
Seroprevalence of Binding and Neutralizing Antibodies against 39 Human Adenovirus Types in Patients with Neuromuscular Disorders
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Human Samples
2.3. Cells
2.4. Viruses
2.5. Binding Antibody ELISA
2.6. IC ELISA for Detection of Neutralizing Antibodies
2.7. Statistical Analysis
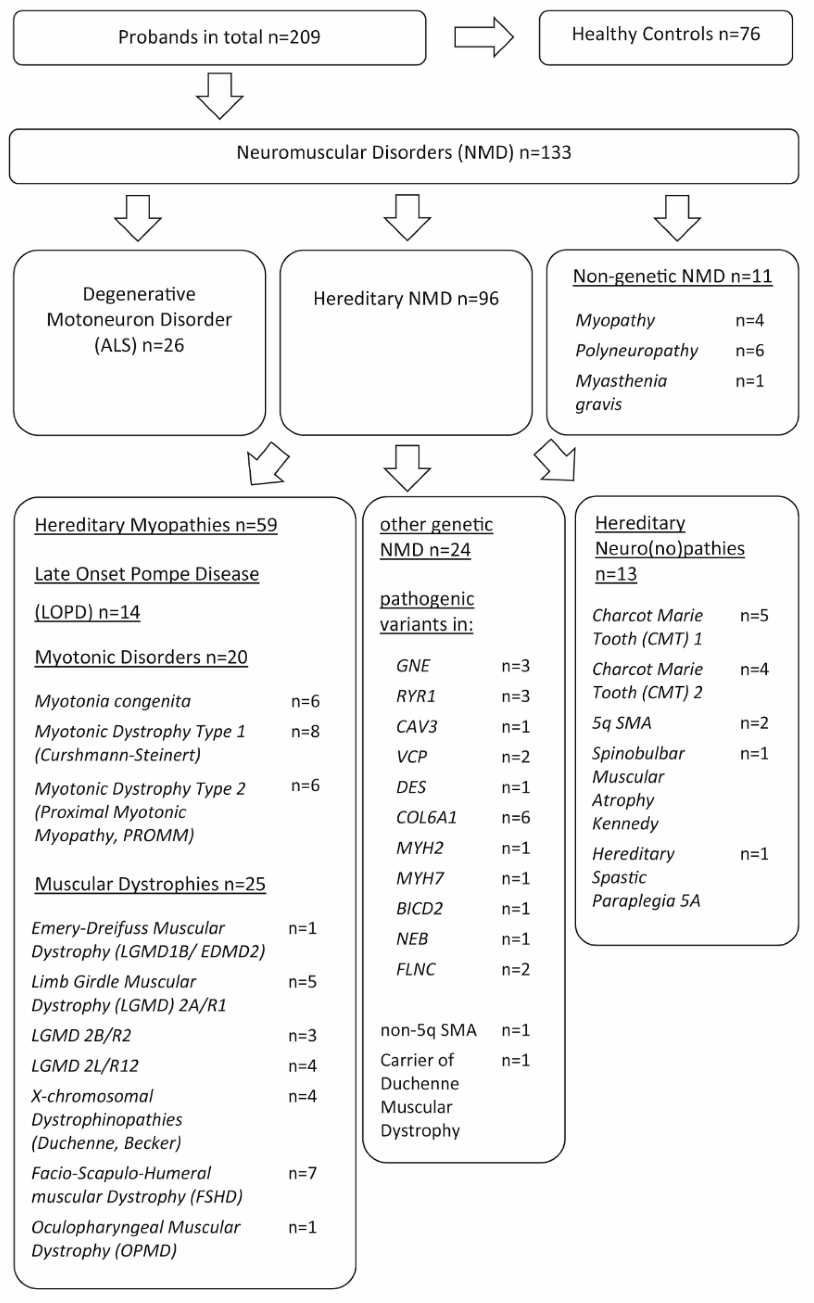
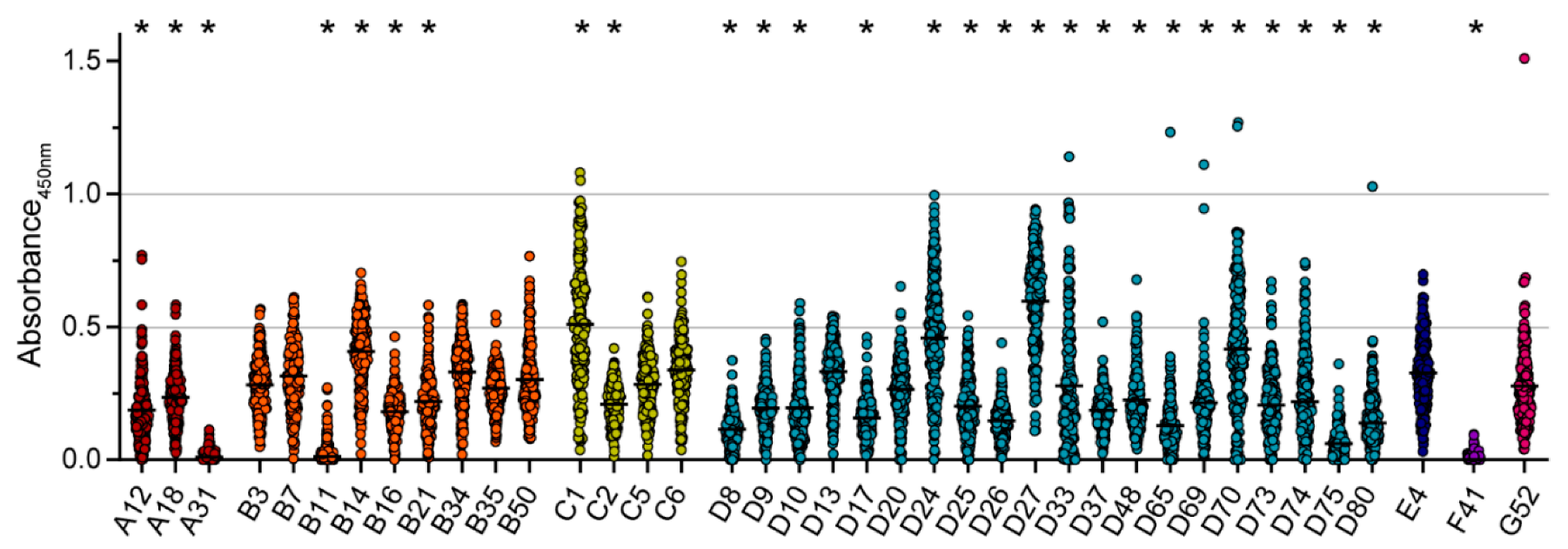
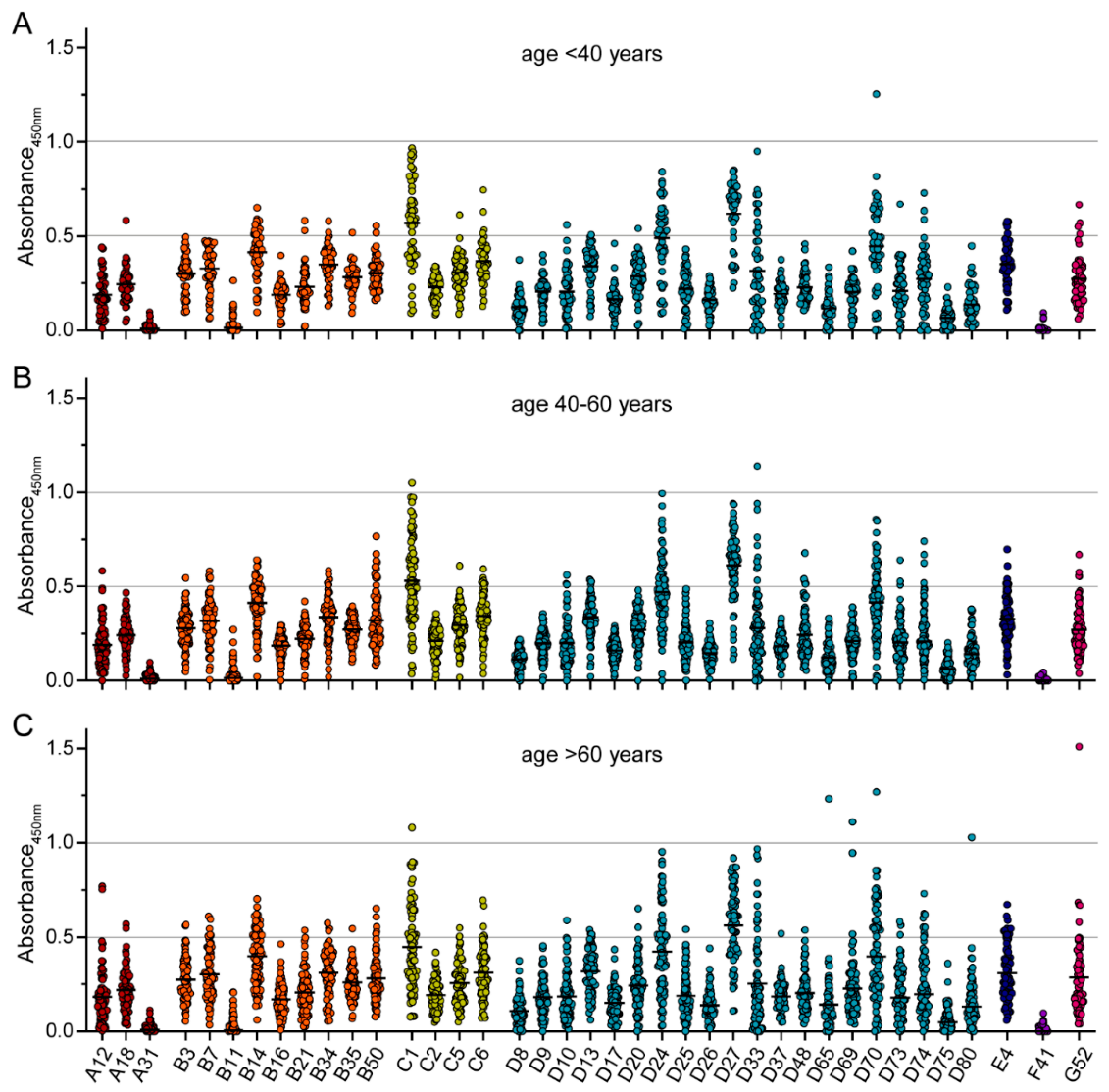
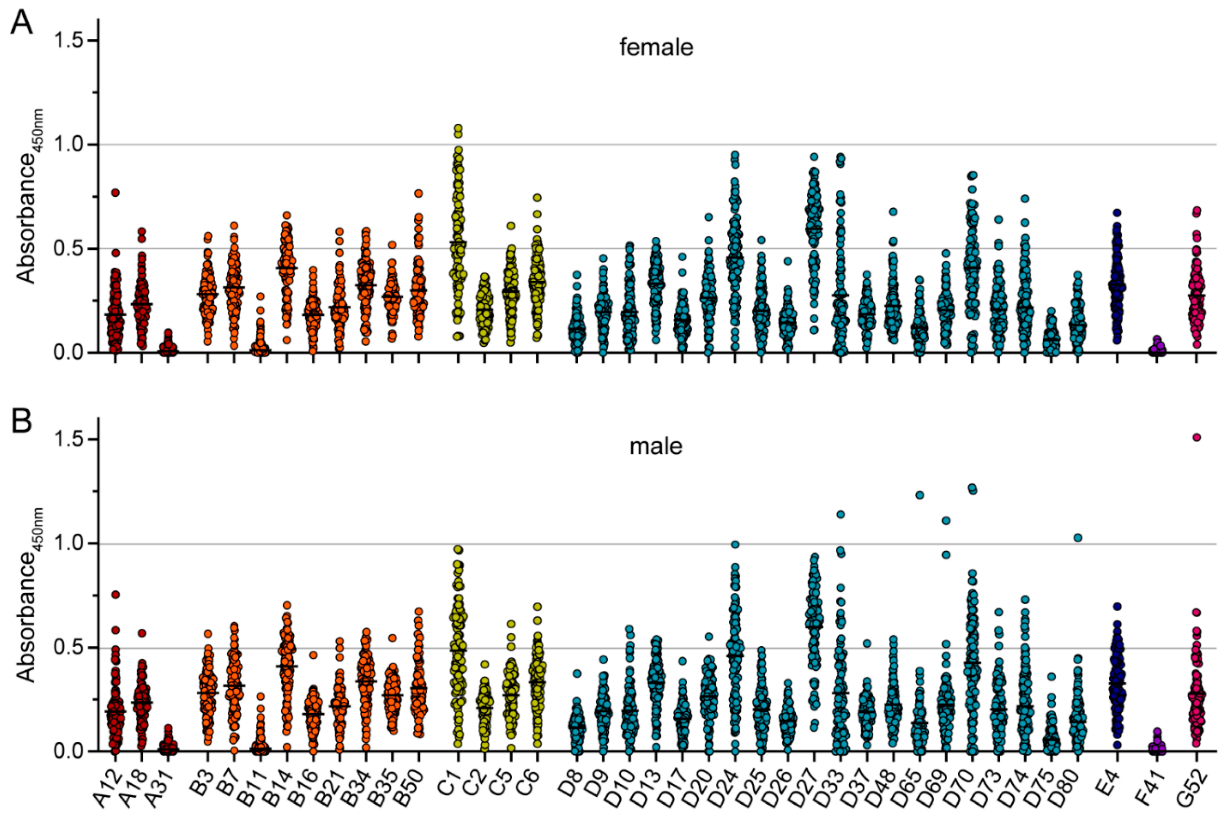
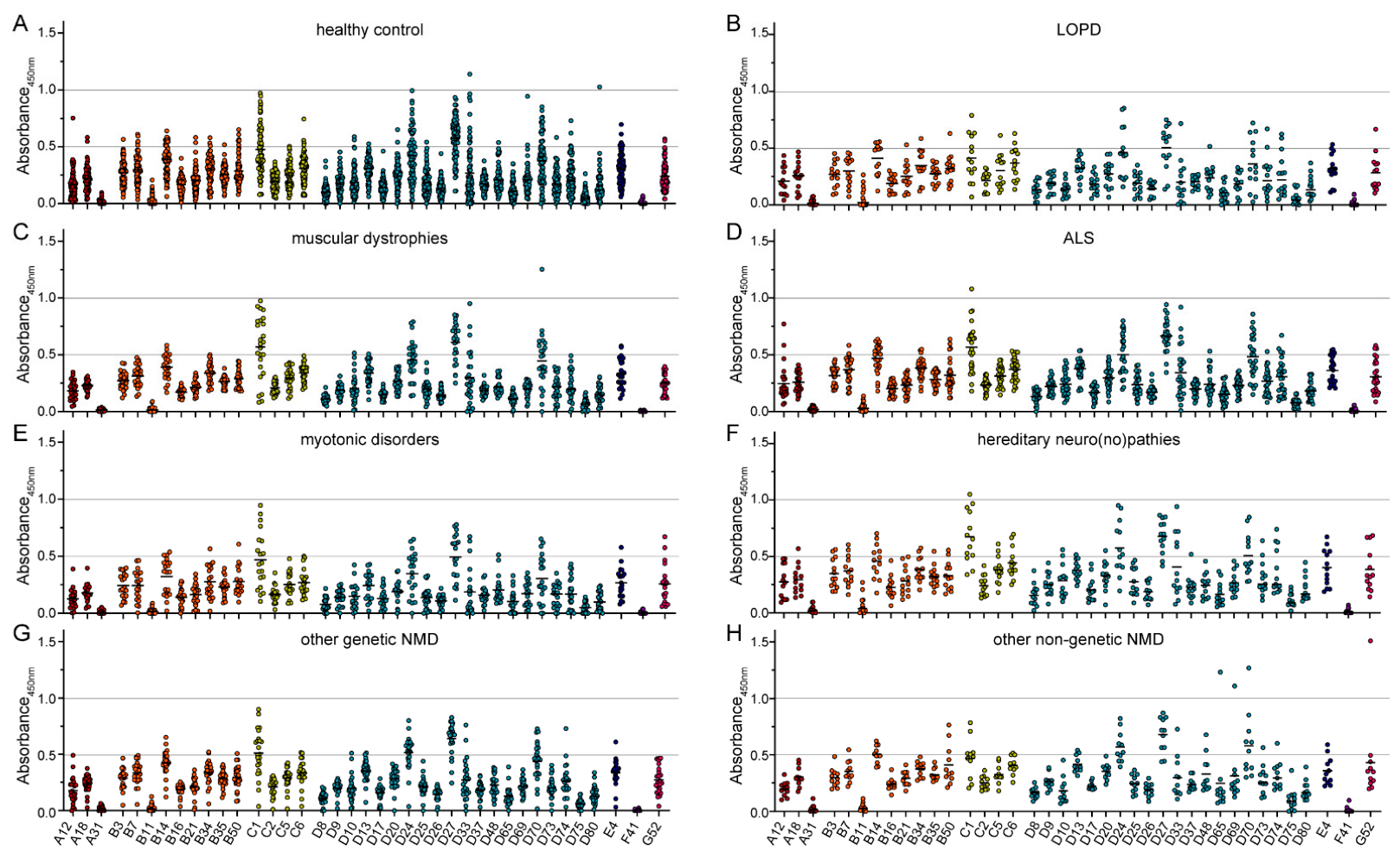
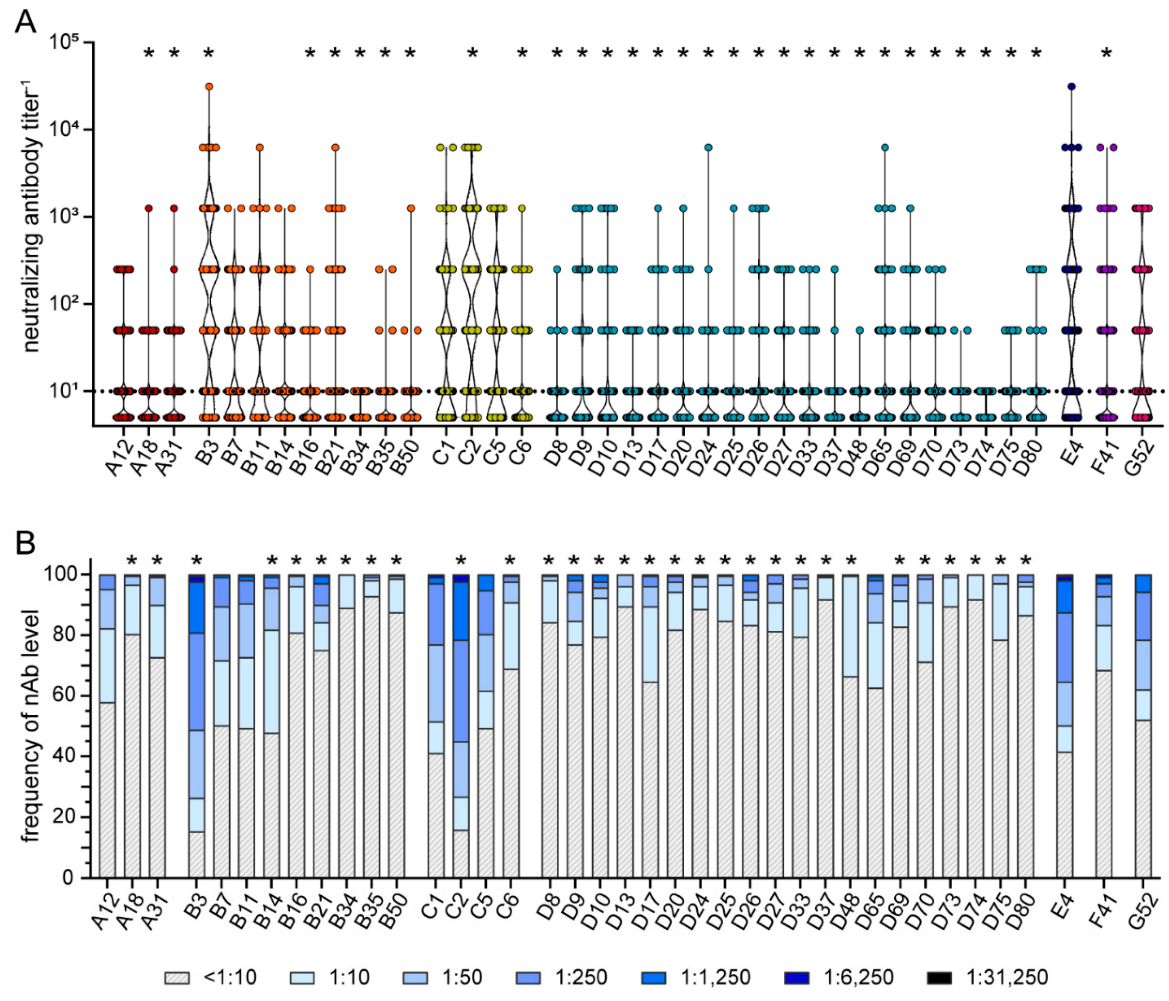
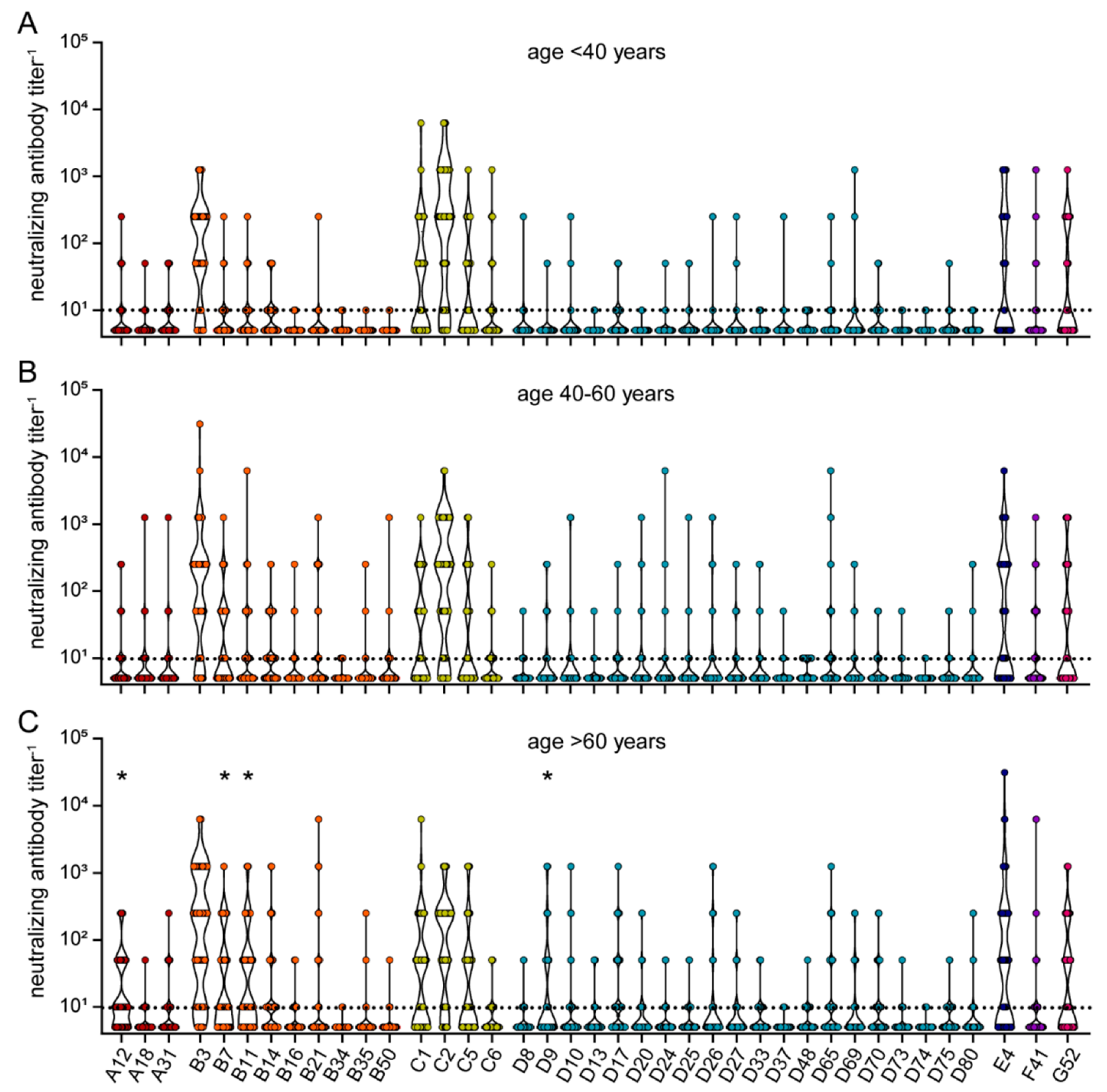
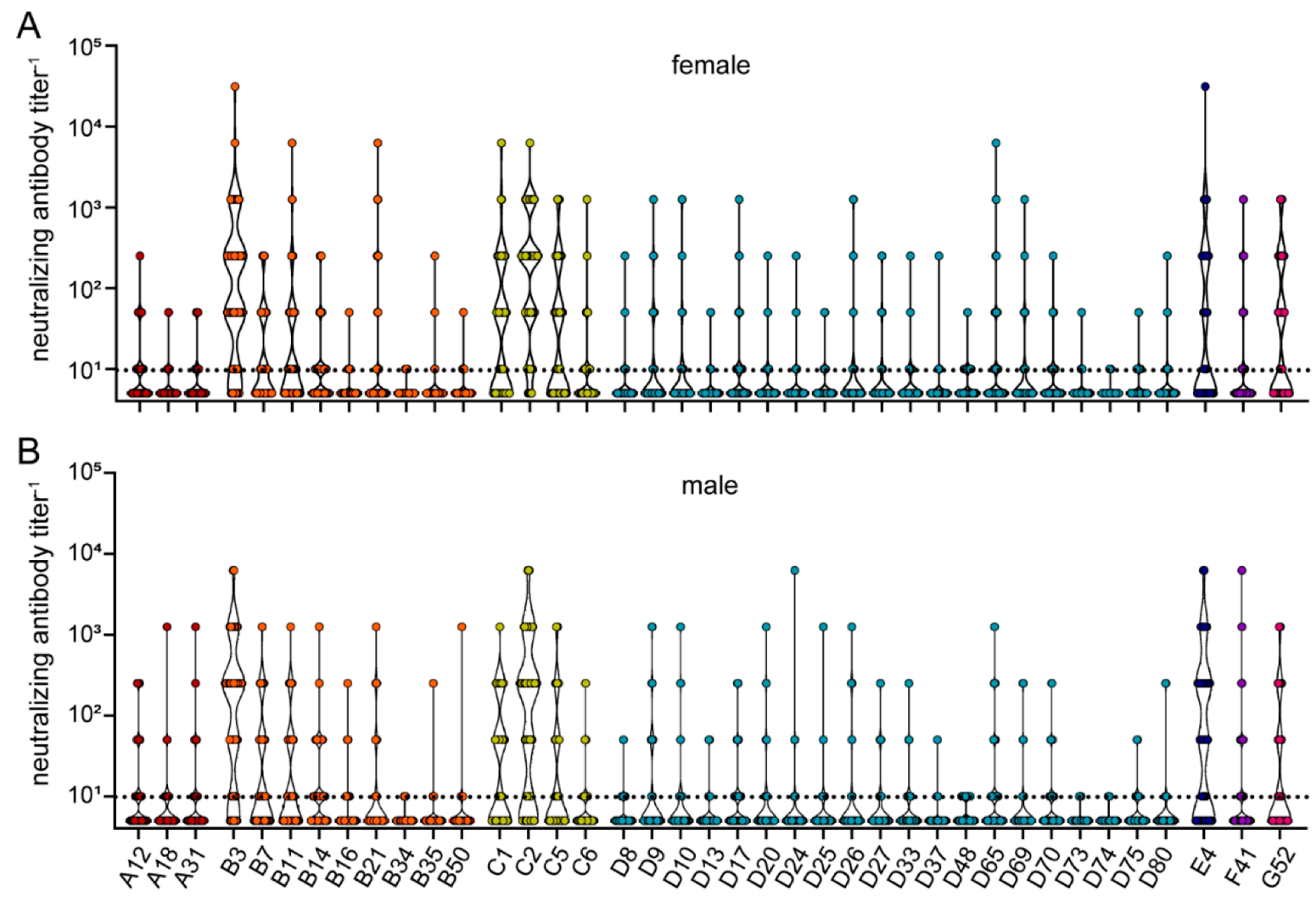
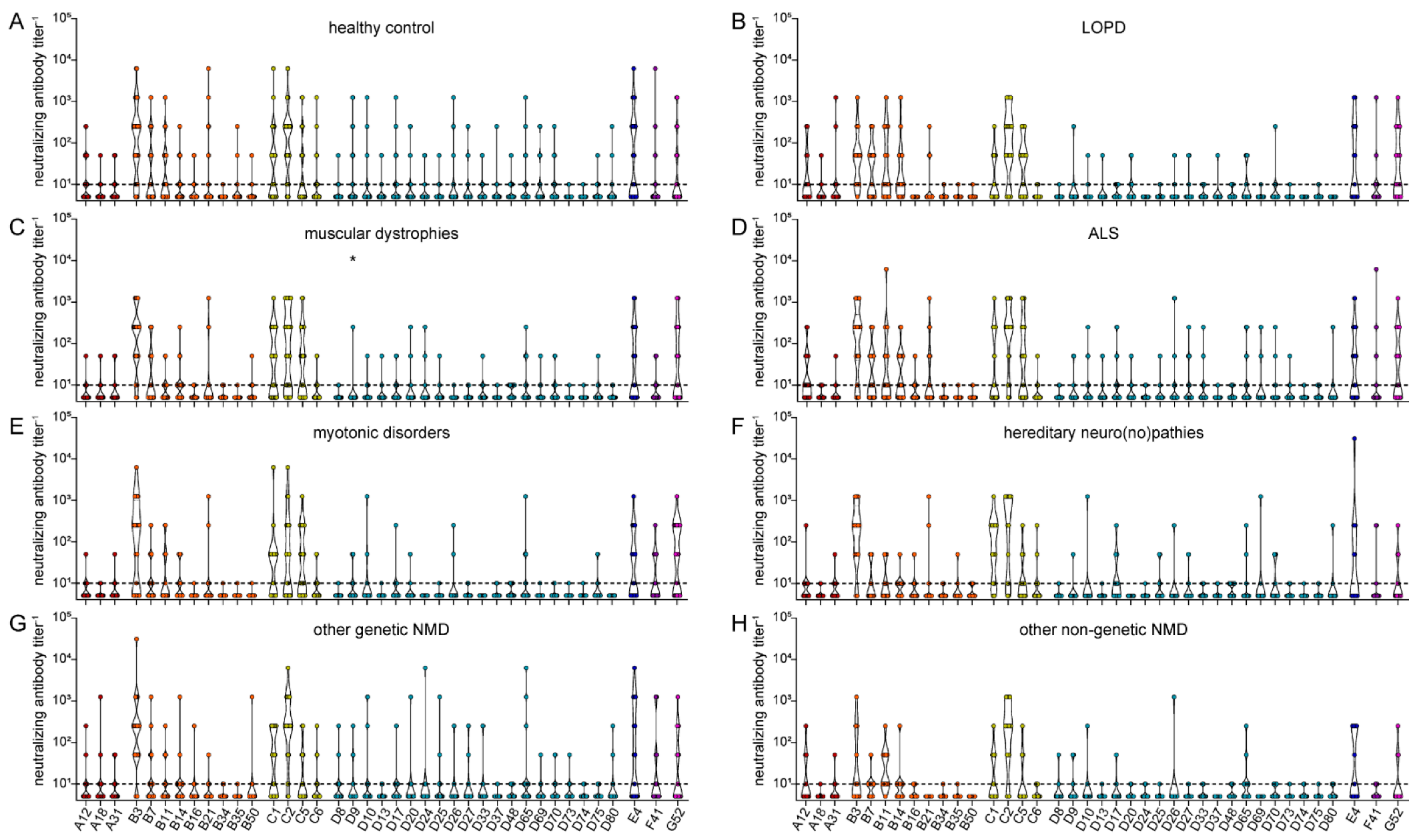
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seto, D.; Chodosh, J.; Brister, J.R.; Jones, M.S.; Members of the Adenovirus Research, C. Using the whole-genome sequence to characterize and name human adenoviruses. J. Virol. 2011, 85, 5701–5702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, C.M.; Singh, G.; Lee, J.Y.; Dehghan, S.; Rajaiya, J.; Liu, E.B.; Yousuf, M.A.; Betensky, R.A.; Jones, M.S.; Dyer, D.W.; et al. Molecular evolution of human adenoviruses. Sci. Rep. 2013, 3, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Mese, K.; Bunz, O.; Ehrhardt, A. State-of-the-art human adenovirus vectorology for therapeutic approaches. FEBS Lett. 2019, 593, 3609–3622. [Google Scholar] [CrossRef] [Green Version]
- Saito, I.; Oya, Y.; Yamamoto, K.; Yuasa, T.; Shimojo, H. Construction of nondefective adenovirus type 5 bearing a 2.8-kilobase hepatitis B virus DNA near the right end of its genome. J. Virol. 1985, 54, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Lewis, J.A.; Grodzicker, T. Overproduction of the protein product of a nonselected foreign gene carried by an adenovirus vector. Proc. Natl. Acad. Sci. USA 1985, 82, 3567–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haj-Ahmad, Y.; Graham, F.L. Development of a helper-independent human adenovirus vector and its use in the transfer of the herpes simplex virus thymidine kinase gene. J. Virol. 1986, 57, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Massie, B.; Gluzman, Y.; Hassell, J.A. Construction of a helper-free recombinant adenovirus that expresses polyomavirus large T antigen. Mol. Cell. Biol. 1986, 6, 2872–2883. [Google Scholar] [CrossRef]
- Saito, I.; Oya, Y.; Shimojo, H. Novel RNA family structure of hepatitis B virus expressed in human cells, using a helper-free adenovirus vector. J. Virol. 1986, 58, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Tomita, K.; Sakurai, F.; Tachibana, M.; Mizuguchi, H. Correlation between adenovirus-neutralizing antibody titer and adenovirus vector-mediated transduction efficiency following intratumoral injection. Anticancer. Res. 2012, 32, 1145–1152. [Google Scholar]
- Gray, G.E.; Allen, M.; Moodie, Z.; Churchyard, G.; Bekker, L.G.; Nchabeleng, M.; Mlisana, K.; Metch, B.; de Bruyn, G.; Latka, M.H.; et al. Safety and efficacy of the HVTN 503/Phambili study of a clade-B-based HIV-1 vaccine in South Africa: A double-blind, randomised, placebo-controlled test-of-concept phase 2b study. Lancet Infect. Dis. 2011, 11, 507–515. [Google Scholar] [CrossRef] [Green Version]
- McElrath, M.J.; De Rosa, S.C.; Moodie, Z.; Dubey, S.; Kierstead, L.; Janes, H.; Defawe, O.D.; Carter, D.K.; Hural, J.; Akondy, R.; et al. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: A case-cohort analysis. Lancet 2008, 372, 1894–1905. [Google Scholar] [CrossRef] [PubMed]
- Tatsis, N.; Ertl, H.C. Adenoviruses as vaccine vectors. Mol. Ther. 2004, 10, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Milligan, I.D.; Gibani, M.M.; Sewell, R.; Clutterbuck, E.A.; Campbell, D.; Plested, E.; Nuthall, E.; Voysey, M.; Silva-Reyes, L.; McElrath, M.J.; et al. Safety and Immunogenicity of Novel Adenovirus Type 26- and Modified Vaccinia Ankara-Vectored Ebola Vaccines: A Randomized Clinical Trial. JAMA 2016, 315, 1610–1623. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Wurie, A.H.; Hou, L.H.; Liang, Q.; Li, Y.H.; Russell, J.B.; Wu, S.P.; Li, J.X.; Hu, Y.M.; Guo, Q.; et al. Safety and immunogenicity of a recombinant adenovirus type-5 vector-based Ebola vaccine in healthy adults in Sierra Leone: A single-centre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 389, 621–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Dowling, J.J.; Gonorazky, H.D.; Cohn, R.D.; Campbell, C. Treating pediatric neuromuscular disorders: The future is now. Am. J. Med. Genet. A 2018, 176, 804–841. [Google Scholar] [CrossRef] [Green Version]
- Chikkannaiah, M.; Reyes, I. New diagnostic and therapeutic modalities in neuromuscular disorders in children. Curr. Probl. Pediatr. Adolesc. Health Care 2021, 51, 101033. [Google Scholar] [CrossRef]
- Mendell, J.R.; Al-Zaidy, S.; Shell, R.; Arnold, W.D.; Rodino-Klapac, L.R.; Prior, T.W.; Lowes, L.; Alfano, L.; Berry, K.; Church, K.; et al. Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N. Engl. J. Med. 2017, 377, 1713–1722. [Google Scholar] [CrossRef]
- Kirschner, J.; Butoianu, N.; Goemans, N.; Haberlova, J.; Kostera-Pruszczyk, A.; Mercuri, E.; van der Pol, W.L.; Quijano-Roy, S.; Sejersen, T.; Tizzano, E.F.; et al. European ad-hoc consensus statement on gene replacement therapy for spinal muscular atrophy. Eur. J. Paediatr. Neurol. 2020, 28, 38–43. [Google Scholar] [CrossRef]
- Flotats-Bastardas, M.; Hahn, A. New Therapeutics Options for Pediatric Neuromuscular Disorders. Front. Pediatr. 2020, 8, 583877. [Google Scholar] [CrossRef] [PubMed]
- Benchaouir, R.; Robin, V.; Goyenvalle, A. Gene and splicing therapies for neuromuscular diseases. Front. Biosci. 2015, 20, 1190–1233. [Google Scholar] [CrossRef] [Green Version]
- Elangkovan, N.; Dickson, G. Gene Therapy for Duchenne Muscular Dystrophy. J. Neuromuscul. Dis. 2021, 8, S303–S316. [Google Scholar] [CrossRef]
- Manini, A.; Abati, E.; Nuredini, A.; Corti, S.; Comi, G.P. Adeno-Associated Virus (AAV)-Mediated Gene Therapy for Duchenne Muscular Dystrophy: The Issue of Transgene Persistence. Front. Neurol. 2021, 12, 814174. [Google Scholar] [CrossRef]
- Mueller, C.; Berry, J.D.; McKenna-Yasek, D.M.; Gernoux, G.; Owegi, M.A.; Pothier, L.M.; Douthwright, C.L.; Gelevski, D.; Luppino, S.D.; Blackwood, M.; et al. Suppression with Adeno-Associated Virus and MicroRNA in Familial ALS. N. Engl. J. Med. 2020, 383, 151–158. [Google Scholar] [CrossRef]
- Mendell, J.R.; Rodino-Klapac, L.R.; Rosales-Quintero, X.; Kota, J.; Coley, B.D.; Galloway, G.; Craenen, J.M.; Lewis, S.; Malik, V.; Shilling, C.; et al. Limb-girdle muscular dystrophy type 2D gene therapy restores alpha-sarcoglycan and associated proteins. Ann. Neurol. 2009, 66, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Colella, P.; Sellier, P.; Gomez, M.J.; Biferi, M.G.; Tanniou, G.; Guerchet, N.; Cohen-Tannoudji, M.; Moya-Nilges, M.; van Wittenberghe, L.; Daniele, N.; et al. Gene therapy with secreted acid alpha-glucosidase rescues Pompe disease in a novel mouse model with early-onset spinal cord and respiratory defects. EBioMedicine 2020, 61, 103052. [Google Scholar] [CrossRef] [PubMed]
- Kagiava, A.; Karaiskos, C.; Richter, J.; Tryfonos, C.; Jennings, M.J.; Heslegrave, A.J.; Sargiannidou, I.; Stavrou, M.; Zetterberg, H.; Reilly, M.M.; et al. AAV9-mediated Schwann cell-targeted gene therapy rescues a model of demyelinating neuropathy. Gene. Ther. 2021, 28, 659–675. [Google Scholar] [CrossRef]
- Arimura, S.; Okada, T.; Tezuka, T.; Chiyo, T.; Kasahara, Y.; Yoshimura, T.; Motomura, M.; Yoshida, N.; Beeson, D.; Takeda, S.; et al. Neuromuscular disease. DOK7 gene therapy benefits mouse models of diseases characterized by defects in the neuromuscular junction. Science 2014, 345, 1505–1508. [Google Scholar] [CrossRef]
- Okubo, M.; Minami, N.; Goto, K.; Goto, Y.; Noguchi, S.; Mitsuhashi, S.; Nishino, I. Genetic diagnosis of Duchenne/Becker muscular dystrophy using next-generation sequencing: Validation analysis of DMD mutations. J. Hum. Genet. 2016, 61, 483–489. [Google Scholar] [CrossRef]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Shayakhmetov, D.M.; Li, Z.Y.; Gaggar, A.; Gharwan, H.; Ternovoi, V.; Sandig, V.; Lieber, A. Genome size and structure determine efficiency of postinternalization steps and gene transfer of capsid-modified adenovirus vectors in a cell-type-specific manner. J. Virol. 2004, 78, 10009–10022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, G.; Kennel, P.; Pettmann, B.; Vigne, E.; Akli, S.; Revah, F.; Schmalbruch, H.; Kahn, A. Gene therapy of murine motor neuron disease using adenoviral vectors for neurotrophic factors. Nat. Med. 1997, 3, 429–436. [Google Scholar] [CrossRef]
- Gravel, C.; Gotz, R.; Lorrain, A.; Sendtner, M. Adenoviral gene transfer of ciliary neurotrophic factor and brain-derived neurotrophic factor leads to long-term survival of axotomized motor neurons. Nat. Med. 1997, 3, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.; Nagano, I.; Gazi, M.S.; Murakami, T.; Shiote, M.; Shoji, M.; Kitagawa, H.; Setoguchi, Y.; Abe, K. Adenovirus-mediated gene transfer of glial cell line-derived neurotrophic factor prevents motor neuron loss of transgenic model mice for amyotrophic lateral sclerosis. Apoptosis 2002, 7, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Tsujino, S.; Kinoshita, N.; Tashiro, T.; Ikeda, K.; Ichihara, N.; Kikuchi, H.; Hagiwara, Y.; Mizutani, M.; Kikuchi, T.; Sakuragawa, N. Adenovirus-mediated transfer of human acid maltase gene reduces glycogen accumulation in skeletal muscle of Japanese quail with acid maltase deficiency. Hum. Gene. Ther. 1998, 9, 1609–1616. [Google Scholar] [CrossRef]
- Ding, E.Y.; Hodges, B.L.; Hu, H.; McVie-Wylie, A.J.; Serra, D.; Migone, F.K.; Pressley, D.; Chen, Y.T.; Amalfitano, A. Long-term efficacy after [E1-, polymerase-] adenovirus-mediated transfer of human acid-alpha-glucosidase gene into glycogen storage disease type II knockout mice. Hum. Gene. Ther. 2001, 12, 955–965. [Google Scholar] [CrossRef]
- Rastall, D.P.; Seregin, S.S.; Aldhamen, Y.A.; Kaiser, L.M.; Mullins, C.; Liou, A.; Ing, F.; Pereria-Hicks, C.; Godbehere-Roosa, S.; Palmer, D.; et al. Long-term, high-level hepatic secretion of acid alpha-glucosidase for Pompe disease achieved in non-human primates using helper-dependent adenovirus. Gene. Ther. 2016, 23, 743–752. [Google Scholar] [CrossRef]
- Xu, F.; Ding, E.; Migone, F.; Serra, D.; Schneider, A.; Chen, Y.T.; Amalfitano, A. Glycogen storage in multiple muscles of old GSD-II mice can be rapidly cleared after a single intravenous injection with a modified adenoviral vector expressing hGAA. J. Gene. Med. 2005, 7, 171–178. [Google Scholar] [CrossRef]
- Pauly, D.F.; Fraites, T.J.; Toma, C.; Bayes, H.S.; Huie, M.L.; Hirschhorn, R.; Plotz, P.H.; Raben, N.; Kessler, P.D.; Byrne, B.J. Intercellular transfer of the virally derived precursor form of acid alpha-glucosidase corrects the enzyme deficiency in inherited cardioskeletal myopathy Pompe disease. Hum. Gene. Ther. 2001, 12, 527–538. [Google Scholar] [CrossRef]
- Floyd, S.S., Jr.; Clemens, P.R.; Ontell, M.R.; Kochanek, S.; Day, C.S.; Yang, J.; Hauschka, S.D.; Balkir, L.; Morgan, J.; Moreland, M.S.; et al. Ex vivo gene transfer using adenovirus-mediated full-length dystrophin delivery to dystrophic muscles. Gene. Ther. 1998, 5, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lochmuller, H.; Luo, J.; Massie, B.; Nalbantoglu, J.; Karpati, G.; Petrof, B.J. Adenovirus-mediated dystrophin minigene transfer improves muscle strength in adult dystrophic (MDX) mice. Gene. Ther. 1998, 5, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Schiedner, G.; Gilchrist, S.C.; Kochanek, S.; Clemens, P.R. CTLA4Ig delivered by high-capacity adenoviral vector induces stable expression of dystrophin in mdx mouse muscle. Gene. Ther. 2004, 11, 1453–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Schiedner, G.; van Rooijen, N.; Liu, C.C.; Kochanek, S.; Clemens, P.R. Sustained muscle expression of dystrophin from a high-capacity adenoviral vector with systemic gene transfer of T cell costimulatory blockade. Mol. Ther. 2004, 10, 688–696. [Google Scholar] [CrossRef]
- Reay, D.P.; Bilbao, R.; Koppanati, B.M.; Cai, L.; O’Day, T.L.; Jiang, Z.; Zheng, H.; Watchko, J.F.; Clemens, P.R. Full-length dystrophin gene transfer to the mdx mouse in utero. Gene. Ther. 2008, 15, 531–536. [Google Scholar] [CrossRef]
- Cho, W.K.; Ebihara, S.; Nalbantoglu, J.; Gilbert, R.; Massie, B.; Holland, P.; Karpati, G.; Petrof, B.J. Modulation of Starling forces and muscle fiber maturity permits adenovirus-mediated gene transfer to adult dystrophic (mdx) mice by the intravascular route. Hum. Gene. Ther. 2000, 11, 701–714. [Google Scholar] [CrossRef]
- Gilbert, R.; Nalbantoglu, J.; Petrof, B.J.; Ebihara, S.; Guibinga, G.H.; Tinsley, J.M.; Kamen, A.; Massie, B.; Davies, K.E.; Karpati, G. Adenovirus-mediated utrophin gene transfer mitigates the dystrophic phenotype of mdx mouse muscles. Hum. Gene. Ther. 1999, 10, 1299–1310. [Google Scholar] [CrossRef]
- Hartigan-O’Connor, D.; Kirk, C.J.; Crawford, R.; Mule, J.J.; Chamberlain, J.S. Immune evasion by muscle-specific gene expression in dystrophic muscle. Mol. Ther. 2001, 4, 525–533. [Google Scholar] [CrossRef]
- Matecki, S.; Dudley, R.W.; Divangahi, M.; Gilbert, R.; Nalbantoglu, J.; Karpati, G.; Petrof, B.J. Therapeutic gene transfer to dystrophic diaphragm by an adenoviral vector deleted of all viral genes. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2004, 287, L569–L576. [Google Scholar] [CrossRef]
- Mennechet, F.J.D.; Paris, O.; Ouoba, A.R.; Salazar Arenas, S.; Sirima, S.B.; Takoudjou Dzomo, G.R.; Diarra, A.; Traore, I.T.; Kania, D.; Eichholz, K.; et al. A review of 65 years of human adenovirus seroprevalence. Expert. Rev. Vaccines 2019, 18, 597–613. [Google Scholar] [CrossRef]
- Lynch, J.P.; Kajon, A.E. Adenovirus: Epidemiology, Global Spread of Novel Serotypes, and Advances in Treatment and Prevention. Semin. Respir. Crit. Care Med. 2016, 37, 586–602. [Google Scholar] [CrossRef]
- D’Ambrosio, E.; Del Grosso, N.; Chicca, A.; Midulla, M. Neutralizing antibodies against 33 human adenoviruses in normal children in Rome. J. Hyg. 1982, 89, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogels, R.; Zuijdgeest, D.; van Rijnsoever, R.; Hartkoorn, E.; Damen, I.; de Bethune, M.P.; Kostense, S.; Penders, G.; Helmus, N.; Koudstaal, W.; et al. Replication-deficient human adenovirus type 35 vectors for gene transfer and vaccination: Efficient human cell infection and bypass of preexisting adenovirus immunity. J. Virol. 2003, 77, 8263–8271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kerkmann, L.; Hetzel, M.; Windmann, S.; Trilling, M.; Zhang, W.; Ehrhardt, A.; Bayer, W. Analysis of the Prevalence of Binding and Neutralizing Antibodies against 39 Human Adenovirus Types in Student Cohorts Reveals Low-Prevalence Types and a Decline in Binding Antibody Levels during the SARS-CoV-2 Pandemic. J. Virol. 2022, 96, e01133-22. [Google Scholar] [CrossRef]
- Mroczek, M.; Sanchez, M.G. Genetic modifiers and phenotypic variability in neuromuscular disorders. J. Appl. Genet. 2020, 61, 547–558. [Google Scholar] [CrossRef]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef] [PubMed]
- James, K.T.; Cooney, B.; Agopsowicz, K.; Trevors, M.A.; Mohamed, A.; Stoltz, D.; Hitt, M.; Shmulevitz, M. Novel High-throughput Approach for Purification of Infectious Virions. Sci. Rep. 2016, 6, 36826. [Google Scholar] [CrossRef] [Green Version]
- Abbink, P.; Lemckert, A.A.; Ewald, B.A.; Lynch, D.M.; Denholtz, M.; Smits, S.; Holterman, L.; Damen, I.; Vogels, R.; Thorner, A.R.; et al. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 2007, 81, 4654–4663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrycak, C.P.; Windmann, S.; Bayer, W. Comparative Evaluation of the Vaccine Efficacies of Three Adenovirus-Based Vector Types in the Friend Retrovirus Infection Model. J. Virol. 2019, 93, e01155-19. [Google Scholar] [CrossRef]
- Huebner, R.J.; Rowe, W.P.; Ward, T.G.; Parrott, R.H.; Bell, J.A. Adenoidal-pharyngeal-conjunctival agents: A newly recognized group of common viruses of the respiratory system. N. Engl. J. Med. 1954, 251, 1077–1086. [Google Scholar] [CrossRef]
- Tai, F.H.; Grayston, J.T. Adenovirus neutralizing antibodies in persons on Taiwan. Proc. Soc. Exp. Biol. Med. 1962, 109, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Lemckert, A.A.C.; Grimbergen, J.; Smits, S.; Hartkoorn, E.; Holterman, L.; Berkhout, B.; Barouch, D.H.; Vogels, R.; Quax, P.; Goudsmit, J.; et al. Generation of a novel replication-incompetent adenoviral vector derived from human adenovirus type 49: Manufacture on PER.C6 cells, tropism and immunogenicity. J. Gen. Virol. 2006, 87, 2891–2899. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiang, Z.Q.; Li, Y.; Kurupati, R.K.; Jia, B.; Bian, A.; Zhou, D.M.; Hutnick, N.; Yuan, S.; Gray, C.; et al. Adenovirus-based vaccines: Comparison of vectors from three species of adenoviridae. J. Virol. 2010, 84, 10522–10532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast, T.C.; Kierstead, L.; Gupta, S.B.; Nikas, A.A.; Kallas, E.G.; Novitsky, V.; Mbewe, B.; Pitisuttithum, P.; Schechter, M.; Vardas, E.; et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: Correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010, 28, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Kik, S.V.; Weverling, G.J.; Dilan, R.; King, S.L.; Maxfield, L.F.; Clark, S.; Ng’ang’a, D.; Brandariz, K.L.; Abbink, P.; et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 2011, 29, 5203–5209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Zhou, Y.; Wu, H.; Wang, Z.; Zhan, Y.; Feng, X.; Geng, R.; Wu, Y.; Kong, W.; Yu, X. Seroprevalence of neutralizing antibodies to human adenovirus type 5 in healthy adults in China. J. Med. Virol. 2012, 84, 1408–1414. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): A single-blind, randomised, controlled, phase 2/3 trial. Lancet 2021, 396, 1979–1993. [Google Scholar] [CrossRef]
- Deenen, J.C.; Horlings, C.G.; Verschuuren, J.J.; Verbeek, A.L.; van Engelen, B.G. The Epidemiology of Neuromuscular Disorders: A Comprehensive Overview of the Literature. J. Neuromuscul. Dis. 2015, 2, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.L.; Waddington, S.N.; Buckley, S.M.; Custers, J.; Havenga, M.J.; van Rooijen, N.; Goudsmit, J.; McVey, J.H.; Nicklin, S.A.; Baker, A.H. Effect of neutralizing sera on factor x-mediated adenovirus serotype 5 gene transfer. J. Virol. 2009, 83, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Sterman, D.H.; Haas, A.; Moon, E.; Recio, A.; Schwed, D.; Vachani, A.; Katz, S.I.; Gillespie, C.T.; Cheng, G.; Sun, J.; et al. A trial of intrapleural adenoviral-mediated Interferon-alpha2b gene transfer for malignant pleural mesothelioma. Am. J. Respir. Crit. Care Med. 2011, 184, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Kumon, H.; Sasaki, K.; Ariyoshi, Y.; Sadahira, T.; Ebara, S.; Hiraki, T.; Kanazawa, S.; Yanai, H.; Watanabe, M.; Nasu, Y. Ad-REIC Gene Therapy: Promising Results in a Patient with Metastatic CRPC Following Chemotherapy. Clin. Med. Insights. Oncol. 2015, 9, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Povsic, T.J.; Henry, T.D.; Ohman, E.M.; Pepine, C.J.; Crystal, R.G.; Rosengart, T.K.; Reinhardt, R.R.; Dittrich, H.C.; Traverse, J.H.; Answini, G.A.; et al. Epicardial delivery of XC001 gene therapy for refractory angina coronary treatment (The EXACT Trial): Rationale, design, and clinical considerations. Am. Heart J. 2021, 241, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonero, R.; Bazan-Peregrino, M.; Gil-Martin, M.; Alvarez, R.; Macarulla, T.; Riesco-Martinez, M.C.; Verdaguer, H.; Guillen-Ponce, C.; Farrera-Sal, M.; Moreno, R.; et al. Phase I, multicenter, open-label study of intravenous VCN-01 oncolytic adenovirus with or without nab-paclitaxel plus gemcitabine in patients with advanced solid tumors. J. Immunother. Cancer 2022, 10, e003255. [Google Scholar] [CrossRef]
- Hutnick, N.A.; Carnathan, D.; Demers, K.; Makedonas, G.; Ertl, H.C.; Betts, M.R. Adenovirus-specific human T cells are pervasive, polyfunctional, and cross-reactive. Vaccine 2010, 28, 1932–1941. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, L. Factors Which Contribute to the Immunogenicity of Non-replicating Adenoviral Vectored Vaccines. Front Immunol. 2020, 11, 909. [Google Scholar] [CrossRef]
- Raper, S.E.; Chirmule, N.; Lee, F.S.; Wivel, N.A.; Bagg, A.; Gao, G.P.; Wilson, J.M.; Batshaw, M.L. Fatal systemic inflammatory response syndrome in a ornithine transcarbamylase deficient patient following adenoviral gene transfer. Mol. Genet. Metab. 2003, 80, 148–158. [Google Scholar] [CrossRef]
- Stasiak, A.C.; Stehle, T. Human adenovirus binding to host cell receptors: A structural view. Med. Microbiol. Immunol. 2020, 209, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Sinnreich, M.; Shaw, C.A.; Pari, G.; Nalbantoglu, J.; Holland, P.C.; Karpati, G. Localization of coxsackie virus and adenovirus receptor (CAR) in normal and regenerating human muscle. Neuromuscul. Disord. 2005, 15, 541–548. [Google Scholar] [CrossRef]
- Nalbantoglu, J.; Pari, G.; Karpati, G.; Holland, P.C. Expression of the primary coxsackie and adenovirus receptor is downregulated during skeletal muscle maturation and limits the efficacy of adenovirus-mediated gene delivery to muscle cells. Hum. Gene. Ther. 1999, 10, 1009–1019. [Google Scholar] [CrossRef]
- Larochelle, N.; Deol, J.R.; Srivastava, V.; Allen, C.; Mizuguchi, H.; Karpati, G.; Holland, P.C.; Nalbantoglu, J. Downregulation of CD46 during muscle differentiation: Implications for gene transfer to human skeletal muscle using group B adenoviruses. Hum. Gene. Ther. 2008, 19, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Patzke, C.; Max, K.E.; Behlke, J.; Schreiber, J.; Schmidt, H.; Dorner, A.A.; Kroger, S.; Henning, M.; Otto, A.; Heinemann, U.; et al. The coxsackievirus-adenovirus receptor reveals complex homophilic and heterophilic interactions on neural cells. J. Neurosci. 2010, 30, 2897–2910. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy | NMD Patients | Total | p Value * | |
|---|---|---|---|---|
| Serum donors | 76 | 133 | 209 | |
| Age (mean) (SD) | 60.12 (20.68) | 52.15 (15.76) | 55.04 (18.12) | 0.00449 |
| age < 40 years (n) | 15 | 34 | 49 | |
| age 40–60 years (n) | 24 | 60 | 84 | |
| age > 60 years (n) | 37 | 39 | 76 | |
| Weight (kg) (SD) | 76.71 (15.59) | 77.38 (19.46) | 77.14 (18.16) | 0.78906 |
| Height (cm) (SD) | 170.8 (12) | 172.2 (10.4) | 171.8 (11.1) | 0.38357 |
| BMI (kg/m²) (SD) | 26.08 (4.01) | 25.98 (5.58) | 26.02 (5.07) | 0.85816 |
| d.a. on weight and height (%) | 73 (96) | 129 (97) | 202 (96.7) | |
| Sex | 0.07991 | |||
| male (%) | 31 (40.8) | 71 (53.4) | 102 (48.8) | |
| female (%) | 45 (59.2) | 62 (46.6) | 107 (51.2) | |
| Neuromuscular Disorder | ||||
| late-onset Pompe disease (LOPD) (%) | 14 (10.5) | |||
| Muscular Dystrophies (%) | 25 (18.8) | |||
| Degenerative motoneuron disorder (ALS) (%) | 26 (19.6) | |||
| Myotonic Disorders (%) Hereditary Neuro(no)pathies (%) other genetic NMD (%) other non-genetic NMD (%) | 20 (15.0) 13 (9.8) 24 (18.1) 11 (8.3) | |||
| other diseases | ||||
| Hypertension (%) | 33 (43.4) | 37 (27.8) | 70 (33.5) | 0.02580 |
| Diabetes mellitus (%) | 4 (5.3) | 11 (8.27) | 15 (7.18) | 0.39409 |
| conditions after spinal disc herniation, spinal stenosis (%) | 4 (5.3) | 8 (6) | 12 (5.74) | 0.82038 |
| conditions after infarct (all localisations), TIA (%) | 35 (46.1) | 5 (3.76) | 40 (19.14) | <0.00001 |
| Hypothyroidism (%) | 4 (5.3) | 17 (12.8) | 21 (10.05) | 0.05435 |
| Depression (%) | 3 (3.95) | 6 (4.51) | 9 (4.31) | 0.84523 |
| Smoking (%) | 12 (19.05) | 30 (24.79) | 42 (22.83) | 0.22559 |
| d.a. on smoking (%) | 63 (82.89) | 121 (91) | 184 (88.04) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klann, P.J.; Wang, X.; Elfert, A.; Zhang, W.; Köhler, C.; Güttsches, A.-K.; Jacobsen, F.; Weyen, U.; Roos, A.; Ehrke-Schulz, E.; et al. Seroprevalence of Binding and Neutralizing Antibodies against 39 Human Adenovirus Types in Patients with Neuromuscular Disorders. Viruses 2023, 15, 79. https://doi.org/10.3390/v15010079
Klann PJ, Wang X, Elfert A, Zhang W, Köhler C, Güttsches A-K, Jacobsen F, Weyen U, Roos A, Ehrke-Schulz E, et al. Seroprevalence of Binding and Neutralizing Antibodies against 39 Human Adenovirus Types in Patients with Neuromuscular Disorders. Viruses. 2023; 15(1):79. https://doi.org/10.3390/v15010079
Chicago/Turabian StyleKlann, Patrick Julian, Xiaoyan Wang, Anna Elfert, Wenli Zhang, Cornelia Köhler, Anne-Katrin Güttsches, Frank Jacobsen, Ute Weyen, Andreas Roos, Eric Ehrke-Schulz, and et al. 2023. "Seroprevalence of Binding and Neutralizing Antibodies against 39 Human Adenovirus Types in Patients with Neuromuscular Disorders" Viruses 15, no. 1: 79. https://doi.org/10.3390/v15010079