 2012, 25, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Vaira, A.M.; Lim, H.S.; Bauchan, G.; Gulbronson, C.J.; Miozzi, L.; Vinals, N.; Natilla, A.; Hammond, J. The interaction of lolium latent virus major coat protein with ankyrin repeat protein NbANKr redirects it to chloroplasts and modulates virus infection. J. Gen. Virol. 2018, 99, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Bol, J.F. Role of capsid proteins. In Plant Virology Protocols: From Viral Sequence to Protein Function; Foster, G.D., Johansen, I.E., Hong, Y., Nagy, P.D., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 21–31. [Google Scholar] [CrossRef]
- Luque, D.; Mata, C.P.; Suzuki, N.; Ghabrial, S.A.; Castón, J.R. Capsid structure of dsRNA fungal viruses. Viruses 2018, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata, C.P.; Luque, D.; Gómez-Blanco, J.; Rodríguez, J.M.; González, J.M.; Suzuki, N.; Ghabrial, S.A.; Carrascosa, J.L.; Trus, B.L.; Castón, J.R. Acquisition of functions on the outer capsid surface during evolution of double-stranded RNA fungal viruses. PLoS Path. 2017, 13, e1006755. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.P.; Ventura, J.A.; Aguilar, C.; Nakayasu, E.S.; Almeida, I.C.; Fernandes, P.; Zingali, R.B. Proteomic analysis of papaya (Carica papaya L.) displaying typical sticky disease symptoms. Proteomics 2011, 11, 2592–2602. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.P.; Ventura, J.A.; Aguilar, C.; Nakayasu, E.S.; Choi, H.; Sobreira, T.J.P.; Nohara, L.L.; Wermelinger, L.S.; Almeida, I.C.; Zingali, R.B.; et al. Label-free quantitative proteomics reveals differentially regulated proteins in the latex of sticky diseased Carica papaya L. plants. J. Proteom. 2012, 75, 3191–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, E.; Werth, E.G.; Madroñero, L.J.; Ventura, J.A.; Rodrigues, S.P.; Hicks, L.M.; Fernandes, P.M.B. Label-free quantitative proteomic analysis of pre-flowering PMeV-infected Carica papaya L. J. Proteom. 2016, 151, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Madroñero, J.; Rodrigues, S.P.; Antunes, T.F.S.; Abreu, P.M.V.; Ventura, J.A.; Fernandes, A.A.R.; Fernandes, P.M.B. Transcriptome analysis provides insights into the delayed sticky disease symptoms in Carica papaya. Plant Cell Rep. 2018, 37, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Bos, L.; Huijberts, N.; Huttinga, H.; Maat, D.Z. Further characterization of dandelion yellow mosaic virus from lettuce and dandelion. Neth. J. Plant Pathol. 1983, 89, 207–222. [Google Scholar] [CrossRef]
- Quiñones-Vega, M.; Velásquez, E.; Sosa-Acosta, P.; Melo, A.; Garcez, P.P.; Nogueira, F.C.S.; Domont, G.B. Proteomic profiles of Zika virus-infected placentas bearing fetuses with microcephaly. Proteom.—Clin. Appl. 2022, 16, 2100042. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Holm, L. Dali server: Structural unification of protein families. Nucleic Acids Res. 2022, 50, W210–W215. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- de Lima, J.G.S.; Teixeira, D.G.; Freitas, T.T.; Lima, J.P.M.S.; Lanza, D.C.F. Evolutionary origin of 2A-like sequences in Totiviridae genomes. Virus Res. 2019, 259, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kushnirov, V.V. Rapid and reliable protein extraction from yeast. Yeast 2000, 16, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chakrabarty, R.; Banerjee, R.; Chung, S.-M.; Farman, M.; Citovsky, V.; Hogenhout, S.A.; Tzfira, T.; Goodin, M. pSITE vectors for stable integration or transient expression of autofluorescent protein fusions in plants: Probing Nicotiana benthamiana-virus interactions. Mol. Plant-Microbe Interact. 2007, 20, 740–750. [Google Scholar] [CrossRef]
- Martin, K.; Kopperud, K.; Chakrabarty, R.; Banerjee, R.; Brooks, R.; Goodin, M.M. Transient expression in Nicotiana benthamiana fluorescent marker lines provides enhanced definition of protein localization, movement and interactions in planta. Plant J. 2009, 59, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Ghabrial, S.A.; Nibert, M.L.; Patterson, J.L.; Wang, C.C. International Committee on Taxonomy of Viruses. Available online: https://ictv.global/taxonomy/ (accessed on 22 September 2022).
- Akinyemi, I.A.; Wang, F.; Chang, Z.-X.; Wu, Q. Genome characterization of the newly identified maize-associated totivirus Anhui. Arch. Virol. 2018, 163, 2929–2931. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cao, L.; Huang, Q.; Qian, Y.; Zhou, X. The complete genome sequence of a novel maize-associated totivirus. Arch. Virol. 2016, 161, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Quinto, R.A.; Espinoza-Lozano, R.F.; Mora-Pinargote, C.A.; Quito-Avila, D.F. Complete genome sequence of a variant of maize-associated totivirus from Ecuador. Arch. Virol. 2017, 162, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, X.; Wu, W.; Tan, G.; Fang, S.; Zhang, S.; Li, F. Identification and molecular characterization of panax notoginseng virus A, which may represent an undescribed novel species of the genus Totivirus, family Totiviridae. Arch. Virol. 2016, 161, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Huang, H.; Han, X.X.; Li, R.; Wu, L.P.; Wu, L. Identification and molecular characterization of tea-oil camellia-associated totivirus 1. Arch. Virol. 2021, 166, 2347–2351. [Google Scholar] [CrossRef]
- Ong, J.W.L.; Li, H.; Sivasithamparam, K.; Dixon, K.W.; Jones, M.G.K.; Wylie, S.J. Novel and divergent viruses associated with australian orchid-fungus symbioses. Virus Res. 2018, 244, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Cox, S.; Mayo, M.A.; Jones, A.T. The Occurrence of dsRNA species in apparently healthy and virus-infected ribes cultivars, and evidence that one such species originates from a member of the virus family Totiviridae. Eur. J. Plant Pathol. 2000, 106, 353–364. [Google Scholar] [CrossRef]
- Das, S.; Hisano, S.; Eusebio-Cope, A.; Kondo, H.; Suzuki, N. A Transfectable fusagravirus from a japanese strain of Cryphonectria carpinicola with spherical particles. Viruses 2022, 14, 1722. [Google Scholar] [CrossRef]
- Poulos, B.T.; Tang, K.F.J.; Pantoja, C.R.; Bonami, J.R.; Lightner, D.V. Purification and characterization of infectious myonecrosis virus of penaeid shrimp. J. Gen. Virol. 2006, 87, 987–996. [Google Scholar] [CrossRef]
- Rampersad, S.; Tennant, P. Chapter 3—Replication and expression strategies of viruses. In Viruses; Tennant, P., Fermin, G., Foster, J.E., Eds.; Academic Press: New York, NY, USA, 2018; pp. 55–82. [Google Scholar] [CrossRef]
- Babé, L.M.; Craik, C.S. Viral proteases: Evolution of diverse structural motifs to optimize function. Cell 1997, 91, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Ji, B.; Yu, B. Laticifer ultrastructural and immunocytochemical studies of papain in Carica papaya. Acta Bot. Sin. 1994, 36, 497–501. [Google Scholar]
- Lagunas-Rangel, F.A.; Kameyama-Kawabe, L.Y.; Bermúdez-Cruz, R.M. Giardiavirus: An update. Parasitol. Res. 2021, 120, 1943–1948. [Google Scholar] [CrossRef]
- Yu, D.; Wang, C.C.; Wang, A.L. Maturation of giardiavirus capsid protein involves posttranslational proteolytic processing by a cysteine protease. J. Virol. 1995, 69, 2825–2830. [Google Scholar] [CrossRef] [Green Version]
- Ghabrial, S.A.; Havens, W.M. The helminthosporium victoriae 190S mycovirus has two forms distinguishable by capsid protein composition and phosphorylation state. Virology 1992, 188, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Soldevila, A.I.; Webb, B.A.; Ghabrial, S.A. Expression, assembly, and proteolytic processing of helminthosporium victoriae 190S totivirus capsid protein in insect cells. Virology 1997, 234, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Ochoa, W.F.; Sinkovits, R.S.; Poulos, B.T.; Ghabrial, S.A.; Lightner, D.V.; Baker, T.S.; Nibert, M.L. Infectious myonecrosis virus has a totivirus-like, 120-subunit capsid, but with fiber complexes at the fivefold axes. Proc. Natl. Acad. Sci. USA 2008, 105, 17526–17531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S. Regulation of ribosomal proteins on viral Infection. Cells 2019, 8, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.M.; Selvam, S.; Fuchs, G. Fatal attraction: The roles of ribosomal proteins in the viral life cycle. WIREs RNA 2021, 12, e1613. [Google Scholar] [CrossRef] [PubMed]
- Bureau, M.; Leh, V.; Haas, M.; Geldreich, A.; Ryabova, L.; Yot, P.; Keller, M. P6 protein of Cauliflower mosaic virus, a translation reinitiator, interacts with ribosomal protein L13 from Arabidopsis thaliana. J. Gen. Virol. 2004, 85, 3765–3775. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.B.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P.B. Regulated nuclear trafficking of rpL10A mediated by NIK1 tepresents a defense strategy of plant cells against virus. PLoS Path. 2008, 4, e1000247. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone No. | Growth Media | NCBI or TAIR Description | TAIR Accession | Gene Ontology Information |
|---|---|---|---|---|
| 1 | QXA | Beta-1,4-N-acetylglucosaminyltransferase family protein | AT3G01620 | Located in Golgi apparatus |
| 2 | Zinc finger protein 2 | AT5G57520 | Located in nucleus | |
| 3 | DHHC-type zinc finger family protein | AT2G40990 | Is active in Golgi apparatus and endoplasmic reticulum | |
| 4 | Polynucleotide adenylyltransferase family protein | AT5G23690 | Involved in RNA processing | |
| 5 | Transmembrane protein | AT2G35750 | Located in mitochondrion | |
| 6 | Inorganic carbon transport protein-like protein (NdhL) | AT1G70760.1 | Located in chloroplast, chloroplast thylakoid membrane, thylakoid membrane | |
| 7 | QXA 2.5 mM | Cytochrome c oxidase assembly protein CtaG / Cox11 family | AT1G02410 | Located in chloroplast, integral component of mitochondrial membrane, mitochondrion |
| 8 | Pectin lyase-like superfamily protein | AT1G04680 | Located in extracellular region | |
| 9 | Plastid developmental protein DAG (MORF9) | AT1G11430 | Located in chloroplast, chloroplast envelope, chloroplast stroma | |
| 10 | Plant/protein | AT1G13990 | Located in chloroplast | |
| 11 | Peroxidase CB (PRXCB) | AT3G49120 | Located in Golgi apparatus, apoplast, cell wall, cytosol, extracellular region, plant-type cell wall, plant-type vacuole, secretory vesicle | |
| 12 | RmlC-like cupins superfamily protein | AT1G03890 | Located in extracellular region | |
| 13 | RNA polymerase transcriptional regulation mediator-like protein (MED6) | AT3G21350 | Located in nucleus | |
| 14 | Chloroplast ribosomal protein S3 | ATCG00800.1 | Located in chloroplast, chloroplast envelope, chloroplast nucleoid, chloroplast stroma, plastid | |
| 15 | Chloroplast GRX 12, GRXS12 | AT2G20270 | Located in chloroplast, chloroplast stroma, mitochondrion | |
| 16 | QXA 5 mM | DNAJ heat shock family protein | AT2G22360 | Located in chloroplast, chloroplast envelope, chloroplast thylakoid membrane, cytoplasm, vacuole |
| 17 | Ribosomal protein L17 family protein | AT3G54210 | Located in chloroplast, chloroplast envelope, chloroplast stroma, cytosol | |
| 18 | Sec14p-like phosphatidylinositol transfer family protein | AT1G72160 | Located in plasma membrane | |
| 19 | GDSL-like Lipase/Acylhydrolase superfamily protein | AT5G45670 | Located in extracellular region | |
| 20 | Chaperone protein dnaJ-like protein | AT5G06130 | Located in chloroplast membrane, mitochondrion | |
| 21 | GPI-anchored protein | AT3G18050 | Located in anchored component of membrane, chloroplast | |
| 22 | Pyrimidin 4 (PYR4) | AT4G22930 | Located in chloroplast, cytosol, mitochondrion | |
| 23 | Pectinacetylesterase family protein | AT4G19420 | Located in extracellular region | |
| 24 | Double Clp-N motif protein | AT4G12060 | Located in chloroplast, chloroplast envelope, chloroplast stroma, cytosol, plastid stroma | |
| 25 | PEBP (phosphatidylethanolamine-binding protein) family protein (FT) | AT1G65480 | Located in cytoplasm nucleus | |
| 26 | Clone RAFL09-89-G08 (R19778) putative cellulose synthase catalytic subunit (RSW1) | AT4G32410.1 | Located in Golgi apparatus, endosome, plasma membrane, trans-Golgi network | |
| 27 | mRNA for plastid protein, complete cds, clone: RAFL15-06-D14 | AT1G32580.1 | Located in chloroplast, mitochondrion, nucleus | |
| 28 | Papain family cysteine protease | AT4G16190 | Located in extracellular region, lytic vacuole, plant-type vacuole, vacuole |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurastoni, M.; Sá Antunes, T.F.; Abreu, E.F.M.; Ribeiro, S.G.; Mehta, A.; Sanches, M.M.; Fontes, W.; Kitajima, E.W.; Cruz, F.T.; Santos, A.M.C.; et al. A Capsid Protein Fragment of a Fusagra-like Virus Found in Carica papaya Latex Interacts with the 50S Ribosomal Protein L17. Viruses 2023, 15, 541. https://doi.org/10.3390/v15020541
Maurastoni M, Sá Antunes TF, Abreu EFM, Ribeiro SG, Mehta A, Sanches MM, Fontes W, Kitajima EW, Cruz FT, Santos AMC, et al. A Capsid Protein Fragment of a Fusagra-like Virus Found in Carica papaya Latex Interacts with the 50S Ribosomal Protein L17. Viruses. 2023; 15(2):541. https://doi.org/10.3390/v15020541
Chicago/Turabian StyleMaurastoni, Marlonni, Tathiana F. Sá Antunes, Emanuel F. M. Abreu, Simone G. Ribeiro, Angela Mehta, Marcio M. Sanches, Wagner Fontes, Elliot W. Kitajima, Fabiano T. Cruz, Alexandre M. C. Santos, and et al. 2023. "A Capsid Protein Fragment of a Fusagra-like Virus Found in Carica papaya Latex Interacts with the 50S Ribosomal Protein L17" Viruses 15, no. 2: 541. https://doi.org/10.3390/v15020541