
Bacteriophages as Biotechnological Tools
 , ,
, ,  and
and 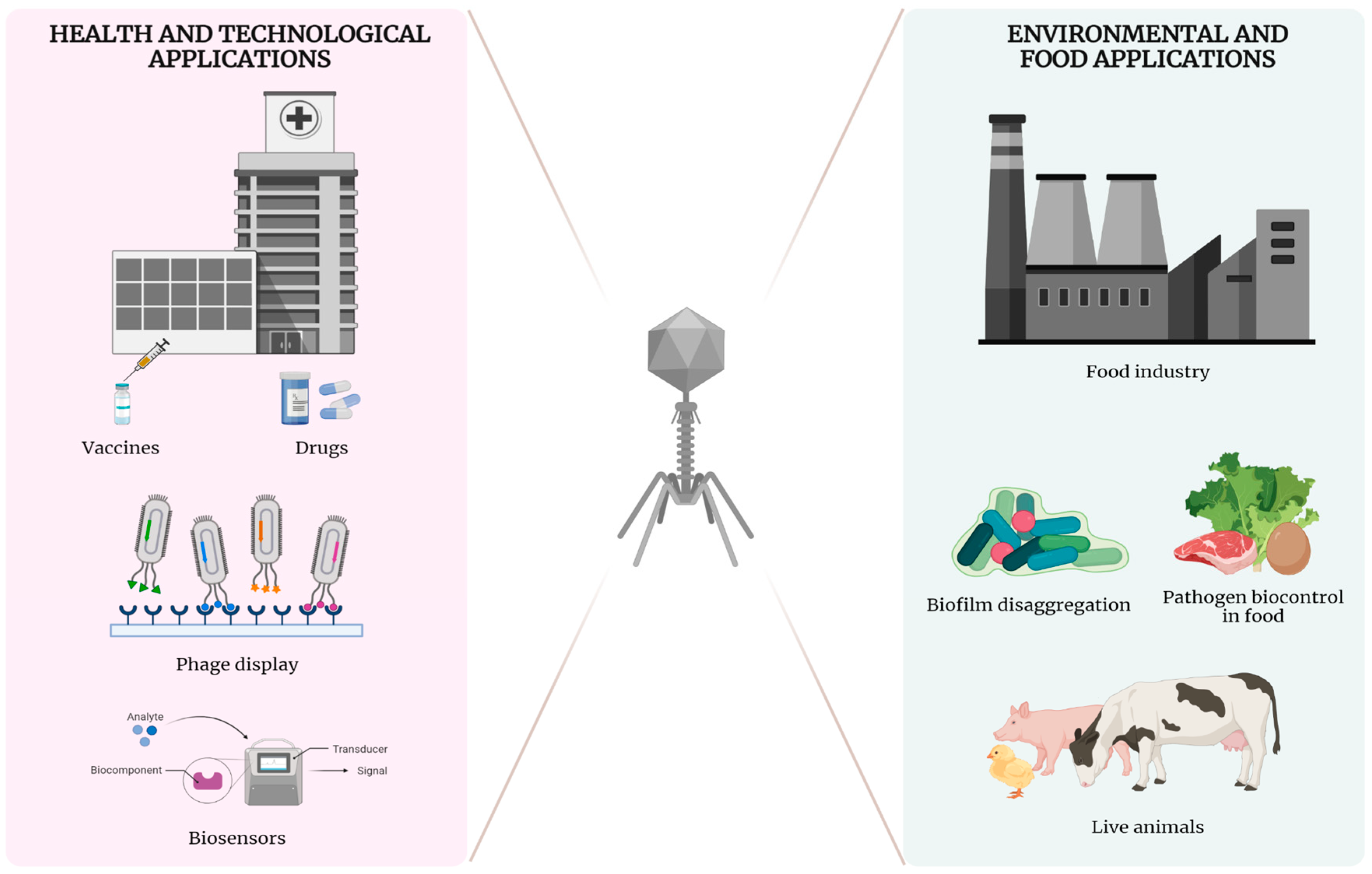{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Life Cycles of Bacteriophages
3. Mechanisms and Determinants of Host Range
4. Biotechnological Applications
4.1. Human Phage Therapy: Advantages and Limitations
Horizontal Gene Transfer (HGT) in Prokaryotes
4.2. Applications of Phages in Livestock and Food Industry
4.2.1. Applications in Live Animals (Animals Phage Therapy)
4.2.2. Pathogen Biocontrol in Food
4.2.3. Biofilm Disaggregation
Enzybiotics
4.2.4. Detection of Bacteria as Biosensor
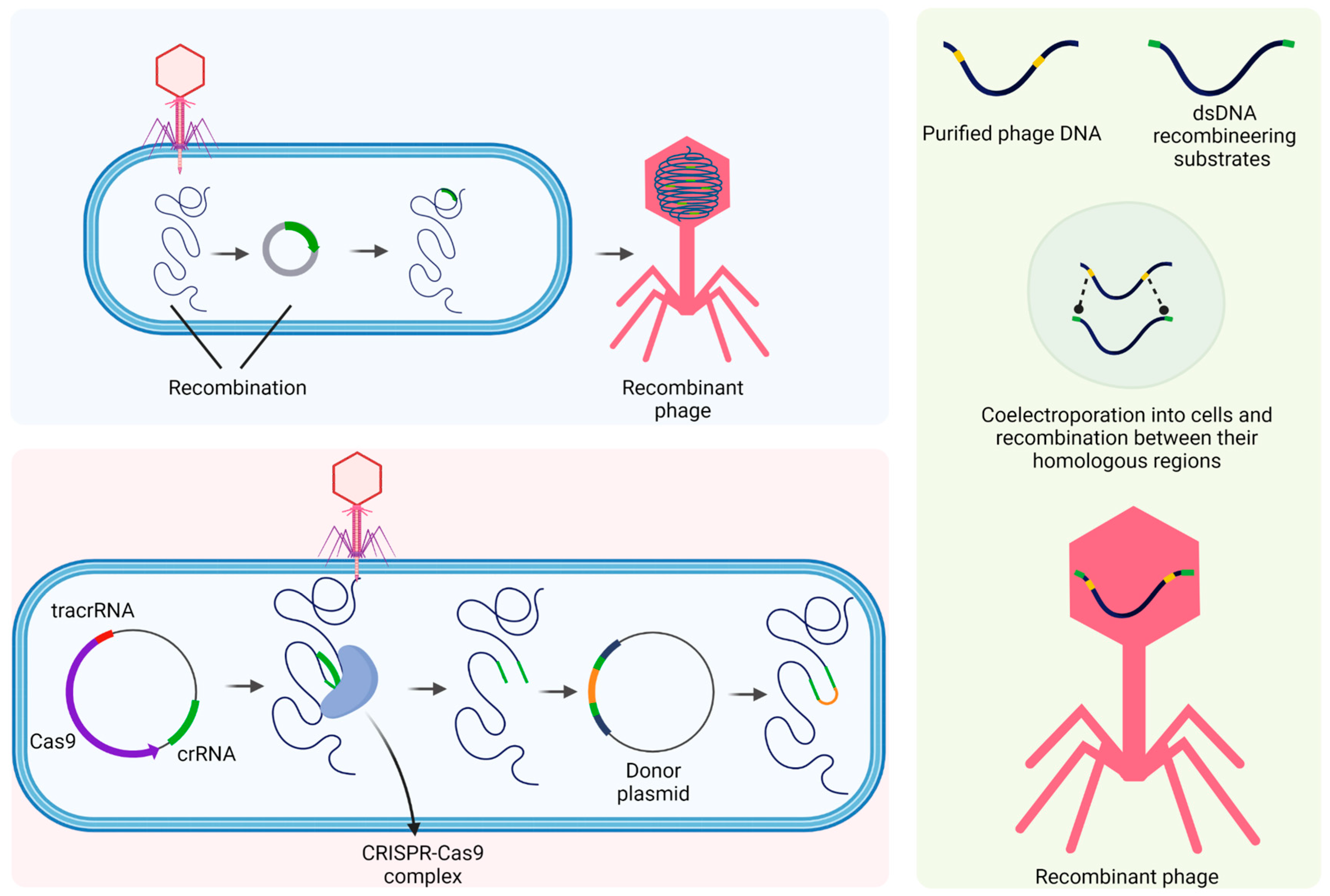
4.3. Bioengineered Phages
4.4. Phage Display
Phage Development for Vaccines
5. Final Considerations
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in Nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershey, A.D.; Chase, M. Independent Functions of Viral Protein and Nucleic Acid in Growth of Bacteriophage. J. Gen. Physiol. 1952, 36, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, N.; Roth, J.; Botstein, D. Genetic Engineering In Vivo Using Translocatable Drug-Resistance Elements. New Methods in Bacterial Genetics. J. Mol. Biol. 1977, 116, 125–159. [Google Scholar] [CrossRef] [PubMed]
- Wasylyk, B.; Derbyshire, R.; Guy, A.; Molko, D.; Roget, A.; Téoule, R.; Chambon, P. Specific in Vitro Transcription of Conalbumin Gene Is Drastically Decreased by Single-Point Mutation in T-A-T-A Box Homology Sequence. Proc. Natl. Acad. Sci. USA 1980, 77, 7024–7028. [Google Scholar] [CrossRef] [Green Version]
- Jacob, F.; Monod, J. Genetic Regulatory Mechanisms in the Synthesis of Proteins. J. Mol. Biol. 1961, 3, 318–356. [Google Scholar] [CrossRef]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; del Fiol, F.S.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef]
- Seed, K.D.; Lazinski, D.W.; Calderwood, S.B.; Camilli, A. A Bacteriophage Encodes Its Own CRISPR/Cas Adaptive Response to Evade Host Innate Immunity. Nature 2013, 494, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Sun, M.; Huang, D.; Zhang, Z.; Zhang, H.; Zhang, S.; Hu, F.; Jiang, X.; Jiao, W. A Review of Bacteriophage Therapy for Pathogenic Bacteria Inactivation in the Soil Environment. Environ. Int. 2019, 129, 488–496. [Google Scholar] [CrossRef]
- Burnet, F.M. The Classification of Dysentery-Coli Bacteriophages. III. A Correlation of the Serological Classification with Certain Biochemical Tests. J. Pathol. Bacteriol. 1933, 37, 179–184. [Google Scholar] [CrossRef]
- Ruska, H. Versuch Zu Einer Ordnung Der Virusarten. Arch. Virol. 1943, 2, 480–498. [Google Scholar] [CrossRef]
- Holmes, F.O. Order Virales. In Bergey’s Manual of Determinative Bacteriology, 6th ed.; Williams & Wilkins: Philadelphia, PA, USA, 1948; pp. 1126–1286. [Google Scholar]
- Lwoff, A.; Horne, R.; Tournier, P. A System of Viruses. Cold Spring Harb. Symp. Quant. Biol. 1962, 27, 51–55. [Google Scholar] [CrossRef]
- Bradley, D.E. Ultrastructure of Bacteriophage and Bacteriocins. Bacteriol. Rev. 1967, 31, 230–314. [Google Scholar] [CrossRef]
- Bradley, D.E. The Fluorescent Staining of Bacteriophage Nucleic Acids. J. Gen. Microbiol. 1966, 44, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Bradley, D.E. The Structure of Coliphages. J. Gen. Microbiol. 1963, 31, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, H.W.; Eisenstark, A. The Present State of Phage Taxonomy. Intervirology 1974, 3, 201–219. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage Diversity, Genomics and Phylogeny. Nat. Rev. 2020, 18, 125–138. [Google Scholar] [CrossRef]
- Lavigne, R.; Seto, D.; Mahadevan, P.; Ackermann, H.W.; Kropinski, A.M. Unifying Classical and Molecular Taxonomic Classification: Analysis of the Podoviridae Using BLASTP-Based Tools. Res. Microbiol. 2008, 159, 406–414. [Google Scholar] [CrossRef]
- Lavigne, R.; Darius, P.; Summer, E.J.; Seto, D.; Mahadevan, P.; Nilsson, A.S.; Ackermann, H.W.; Kropinski, A.M. Classification of Myoviridae Bacteriophages Using Protein Sequence Similarity. BMC Microbiol. 2009, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Edwards, R.; Nash, J.H.E.; Mahadevan, P.; Seto, D.; Ackermann, H.W.; Lavigne, R.; Kropinski, A.M. Integration of Genomic and Proteomic Analyses in the Classification of the Siphoviridae Family. Virology 2015, 477, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Paez-Espino, D.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Thomas, A.D.; Huntemann, M.; Mikhailova, N.; Rubin, E.; Ivanova, N.N.; Kyrpides, N.C. Uncovering Earth’s Virome. Nature 2016, 536, 425–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwer, F.; Edwards, R. The Phage Proteomic Tree: A Genome-Based Taxonomy for Phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, C.A. Marine Viruses—Major Players in the Global Ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.E.; Fuhrmann, J.J.; Wommack, K.E.; Radosevich, M. Viruses in Soil Ecosystems: An Unknown Quantity Within an Unexplored Territory. Annu. Rev. Virol. 2017, 4, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; McCartney, A.L.; Neve, H.; Gibson, G.R.; Sanderson, J.D.; Heller, K.J.; van Sinderen, D. Characterization of Virus-like Particles Associated with the Human Faecal and Caecal Microbiota. Res. Microbiol. 2014, 165, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Lwoff, A. Lysogeny. Bacteriol. Rev. 1953, 17, 269–337. [Google Scholar] [CrossRef]
- Hobbs, Z.; Abedon, S.T. Diversity of Phage Infection Types and Associated Terminology: The Problem with ‘Lytic or Lysogenic’. FEMS Microbiol. Lett. 2016, 363, 47. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Kotay, S.M.; Goel, R. Bacteriophage-Based Biocontrol of Biological Sludge Bulking in Wastewater. Bioeng. Bugs 2011, 2, 214–217. [Google Scholar] [CrossRef]
- Dennehy, J.J.; Abedon, S.T. Adsorption: Phage Acquisition of Bacteria. In Bacteriophages; Springer: Cham, Switzerland, 2020; pp. 1–25. [Google Scholar] [CrossRef]
- Lee, K.L.; Hubbard, L.C.; Hern, S.; Yildiz, I.; Gratzl, M.; Steinmetz, N.F. Shape Matters: The Diffusion Rates of TMV Rods and CPMV Icosahedrons in a Spheroid Model of Extracellular Matrix Are Distinct. Biomater. Sci. 2013, 1, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Saltzman, W.M.; Radomsky, M.L.; Whaley, K.J.; Cone, R.A. Antibody Diffusion in Human Cervical Mucus. Biophys. J. 1994, 66, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.L. Encounter Efficiency of Coliphage-Bacterium Interaction. Biochim. Biophys. Acta 1960, 39, 311–318. [Google Scholar] [CrossRef]
- Sisler, F.D. The Transmission of Bacteriophage by Mosquitoes. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 1940. [Google Scholar]
- Dennehy, J.J.; Friedenberg, N.A.; Yang, Y.W.; Turner, P.E. Bacteriophage Migration via Nematode Vectors: Host-Parasite-Consumer Interactions in Laboratory Microcosms. Appl. Environ. Microbiol. 2006, 72, 1974–1979. [Google Scholar] [CrossRef] [Green Version]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition Rates of Viruses and Bacteria above the Atmospheric Boundary Layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Forterre, P. The Virocell Concept and Environmental Microbiology. ISME J. 2012, 7, 233–236. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-Encoded Depolymerases: Their Diversity and Biotechnological Applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson’ And, S.; Taylor12, P.W. Neuraminidase Associated with Coliphage E That Specifically Depolymerizes the Escherichia Coli K1 Capsular Polysaccharide. J. Virol. 1985, 55, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Geyer, H.; Himmelspach, K.; Kwiatkowski, B.; Schlecht, S.; Stirm, S. Degradation of Bacterial Surface Carbohydrates by Virus-Associated Enzymes. Pure Appl. Chem. 1983, 55, 637–653. [Google Scholar] [CrossRef]
- Dennehy, J.J.; Abedon, S.T. Phage Infection and Lysis. In Bacteriophages; Springer: Cham, Switzerland, 2021; pp. 341–383. [Google Scholar] [CrossRef]
- Borodovich, T.; Shkoporov, A.N.; Ross, R.P.; Hill, C. Phage-Mediated Horizontal Gene Transfer and Its Implications for the Human Gut Microbiome. Gastroenterol. Rep. 2022, 10, goac012. [Google Scholar] [CrossRef]
- Dowah, A.S.A.; Clokie, M.R.J. Review of the Nature, Diversity and Structure of Bacteriophage Receptor Binding Proteins That Target Gram-Positive Bacteria. Biophys. Rev. 2018, 10, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Arutyunov, D.; Foss, M.; Cunningham, A.; Ding, W.; Singh, A.; Pavlov, A.R.; Henry, M.; Evoy, S.; Kelly, J.; et al. Genome and Proteome of Campylobacter Jejuni Bacteriophage NCTC 12673. Appl. Environ. Microbiol. 2011, 77, 8265–8271. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.J. The Discovery and Application of Bacteriophage Receptor Binding Proteins. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2016. [Google Scholar]
- Hu, B.; Margolin, W.; Molineux, I.J.; Liu, J. Structural Remodeling of Bacteriophage T4 and Host Membranes during Infection Initiation. Proc. Natl. Acad. Sci. USA 2015, 112, E4919–E4928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.B.; Storms, Z.; Sauvageau, D. Host Receptors for Bacteriophage Adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tétart, F.; Repoila, F.; Monod, C.; Krisch, H.M. Bacteriophage T4 Host Range Is Expanded by Duplications of a Small Domain of the Tail Fiber Adhesin. J. Mol. Biol. 1996, 258, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Mosig, G.; Eiserling, F.E. T4 and Related Phages: Structure and Development. In The Bacteriophages, 2nd ed.; Oxford University Press: New York, NY, USA, 2006; pp. 1–131. [Google Scholar]
- Sun, Y.; Roznowski, A.P.; Tokuda, J.M.; Klose, T.; Mauney, A.; Pollack, L.; Fane, B.A.; Rossmann, M.G. Structural Changes of Tailless Bacteriophage Φx174 during Penetration of Bacterial Cell Walls. Proc. Natl. Acad. Sci. USA 2017, 114, 13708–13713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, F.; Air, G.M.; Barrell, B.G.; Brown, N.L.; Coulson, A.R.; Fiddes, J.C.; Hutchison, C.A.; Slocombe, P.M.; Smith, M. Nucleotide Sequence of Bacteriophage Phi X174 DNA. Nature 1977, 265, 687–695. [Google Scholar] [CrossRef]
- Liu, M.; Deora, R.; Doulatov, S.R.; Gingery, M.; Eiserling, F.A.; Preston, A.; Maskell, D.J.; Simons, R.W.; Cotter, P.A.; Parkhill, J.; et al. Reverse Transcriptase-Mediated Tropism Switching in Bordetella Bacteriophage. Science (1979) 2002, 295, 2091–2094. [Google Scholar] [CrossRef]
- Schwarzer, D.; Buettner, F.F.R.; Browning, C.; Nazarov, S.; Rabsch, W.; Bethe, A.; Oberbeck, A.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; et al. A Multivalent Adsorption Apparatus Explains the Broad Host Range of Phage Phi92: A Comprehensive Genomic and Structural Analysis. J. Virol. 2012, 86, 10384–10398. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Lemire, S.; Pires, D.P.; Lu, T.K. Engineering Modular Viral Scaffolds for Targeted Bacterial Population Editing. Cell Syst. 2015, 1, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Zhang, L.; Abdelgader, S.A.; Yu, L.; Xu, J.; Yao, H.; Lu, C.; Zhang, W. Alterations in Gp37 Expand the Host Range of a T4-like Phage. Appl. Environ. Microbiol. 2017, 83, e01576-17. [Google Scholar] [CrossRef] [Green Version]
- Duplessis, M.; Moineau, S. Identification of a Genetic Determinant Responsible for Host Specificity in Streptococcus Thermophilus Bacteriophages. Mol. Microbiol. 2001, 41, 325–3366. [Google Scholar] [CrossRef]
- Marzari, R.; Sblattero, D.; Righi, M.; Bradbury, A. Extending Filamentous Phage Host Range by the Grafting of a Heterologous Receptor Binding Domain. Gene 1997, 185, 27–33. [Google Scholar] [CrossRef]
- Dams, D.; Brøndsted, L.; Drulis-Kawa, Z.; Briers, Y. Engineering of Receptor-Binding Proteins in Bacteriophages and Phage Tail-like Bacteriocins. Biochem. Soc. Trans. 2019, 47, 449–460. [Google Scholar] [CrossRef]
- Moller, A.G.; Lindsay, J.A.; Read, T.D. Determinants of Phage Host Range in Staphylococcus Species. Appl. Environ. Microbiol. 2019, 85, e00209-19. [Google Scholar] [CrossRef] [Green Version]
- Mavrich, T.N.; Hatfull, G.F. Evolution of Superinfection Immunity in Cluster A Mycobacteriophages. mBio 2019, 10, e00971-19. [Google Scholar] [CrossRef] [Green Version]
- Berngruber, T.W.; Weissing, F.J.; Gandon, S. Inhibition of Superinfection and the Evolution of Viral Latency. J. Virol. 2010, 84, 10200–10208. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.G.; Murray, N.E. Restriction and Modification Systems. Annu. Rev. Genet. 1991, 25, 585–627. [Google Scholar] [CrossRef]
- Barrangou, R.; Marraffini, L.A. CRISPR-Cas Systems: Prokaryotes Upgrade to Adaptive Immunity. Mol. Cell 2014, 54, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Hryhorowicz, M.; Lipiński, D.; Zeyland, J.; Słomski, R. CRISPR/Cas9 Immune System as a Tool for Genome Engineering. Arch. Immunol. Ther. Exp. (Warsz) 2017, 65, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Rath, D.; Amlinger, L.; Rath, A.; Lundgren, M. The CRISPR-Cas Immune System: Biology, Mechanisms and Applications. Biochimie 2015, 117, 119–128. [Google Scholar] [CrossRef]
- Edgar, R.; Friedman, N.; Shahar, M.M.; Qimron, U. Reversing Bacterial Resistance to Antibiotics by Phage-Mediated Delivery of Dominant Sensitive Genes. Appl. Environ. Microbiol. 2012, 78, 744–751. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage Endolysins as Novel Antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, H.J.; Shield, C.G.; Swift, B.M.C. The Application of Bacteriophage Diagnostics for Bacterial Pathogens in the Agricultural Supply Chain: From Farm-To-Fork. PHAGE Ther. Appl. Res. 2020, 1, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Nakayinga, R.; Makumi, A.; Tumuhaise, V.; Tinzaara, W. Xanthomonas Bacteriophages: A Review of Their Biology and Biocontrol Applications in Agriculture. BMC Microbiol. 2021, 21, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, R.; Miyashita, S.; Ando, S.; Ito, K.; Fukuhara, T.; Takahashi, H. Isolation and Characterization of a Novel Jumbo Phage from Leaf Litter Compost and Its Suppressive Effect on Rice Seedling Rot Diseases. Viruses 2021, 13, 591. [Google Scholar] [CrossRef] [PubMed]
- Cristobal-Cueto, P.; García-Quintanilla, A.; Esteban, J.; García-Quintanilla, M. Phages in Food Industry Biocontrol and Bioremediation. Antibiotics 2021, 10, 786. [Google Scholar] [CrossRef]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M.; Sip, A. Effectiveness of Phage-Based Inhibition of Listeria Monocytogenes in Food Products and Food Processing Environments. Microorganisms 2020, 8, 1764. [Google Scholar] [CrossRef]
- Drulis-Kawa, Z.; Majkowska-Skrobek, G.; Maciejewska, B. Bacteriophages and Phage-Derived Proteins-Application Approaches. Curr. Med. Chem. 2015, 22, 1757–1773. [Google Scholar] [CrossRef]
- Principi, N.; Silvestri, E.; Esposito, S. Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections. Front. Pharmacol. 2019, 10, 513. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.B.; Azeredo, J. Bacteriophage-Based Biotechnological Applications. Viruses 2019, 11, 737. [Google Scholar] [CrossRef] [Green Version]
- Summers, W.C. Cholera and Plague in India: The Bacteriophage Inquiry of 1927–1936. J. Hist. Med. 1993, 48, 275–301. [Google Scholar] [CrossRef] [Green Version]
- Abul-Hassan, H.S.; El-Tahan k Massoud, B.; Gomaa, R. Bacteriophage Therapy of Pseudomonas Burn Wound Sepsis. Ann. MBC 1990, 3, 262–264. [Google Scholar]
- Essoh, C.; Blouin, Y.; Loukou, G.; Cablanmian, A.; Lathro, S.; Kutter, E.; Thien, V.H.; Vergnaud, G.; Pourcel, C. The Susceptibility of Pseudomonas Aeruginosa Strains from Cystic Fibrosis Patients to Bacteriophages. PLoS ONE 2013, 8, e60575. [Google Scholar] [CrossRef] [Green Version]
- Hoyle, N.; Zhvaniya, P.; Balarjishvili, N.; Bolkvadze, D.; Nadareishvili, L.; Nizharadze, D.; Wittmann, J.; Rohde, C.; Kutateladze, M. Phage Therapy against Achromobacter Xylosoxidans Lung Infection in a Patient with Cystic Fibrosis: A Case Report. Res. Microbiol. 2018, 169, 540–542. [Google Scholar] [CrossRef]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; le Floch, R.; et al. Efficacy and Tolerability of a Cocktail of Bacteriophages to Treat Burn Wounds Infected by Pseudomonas Aeruginosa (PhagoBurn): A Randomised, Controlled, Double-Blind Phase 1/2 Trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Kvachadze, L.; Balarjishvili, N.; Meskhi, T.; Tevdoradze, E.; Skhirtladze, N.; Pataridze, T.; Adamia, R.; Topuria, T.; Kutter, E.; Rohde, C.; et al. Evaluation of Lytic Activity of Staphylococcal Bacteriophage Sb-1 against Freshly Isolated Clinical Pathogens. Microb. Biotechnol. 2011, 4, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Markoishvili, K.; Tsitlanadze, G.; Katsarava, R.; Glenn Morris, J.; Sulakvelidze, A.; Alexander Sulakvelidze, C. Pharmacology and Therapeutics A Novel Sustained-Release Matrix Based on Biodegradable Poly(Ester Amide)s and Impregnated with Bacteriophages and an Antibiotic Shows Promise in Management of Infected Venous Stasis Ulcers and Other Poorly Healing Wounds. Int. J. Dermatol. 2002, 41, 453–458. [Google Scholar] [CrossRef]
- Rhoads, D.D.; Wolcott, R.D.; Kuskowski, M.A.; Wolcott, B.M.; Ward, L.S.; Sulakvelidze, A. Bacteriophage Therapy of Venous Leg Ulcers in Humans: Results of a Phase I Safety Trial. J. Wound Care 2009, 18, 237–243. [Google Scholar] [CrossRef]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage Adhering to Mucus Provide a Non-Host-Derived Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef] [Green Version]
- Barr, J.J.; Auro, R.; Sam-Soon, N.; Kassegne, S.; Peters, G.; Bonilla, N.; Hatay, M.; Mourtada, S.; Bailey, B.; Youle, M.; et al. Subdiffusive Motion of Bacteriophage in Mucosal Surfaces Increases the Frequency of Bacterial Encounters. Proc. Natl. Acad. Sci. USA 2015, 112, 13675–13680. [Google Scholar] [CrossRef] [Green Version]
- Guerin, E.; Hill, C. Shining Light on Human Gut Bacteriophages. Front. Cell. Infect. Microbiol. 2020, 10, 481. [Google Scholar] [CrossRef]
- An, T.W.; Kim, S.J.; Lee, Y.D.; Park, J.H.; Chang, H.I. The Immune-Enhancing Effect of the Cronobacter Sakazakii ES2 Phage Results in the Activation of Nuclear Factor-ΚB and Dendritic Cell Maturation via the Activation of IL-12p40 in the Mouse Bone Marrow. Immunol. Lett. 2014, 157, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the Host Immune System and Bacteriophage Is Essential for Successful Phage Therapy against an Acute Respiratory Pathogen. Cell Host Microbe 2017, 22, 38–47.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Li, C.; Xia, S.; Tian, X.; Kong, Y.; Wang, Z.; Gu, C.; Zhang, R.; Tu, C.; Xie, Y.; et al. Identification of Human Single-Domain Antibodies against SARS-CoV-2. Cell Host Microbe 2020, 27, 891–898.e5. [Google Scholar] [CrossRef] [PubMed]
- Dor-On, E.; Solomon, B. Targeting Glioblastoma via Intranasal Administration of Ff Bacteriophages. Front. Microbiol. 2015, 6, 530. [Google Scholar] [CrossRef] [PubMed]
- Ghasemian, A.; Moradpour, Z.; Yousefi, N.; Sadeghi, D.; Synergistic, G.A. Spectroscopic Investigation on the Interaction of DNA with Superparamagnetic Iron Oxide Nanoparticles Doped with Chromene via Dopamine as Cross Linker. Iran J. Basic Med. Sci. 2020, 23, 257–263. [Google Scholar] [CrossRef]
- Bao, J.; Wu, N.; Zeng, Y.; Chen, L.; Li, L.; Yang, L.; Zhang, Y.; Guo, M.; Li, L.; Li, J.; et al. Non-Active Antibiotic and Bacteriophage Synergism to Successfully Treat Recurrent Urinary Tract Infection Caused by Extensively Drug-Resistant Klebsiella Pneumoniae. Emerg. Microbes Infect. 2020, 9, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Da̧browska, K.; Opolski, A.; Wietrzyk, J.; Nevozhay, D.; Szczaurska, K.; Świtała-Jeleń, K.; Boratyński, J.; Górski, A. Activity of Bacteriophages in Murine Tumor Models Depends on the Route of Phage Administration. Oncol. Res. 2005, 15, 183–187. [Google Scholar] [CrossRef]
- Eriksson, F.; Culp, W.D.; Massey, R.; Egevad, L.; Garland, D.; Persson, M.A.A.; Pisa, P. Tumor Specific Phage Particles Promote Tumor Regression in a Mouse Melanoma Model. Cancer Immunol. Immunother. 2007, 56, 677–687. [Google Scholar] [CrossRef]
- Sanmukh, S.G.; Santos, S.; Felisbino, S.L. Natural Bacteriophages T4 and M13 Down-Regulates Hsp90 Gene Expression in Human Prostate Cancer Cells (PC-3) Representing a Potential Nanoparticle against Cancer. Characterization of Bacteriophages and Prostate Cancer Cell Interaction View Project Bone Marrow Mesenchymal Stem Cells and Periodontal Regeneration View Project. Virol. Res. J. 2017, 1, 21–23. [Google Scholar]
- Abedon, S.T.; Thomas-Abedon, C. Phage Therapy Pharmacology. Curr. Pharm. Biotechnol. 2010, 11, 28–47. [Google Scholar] [CrossRef]
- Tetz, G.V.; Ruggles, K.V.; Zhou, H.; Heguy, A.; Tsirigos, A.; Tetz, V. Bacteriophages as Potential New Mammalian Pathogens. Sci. Rep. 2017, 7, 7043. [Google Scholar] [CrossRef] [Green Version]
- Górski, A.; Borysowski, J.; Międzybrodzki, R.; Weber-Dąbrowska, B. Bacteriophages in Medicine. Bacteriophage Genet. Mol. Biol. 2007, 125–158. [Google Scholar]
- Sulakvelidze, A. Editorial. Drug Discov. Today 2005, 12, 807–809. [Google Scholar] [CrossRef]
- Fortier, L.; Sekulovic, O. Importance of Prophages to Evolution and Virulence of Bacterial Pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and Cons of Phage Therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [Green Version]
- McDougald, D.; Rice, S.A.; Barraud, N.; Steinberg, P.D.; Kjelleberg, S. Should We Stay or Should We Go: Mechanisms and Ecological Consequences for Biofilm Dispersal. Nat. Rev. Microbiol. 2012, 10, 39–50. [Google Scholar] [CrossRef]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a Post-Antibiotic Era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Kifelew, L.G.; Warner, M.S.; Morales, S.; Thomas, N.; Gordon, D.L.; Mitchell, J.G.; Speck, P.G. Efficacy of Lytic Phage Cocktails on Staphylococcus Aureus and Pseudomonas Aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses 2020, 12, 559. [Google Scholar] [CrossRef]
- Furfaro, L.L.; Payne, M.S.; Chang, B.J. Bacteriophage Therapy: Clinical Trials and Regulatory Hurdles. Front. Cell Infect. Microbiol. 2018, 8, 376. [Google Scholar] [CrossRef] [Green Version]
- Parracho, H.M.; Burrowes, B.H.; Enright, M. The Role of Regulated Clinical Trials in the Development of Bacteriophage Therapeutics. J. Mol. Genet. Med. 2012, 6, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.L. Bacteriophage-Mediated Horizontal Gene Transfer: Transduction. In Bacteriophages; Springer: Cham, Switzerland, 2021; pp. 151–192. [Google Scholar] [CrossRef]
- Corpuz, A.V. Potential of Bacteriophage Therapy in Treating Hospital Wastewater. Eur. J. Mol. Clin. Med. 2020, 7, 2020. [Google Scholar]
- Ballesté, E.; Blanch, A.R.; Muniesa, M.; García-Aljaro, C.; Rodríguez-Rubio, L.; Martín-Díaz, J.; Pascual-Benito, M.; Jofre, J. Bacteriophages in Sewage: Abundance, Roles, and Applications. FEMS Microbes 2022, 3, xtac009. [Google Scholar] [CrossRef]
- Guo, Z.; Lin, H.; Ji, X.; Yan, G.; Lei, L.; Han, W.; Gu, J.; Huang, J. Therapeutic Applications of Lytic Phages in Human Medicine. Microb. Pathog. 2020, 142, 104048. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Huiwen, M.; Wang, J.; Wang, Y.; Khan, S.A.; Zhang, Y.; Qiu, H.; Jiang, L.; He, L.; Zhang, Y.; et al. A Novel Polymerase β Inhibitor from Phage Displayed Peptide Library Augments the Anti-Tumour Effects of Temozolomide on Colorectal Cancer. J. Chemother. 2021, 34, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Liang, L.; Yin, P.; Zhou, Y.; Sharoba, A.M.; Lu, Q.; Dong, X.; Liu, K.; Connerton, I.F.; Li, J. Application of a Novel Phage LPSEYT for Biological Control of Salmonella in Foods. Microorganisms 2020, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Waturangi, D.E.; Kasriady, C.P.; Guntama, G.; Sahulata, A.M.; Lestari, D.; Magdalena, S. Application of Bacteriophage as Food Preservative to Control Enteropathogenic Escherichia Coli (EPEC). BMC Res. Notes 2021, 14, 336. [Google Scholar] [CrossRef]
- Islam, M.S.; Yang, X.; Euler, C.W.; Han, X.; Liu, J.; Hossen, M.I.; Zhou, Y.; Li, J. Application of a Novel Phage ZPAH7 for Controlling Multidrug-Resistant Aeromonas Hydrophila on Lettuce and Reducing Biofilms. Food Control 2021, 122, 107785. [Google Scholar] [CrossRef]
- Endersen, L.; Coffey, A. The Use of Bacteriophages for Food Safety. Curr. Opin. Food Sci. 2020, 36, 1–8. [Google Scholar] [CrossRef]
- Cooper, I.R. A Review of Current Methods Using Bacteriophages in Live Animals, Food and Animal Products Intended for Human Consumption. J. Microbiol. Methods 2016, 130, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, V.; Baquero, D.; Hernandez, S.; Farfan, J.C.; Arias, J.; Arévalo, A.; Donado-Godoy, P.; Vives-Flores, M. Phage Cocktail SalmoFREE® Reduces Salmonella on a Commercial Broiler Farm. Poult. Sci. 2019, 98, 5054–5063. [Google Scholar] [CrossRef]
- Ma, Y.; Pacan, J.C.; Wang, Q.; Sabour, P.M.; Huang, X.; Xu, Y. Enhanced Alginate Microspheres as Means of Oral Delivery of Bacteriophage for Reducing Staphylococcus Aureus Intestinal Carriage. Food Hydrocoll. 2012, 26, 434–440. [Google Scholar] [CrossRef]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M.J. Virulent Bacteriophage for Efficient Biocontrol of Listeria Monocytogenes in Ready-to-Eat Foods. Appl. Environ. Microbiol. 2009, 75, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Guenther, S.; Loessner, M.J. Bacteriophage Biocontrol of Listeria Monocytogenes on Soft Ripened White Mold and Red-Smear Cheeses. Bacteriophage 2011, 1, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Hudson, J.A.; Billington, C.; Cornelius, A.J.; Wilson, T.; On, S.L.W.; Premaratne, A.; King, N.J. Use of a Bacteriophage to Inactivate Escherichia coli O157:H7 on Beef. Food Microbiol. 2013, 36, 14–21. [Google Scholar] [CrossRef]
- Hungaro, H.M.; Mendonça, R.C.S.; Gouvêa, D.M.; Vanetti, M.C.D.; Pinto, C.L.d.O. Use of Bacteriophages to Reduce Salmonella in Chicken Skin in Comparison with Chemical Agents. Food Res. Int. 2013, 52, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Liu, Y.; Peng, L.; Cai, X.; Shen, L.; Duan, M.; Ning, Y.; Liu, S.; Li, C.; Liu, Y.; et al. Characterization of the Narrow-Spectrum Bacteriophage LSE7621 towards Salmonella Enteritidis and Its Biocontrol Potential on Lettuce and Tofu. LWT 2020, 118, 108791. [Google Scholar] [CrossRef]
- Wong, C.W.Y.; Delaquis, P.; Goodridge, L.; Lévesque, R.C.; Fong, K.; Wang, S. Inactivation of Salmonella Enterica on Post-Harvest Cantaloupe and Lettuce by a Lytic Bacteriophage Cocktail. Curr. Res. Food Sci. 2019, 2, 25–32. [Google Scholar] [CrossRef]
- García, P.; Rodríguez, L.; Rodríguez, A.; Martínez, B. Food Biopreservation: Promising Strategies Using Bacteriocins, Bacteriophages and Endolysins. Trends Food Sci. Technol. 2010, 21, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, H.W.; Greer, G.G.; Rocourt, J. Morphology of Brochothrix Thermosphacta Phages. Microbios 1988, 56, 19–26. [Google Scholar]
- Hudson, J.A.; Billington, C.; Carey-Smith, G.; Greening, G. Bacteriophages as Biocontrol Agents in Food. J. Food Prot. 2005, 68, 426–437. [Google Scholar] [CrossRef]
- Schmelcher, M.; Loessner, M.J. Application of Bacteriophages for Detection of Foodborne Pathogens. Bacteriophage 2014, 4, e28137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Niu, Y.D.; Nan, Y.; Stanford, K.; Holley, R.; McAllister, T.; Narváez-Bravo, C. SalmoFreshTM Effectiveness in Controlling Salmonella on Romaine Lettuce, Mung Bean Sprouts and Seeds. Int. J. Food Microbiol. 2019, 305, 108250. [Google Scholar] [CrossRef] [PubMed]
- Magnone, J.P.; Marek, P.J.; Sulakvelidze, A.; Senecal, A.G. Additive Approach for Inactivation of Escherichia Coli O157:H7, Salmonella, and Shigella Spp. on Contaminated Fresh Fruits and Vegetables Using Bacteriophage Cocktail and Produce Wash. J. Food Prot. 2013, 76, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dashiell, G.; Handy, E.T.; East, C.; Reynnells, R.; White, C.; Nyarko, E.; Micallef, S.; Hashem, F.; Millner, P.D. Survival of Salmonella Newport on Whole and Fresh-Cut Cucumbers Treated with Lytic Bacteriophages. J. Food Prot. 2017, 80, 668–673. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on Chicken Breast Fillets Stored under Aerobic or Modified Atmosphere Packaging by the Application of Lytic Bacteriophage Preparation SalmoFresh. Poult. Sci. 2016, 95, 668–675. [Google Scholar] [CrossRef]
- Vikram, A.; Tokman, J.I.; Woolston, J.; Sulakvelidze, A. Phage Biocontrol Improves Food Safety by Significantly Reducing the Level and Prevalence of Escherichia Coli O157:H7 in Various Foods. J. Food Prot. 2020, 83, 668–676. [Google Scholar] [CrossRef]
- Perera, M.N.; Abuladze, T.; Li, M.; Woolston, J.; Sulakvelidze, A. Bacteriophage Cocktail Significantly Reduces or Eliminates Listeria Monocytogenes Contamination on Lettuce, Apples, Cheese, Smoked Salmon and Frozen Foods. Food Microbiol. 2015, 52, 42–48. [Google Scholar] [CrossRef]
- Food Safety Authority, E. Evaluation of the Safety and Efficacy of ListexTM P100 for Reduction of Pathogens on Different Ready-to-Eat (RTE) Food Products. EFSA J. 2016, 14, e04565. [Google Scholar] [CrossRef]
- Zoonoses. Available online: https://www.who.int/news-room/fact-sheets/detail/zoonoses (accessed on 5 January 2023).
- Gambino, M.; Brøndsted, L. Looking into the Future of Phage-Based Control of Zoonotic Pathogens in Food and Animal Production. Curr. Opin. Biotechnol. 2021, 68, 96–103. [Google Scholar] [CrossRef]
- Lewis, R.; Hill, C. Overcoming Barriers to Phage Application in Food and Feed. Curr. Opin. Biotechnol. 2020, 61, 38–44. [Google Scholar] [CrossRef]
- Gigante, A.; Atterbury, R.J. Veterinary Use of Bacteriophage Therapy in Intensively-Reared Livestock. Virol. J. 2019, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Cortés, M.E.; Consuegra Bonilla, J.; Sinisterra, R.D. Biofilm Formation, Control and Novel Strategies for Eradication. Sci. Against Microb. Pathog. Commun. Curr. Res. Technol. Adv. 2011, 2, 896–905. [Google Scholar]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Flemming, H.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Satpathy, S.; Sen, S.K.; Pattanaik, S.; Raut, S. Review on Bacterial Biofilm: An Universal Cause of Contamination. Biocatal. Agric. Biotechnol. 2016, 7, 56–66. [Google Scholar] [CrossRef]
- Halan, B.; Buehler, K.; Schmid, A. Biofilms as Living Catalysts in Continuous Chemical Syntheses. Trends Biotechnol. 2012, 30, 453–465. [Google Scholar] [CrossRef]
- Bordi, C.; de Bentzmann, S. Hacking into Bacterial Biofilms: A New Therapeutic Challenge. Ann. Intensive Care 2011, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shirtliff, M.; Leid, J.G. The Role of Biofilms in Device-Related Infections; Shirtliff, M., Leid, J.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 3, ISBN 978-3-540-68113-7. [Google Scholar]
- Harper, D.R.; Parracho, H.M.R.T.; Walker, J.; Sharp, R.; Hughes, G.; Werthén, M.; Lehman, S.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270. [Google Scholar] [CrossRef]
- Cornelissen, A.; Ceyssens, P.; T’Syen, J.; Van Praet, H.; Noben, J.-P.; Shaburova, O.V.; Krylov, V.N.; Volckaert, G.; Lavigne, R. The T7-Related Pseudomonas Putida Phage Φ15 Displays Virion-Associated Biofilm Degradation Properties. PLoS ONE 2011, 6, e18597. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, G.W.; Denyer, S.P.; Olliff, C.J.; Ibrahim, L.J. Reduction in Exopolysaccharide Viscosity as an Aid to Bacteriophage Penetration through Pseudomonas Aeruginosa Biofilms. Appl. Environ. Microbiol. 2001, 67, 2746–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmer, D.B.; Schmitz, J.E.; Euler, C.W.; Fischetti, V.A. Novel Bacteriophage Lysin with Broad Lytic Activity Protects against Mixed Infection by Streptococcus Pyogenes and Methicillin-Resistant Staphylococcus Aureus. Antimicrob. Agents Chemother. 2013, 57, 2743–2750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, U.; Vipra, A.; Channabasappa, S. Phage-Derived Lysins as Potential Agents for Eradicating Biofilms and Persisters. Drug Discov. Today 2018, 23, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, R.; García, E.; García, P. Phage Lysins for Fighting Bacterial Respiratory Infections: A New Generation of Antimicrobials. Front. Immunol. 2018, 9, 2252. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and Elimination of Upper Respiratory Colonization of Mice by Group A Streptococci by Using a Bacteriophage Lytic Enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [Green Version]
- Bamford, D.H.; Romantschuk, M.; Somerharju, P.J. Membrane Fusion in Prokaryotes: Bacteriophage Phi 6 Membrane Fuses with the Pseudomonas Syringae Outer Membrane. EMBO J. 1987, 6, 1467–1473. [Google Scholar] [CrossRef]
- Caldentey, J.; Bamford, D.H. The Lytic Enzyme of the Pseudomonas Phage B6. Purification and Biochemical Characterization. Biochim. Biophys. Acta (BBA)—Protein Struct. Mol. Enzymol. 1992, 1159, 44–50. [Google Scholar] [CrossRef]
- Mindich, L.; Lehman, J. Cell Wall Lysin as a Component of the Bacteriophage C6 Virion. J. Virol. 1979, 30, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Romantschuk, M.; Olkkonen, V.M.; Bamford, D.H. The Nucleocapsid of Bacteriophage 06 Penetrates the Host Cytoplasmic Membrane. EMBO J. 1988, 7, 1821–1829. [Google Scholar] [CrossRef]
- Arisaka, F.; Kanamaru, S.; Leiman, P.; Rossmann, M.G. The Tail Lysozyme Complex of Bacteriophage T4. Int. J. Biochem. Cell Biol. 2003, 35, 16–21. [Google Scholar] [CrossRef]
- Moak, M.; Molineux, I.J. Role of the Gp16 Lytic Transglycosylase Motif in Bacteriophage T7 Virions at the Initiation of Infection. Mol. Microbiol. 2000, 37, 345–355. [Google Scholar] [CrossRef]
- Pritchard, D.G.; Dong, S.; Kirk, M.C.; Cartee, R.T.; Baker, J.R. LambdaSa1 and LambdaSa2 Prophage Lysins of Streptococcus Agalactiae. Appl. Environ. Microbiol. 2007, 73, 7150–7154. [Google Scholar] [CrossRef] [Green Version]
- Stockdale, S.R.; Mahony, J.; Courtin, P.; Chapot-Chartier, M.; van Pijkeren, J.; Britton, R.A.; Neve, H.; Heller, K.J.; Aideh, B.; Vogensen, F.K.; et al. The Lactococcal Phages Tuc2009 and TP901-1 Incorporate Two Alternate Forms of Their Tail Fiber into Their Virions for Infection Specialization. J. Biol. Chem. 2013, 288, 5581–5590. [Google Scholar] [CrossRef] [Green Version]
- Castillo, F.J.; Bartell, P.F. Localization and Functional Role of the Pseudomonas Bacteriophage 2 Depolymerase. J. Virol. 1976, 18, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.A.; Sutherland, I.W.; Jones, M.V. Biofilm Susceptibility to Bacteriophage Attack: The Role of Phage-Borne Polysaccharide Depolymerase. Microbiology (Reading) 1998, 144 Pt 11, 3039–3047. [Google Scholar] [CrossRef] [Green Version]
- Linnerborg, M.; Weintraub, A.; Albert, M.J.; Widmalm, G. Depolymerization of the Capsular Polysaccharide from Vibrio Cholerae O139 by a Lyase Associated with the Bacteriophage JA1. Carbohydr. Res. 2001, 333, 263–269. [Google Scholar] [CrossRef]
- Pelkonen, S.; Aalto, J.; Finne, J. Differential Activities of Bacteriophage Depolymerase on Bacterial Polysaccharide: Binding Is Essential but Degradation Is Inhibitory in Phage Infection of K1-Defective Escherichia Coli. J. Bacteriol. 1992, 174, 7757–7761. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, A.; Ceyssens, P.J.; Krylov, V.N.; Noben, J.P.; Volckaert, G.; Lavigne, R. Identification of EPS-Degrading Activity within the Tail Spikes of the Novel Pseudomonas Putida Phage AF. Virology 2012, 434, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Glonti, T.; Chanishvili, N.; Taylor, P.W. Bacteriophage-Derived Enzyme That Depolymerizes the Alginic Acid Capsule Associated with Cystic Fibrosis Isolates of Pseudomonas Aeruginosa. J. Appl. Microbiol. 2010, 108, 695–702. [Google Scholar] [CrossRef]
- Pires, D.P.; Cleto, S.; Sillankorva, S.; Azeredo, J.; Lu, T.K. Genetically Engineered Phages: A Review of Advances over the Last Decade. Microbiol. Mol. Biol. Rev. 2016, 80, 523–543. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Li, G.; Hu, Y.; Ou, W.; Wan, Y. Construction of a Synthetic Phage-Displayed Nanobody Library with CDR3 Regions Randomized by Trinucleotide Cassettes for Diagnostic Applications. J. Transl. Med. 2014, 12, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzakizwanayo, J.; Hanin, A.; Alves, D.R.; McCutcheon, B.; Dedi, C.; Salvage, J.; Knox, K.; Stewart, B.; Metcalfe, A.; Clark, J.; et al. Bacteriophage Can Prevent Encrustation and Blockage of Urinary Catheters by Proteus Mirabilis. Antimicrob. Agents Chemother. 2015, 60, 1530–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen, K.; Rørbo, N.; Rybtke, M.L.; Martinet, M.G.; Tolker-Nielsen, T.; Høiby, N.; Middelboe, M.; Ciofu, O.P. aeruginosa Flow-Cell Biofilms Are Enhanced by Repeated Phage Treatments but Can Be Eradicated by Phage-Ciprofloxacin Combination. Pathog. Dis. 2019, 77, ftz011. [Google Scholar] [CrossRef] [PubMed]
- Farooq, U.; Yang, Q.; Ullah, M.W.; Wang, S. Bacterial Biosensing: Recent Advances in Phage-Based Bioassays and Biosensors. Biosens. Bioelectron. 2018, 118, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Richter, L.; Ritchter-Janczuk, M.; Niedziółka-Jönsson, J.; Paczesny, J.; Hołyst, R. Recent Advances in Bacteriophage-Based Methods for Bacteria Detection. Drug Discov. Today 2018, 23, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Paczesny, J.; Richter, Ł.; Hołyst, R. Recent Progress in the Detection of Bacteria Using Bacteriophages: A Review. Viruses 2020, 12, 845. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Fuciños, P.; Azinheiro, S.; Carvalho, C.; Carvalho, J.; Prado, M. Specific Detection of Viable Salmonella Enteritidis by Phage Amplification Combined with QPCR (PAA-QPCR) in Spiked Chicken Meat Samples. Food Control 2019, 99, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Anany, H.; Brovko, L.; el Dougdoug, N.K.; Sohar, J.; Fenn, H.; Alasiri, N.; Jabrante, T.; Mangin, P.; Ali, M.M.; Kannan, B.; et al. Print to Detect: A Rapid and Ultrasensitive Phage-Based Dipstick Assay for Foodborne Pathogens. Anal. Bioanal. Chem. 2018, 410, 1217–1230. [Google Scholar] [CrossRef]
- Burnham, S.; Hu, J.; Anany, H.; Brovko, L.; Deiss, F.; Derda, R.; Griffiths, M.W. Towards Rapid On-Site Phage-Mediated Detection of Generic Escherichia Coli in Water Using Luminescent and Visual Readout. Anal. Bioanal. Chem. 2014, 406, 5685–5693. [Google Scholar] [CrossRef]
- Meile, S.; Kilcher, S.; Loessner, M.J.; Dunne, M. Reporter Phage-Based Detection of Bacterial Pathogens: Design Guidelines and Recent Developments. Viruses 2020, 12, 944. [Google Scholar] [CrossRef]
- Gibb, B.; Hyman, P.; Schneider, C.L. The Many Applications of Engineered Bacteriophages—An Overview. Pharmaceuticals 2021, 14, 634. [Google Scholar] [CrossRef]
- Poshtiban, S.; Javed, M.A.; Arutyunov, D.; Singh, A.; Banting, G.; Szymanski, C.M.; Evoy, S. Phage Receptor Binding Protein-Based Magnetic Enrichment Method as an Aid for Real Time PCR Detection of Foodborne Bacteria. Analyst 2013, 138, 5619–5626. [Google Scholar] [CrossRef]
- Weber-Dabrowska, B.; Jończyk-Matysiak, E.; Zaczek, M.; Łobocka, M.; Łusiak-Szelachowska, M.; Górski, A. Bacteriophage Procurement for Therapeutic Purposes. Front. Microbiol. 2016, 7, 1177. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic Engineering of Bacteriophages Against Infectious Diseases. Front. Microbiol. 2019, 10, 954. [Google Scholar] [CrossRef]
- Marinelli, L.J.; Hatfull, G.F.; Piuri, M. Recombineering. Bacteriophage 2012, 2, 5–14. [Google Scholar] [CrossRef]
- Marinelli, L.J.; Piuri, M.; Swigonová, Z.; Balachandran, A.; Oldfield, L.M.; van Kessel, J.C.; Hatfull, G.F. BRED: A Simple and Powerful Tool for Constructing Mutant and Recombinant Bacteriophage Genomes. PLoS ONE 2008, 3, e3957. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Makarova, K.S.; Zhang, F. Diversity, Classification and Evolution of CRISPR-Cas Systems. Curr. Opin. Microbiol. 2017, 37, 67–78. [Google Scholar] [CrossRef]
- Box, A.M.; McGuffie, M.J.; O’Hara, B.J.; Seed, K.D. Functional Analysis of Bacteriophage Immunity through a Type I-E CRISPR-Cas System in Vibrio Cholerae and Its Application in Bacteriophage Genome Engineering. J. Bacteriol. 2016, 198, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Duong, M.M.; Carmody, C.M.; Ma, Q.; Peters, J.E.; Nugen, S.R. Optimization of T4 Phage Engineering via CRISPR_Cas9. Sci. Rep. 2020, 10, 18229. [Google Scholar] [CrossRef]
- Kiro, R.; Shitrit, D.; Qimron, U. Efficient Engineering of a Bacteriophage Genome Using the Type I-E CRISPR-Cas System. RNA Biol. 2014, 11, 42–44. [Google Scholar] [CrossRef] [Green Version]
- Tao, P.; Wu, X.; Tang, W.C.; Zhu, J.; Rao, V. Engineering of Bacteriophage T4 Genome Using CRISPR-Cas9. ACS Synth. Biol. 2017, 6, 1952–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.O.; Hutchison, C.A.; Pfannkoch, C.; Venter, J.C. Generating a Synthetic Genome by Whole Genome Assembly_ ΦX174 Bacteriophage from Synthetic Oligonucleotides. Proc. Natl. Acad. Sci. USA 2003, 100, 15440–15445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.Y.; Kosuri, S.; Endy, D. Refactoring Bacteriophage T7. Mol. Syst. Biol. 2005, 1, 2005.0018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaschke, P.R.; Lieberman, E.K.; Rodriguez, J.; Sierra, A.; Endy, D. A Fully Decompressed Synthetic Bacteriophage ØX174 Genome Assembled and Archived in Yeast. Virology 2012, 434, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered Bacteriophages for Treatment of a Patient with a Disseminated Drug-Resistant Mycobacterium Abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef]
- Lu, T.K.; Collins, J.J. Dispersing Biofilms with Engineered Enzymatic Bacteriophage. Proc. Natl. Acad. Sci. USA 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [Green Version]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Rohwer, F. Global Phage Diversity. Cell 2003, 113, 141. [Google Scholar] [CrossRef] [Green Version]
- Suttle, C.A. Viruses in the Sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Glenn, L.M. Ethical Issues in Genetic Engineering and Transgenics, UC Santa Cruz. 2014. Available online: https://escholarship.org/uc/item/4p82f4x8 (accessed on 30 September 2022).
- Servick, K.U.S. Center Will Fight Infections with Viruses: Proving Ground for Phage Therapy Will Organize Full Clinical Trials of the Approach. Science (1979) 2018, 360, 1280–1281. [Google Scholar]
- Bakhshinejad, B.; Karimi, M.; Khalaj-Kondori, M. Phage Display: Development of Nanocarriers for Targeted Drug Delivery to the Brain. Neural Regen. Res. 2015, 10, 862–865. [Google Scholar] [CrossRef]
- Bakhshinejad, B.; Sadeghizadeh, M. Bacteriophages and Their Applications in the Diagnosis and Treatment of Hepatitis B Virus Infection. World J. Gastroenterol. 2014, 20, 11671–11683. [Google Scholar] [CrossRef]
- Budynek, P.; Dąbrowska, K.; Skaradziński, G.; Górski, A. Bacteriophages and Cancer. Arch. Microbiol. 2010, 192, 315–320. [Google Scholar] [CrossRef]
- Tawil, N.; Sacher, E.; Mandeville, R.; Meunier, M. Bacteriophages: Biosensing Tools for Multi-Drug Resistant Pathogens. Analyst 2014, 139, 1224–1236. [Google Scholar] [CrossRef]
- van Kan-Davelaar, H.E.; van Hest, J.C.; Cornelissen, J.J.; Koay, M.S. Using Viruses as Nanomedicines. Br. J. Pharmacol. 2014, 171, 4001–4009. [Google Scholar] [CrossRef]
- Ju, Z.; Sun, W. Drug Delivery Vectors Based on Filamentous Bacteriophages and Phage-Mimetic Nanoparticles. Drug Deliv. 2017, 24, 1898–1908. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Chen, Y.; Li, S.; Nguyen, H.G.; Niu, Z.; You, S.; Mello, C.M.; Lu, X.; Wang, Q. Chemical Modification of M13 Bacteriophage and Its Application in Cancer Cell Imaging. Bioconjug. Chem. 2010, 21, 1369–1377. [Google Scholar] [CrossRef]
- Kovacs, E.W.; Hooker, J.M.; Romanini, D.W.; Holder, P.G.; Berry, K.E.; Francis, M.B. Dual-Surface-Modified Bacteriophage MS2 as an Ideal Scaffold for a Viral Capsid-Based Drug Delivery System. Bioconjug. Chem. 2007, 18, 1140–1147. [Google Scholar] [CrossRef]
- Ceglarek, I.; Piotrowicz, A.; Lecion, D.; Miernikiewicz, P.; Owczarek, B.; Hodyra, K.; Harhala, M.; Górski, A.; Da̧browska, K. A Novel Approach for Separating Bacteriophages from Other Bacteriophages Using Affinity Chromatography and Phage Display. Sci. Rep. 2013, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Majewska, J.; Beta, W.; Lecion, D.; Hodyra-Stefaniak, K.; Kłopot, A.; Kazmiercźak, Z.; Miernikiewicz, P.; Piotrowicz, A.; Ciekot, J.; Owczarek, B.; et al. Oral Application of T4 Phage Induces Weak Antibody Production in the Gut and in the Blood. Viruses 2015, 7, 4783–4799. [Google Scholar] [CrossRef]
- Adhya, S.; Merril, C.; Biswas, B. Therapeutic and Prophylactic Applications of Bacteriophage Components in Modern Medicine. Cold Spring Harb. Perspect. Med. 2014, 4, a012518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, K.A.; Arbabi-Ghahroudi, M.; Scott, J.K. Beyond Phage Display: Non-Traditional Applications of the Filamentous Bacteriophage as a Vaccine Carrier, Therapeutic Biologic, and Bioconjugation Scaffold. Front. Microbiol. 2015, 6, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicastro, J.; Sheldon, K.; Slavcev, R. Bacteriophage Lambda Display Systems: Developments and Applications. Appl. Microbiol. Biotechnol. 2014, 98, 2853–2866. [Google Scholar] [CrossRef] [PubMed]
- Samoylova, T.I.; Cochran, A.M.; Samoylov, A.M.; Schemera, B.; Breiteneicher, A.H.; Ditchkoff, S.S.; Petrenko, V.A.; Cox, N.R. Phage Display Allows Identification of Zona Pellucida-Binding Peptides with Species-Specific Properties: Novel Approach for Development of Contraceptive Vaccines for Wildlife. J. Biotechnol. 2012, 162, 311–318. [Google Scholar] [CrossRef]
- Bao, Q.; Li, X.; Han, G.; Zhu, Y.; Mao, C.; Yang, M. Phage-Based Vaccines. Adv. Drug Deliv. Rev. 2019, 145, 40–56. [Google Scholar] [CrossRef]
- Lopes, R.S.; Queiroz, M.A.F.; Gomes, S.T.M.; Vallinoto, A.C.R.; Goulart, L.R.; Ishak, R. Phage Display: An Important Tool in the Discovery of Peptides with Anti-HIV Activity. Biotechnol. Adv. 2018, 36, 1847–1854. [Google Scholar] [CrossRef]
- Aghebati-Maleki, L.; Bakhshinejad, B.; Baradaran, B.; Motallebnezhad, M.; Aghebati-Maleki, A.; Nickho, H.; Yousefi, M.; Majidi, J. Phage Display as a Promising Approach for Vaccine Development. J. Biomed. Sci. 2016, 23, 66. [Google Scholar] [CrossRef] [Green Version]
- Noren, K.A.; Noren, C.J. Construction of High-Complexity Combinatorial Phage Display Peptide Libraries. Methods 2001, 23, 169–178. [Google Scholar] [CrossRef]
- Konthur, Z.; Wilde, J.; Lim, T.S. Semi-Automated Magnetic Bead-Based Antibody Selection from Phage Display Libraries. In Antibody Engineering; Springer: Berlin/Heidelberg, Germany, 2010; pp. 267–287. [Google Scholar]
- Jaroszewicz, W.; Morcinek-Orłowska, J.; Pierzynowska, K.; Gaffke, L.; Wȩgrzyn, G. Phage Display and Other Peptide Display Technologies. FEMS Microbiol. Rev. 2022, 46, fuab052. [Google Scholar] [CrossRef]
- Murai, R.; Nogi, T.; Tateoka, K.; Sato, A. Affinity Selection of Peptide Binders with Magnetic Beads via Organic Phase Separation (MOPS). Biol. Pharm. Bull. 2015, 38, 1822–1826. [Google Scholar] [CrossRef] [Green Version]
- González-Mora, A.; Hernández-Pérez, J.; Iqbal, H.M.N.; Rito-Palomares, M.; Benavides, J. Bacteriophage-Based Vaccines: A Potent Approach for Antigen Delivery. Vaccines 2020, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh-Mivehroud, M.; Alizadeh, A.A.; Morris, M.B.; Church, W.B.; Dastmalchi, S. Phage Display as a Technology Delivering on the Promise of Peptide Drug Discovery. Drug Discov. Today 2013, 18, 1144–1157. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.R.; Yoo, S.Y.; Lee, S.W.; Dean, D. Engineered Phage-Based Therapeutic Materials Inhibit Chlamydia Trachomatis Intracellular Infection. Biomaterials 2012, 33, 5166–5174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, M.N. Peptide-Displaying Phage Technology in Glycobiology. Glycobiology 2012, 22, 318–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.; Fowler, J.D.; Furst, E.M. Targeted Binding of the M13 Bacteriophage to Thiamethoxam Organic Crystals. Langmuir 2012, 28, 6013–6020. [Google Scholar] [CrossRef]
- Nobrega, F.L.; Rita Costa, A.; Kluskens, L.D.; Azeredo, J. Revisiting Phage Therapy: New Applications for Old Resources. Trends Microbiol. 2015, 23, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Bárdy, P.; Pantůček, R.; Benešík, M.; Doškař, J. Genetically Modified Bacteriophages in Applied Microbiology. J. Appl. Microbiol. 2016, 121, 618–633. [Google Scholar] [CrossRef]
- Kilcher, S.; Loessner, M.J. Engineering Bacteriophages as Versatile Biologics. Trends Microbiol. 2019, 27, 355–367. [Google Scholar] [CrossRef]
- Gonzalez-Menendez, E.; Fernandez, L.; Gutierrez, D.; Rodríguez, A.; Martínez, B.; GarcíaI, P. Comparative Analysis of Different Preservation Techniques for the Storage of Staphylococcus Phages Aimed for the Industrial Development of Phage-Based Antimicrobial Products. PLoS ONE 2018, 13, e0205728. [Google Scholar] [CrossRef]
- Burrowes, B.H.; Molineux, I.J.; Fralick, J.A. Directed in Vitro Evolution of Therapeutic Bacteriophages: The Appelmans Protocol. Viruses 2019, 11, 241. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elois, M.A.; Silva, R.d.; Pilati, G.V.T.; Rodríguez-Lázaro, D.; Fongaro, G. Bacteriophages as Biotechnological Tools. Viruses 2023, 15, 349. https://doi.org/10.3390/v15020349
Elois MA, Silva Rd, Pilati GVT, Rodríguez-Lázaro D, Fongaro G. Bacteriophages as Biotechnological Tools. Viruses. 2023; 15(2):349. https://doi.org/10.3390/v15020349
Chicago/Turabian StyleElois, Mariana Alves, Raphael da Silva, Giulia Von Tönnemann Pilati, David Rodríguez-Lázaro, and Gislaine Fongaro. 2023. "Bacteriophages as Biotechnological Tools" Viruses 15, no. 2: 349. https://doi.org/10.3390/v15020349