
Rapid and Non-Invasive Detection of Aedes aegypti Co-Infected with Zika and Dengue Viruses Using Near Infrared Spectroscopy

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Experimental Infections
2.3. Scanning of Mosquitoes Using NIR
2.4. Confirmation of ZIKV and DENV Infections by RT-qPCR
2.5. Data Analysis
2.6. Ethical Considerations
3. Results
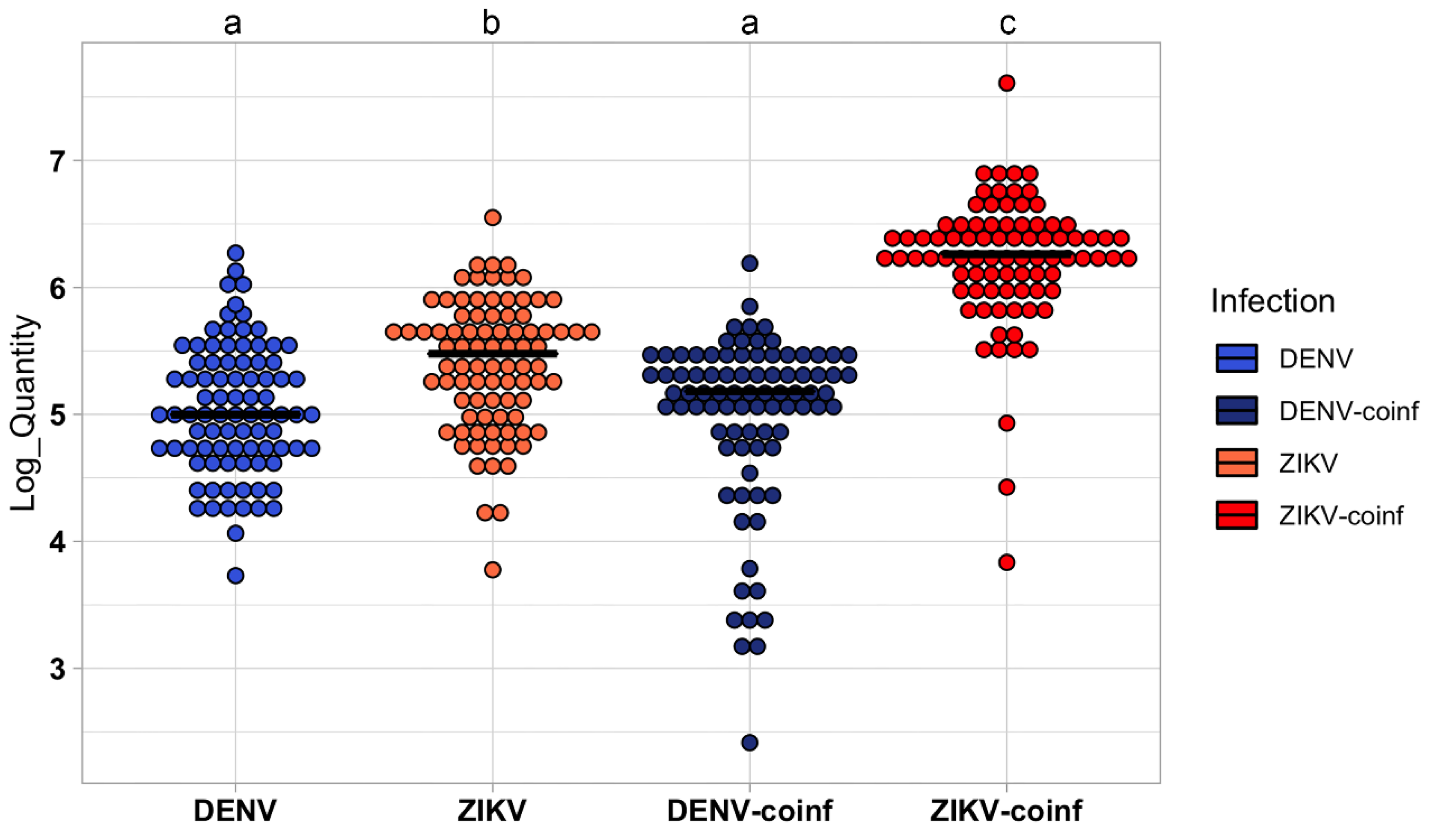
3.1. Infection Results
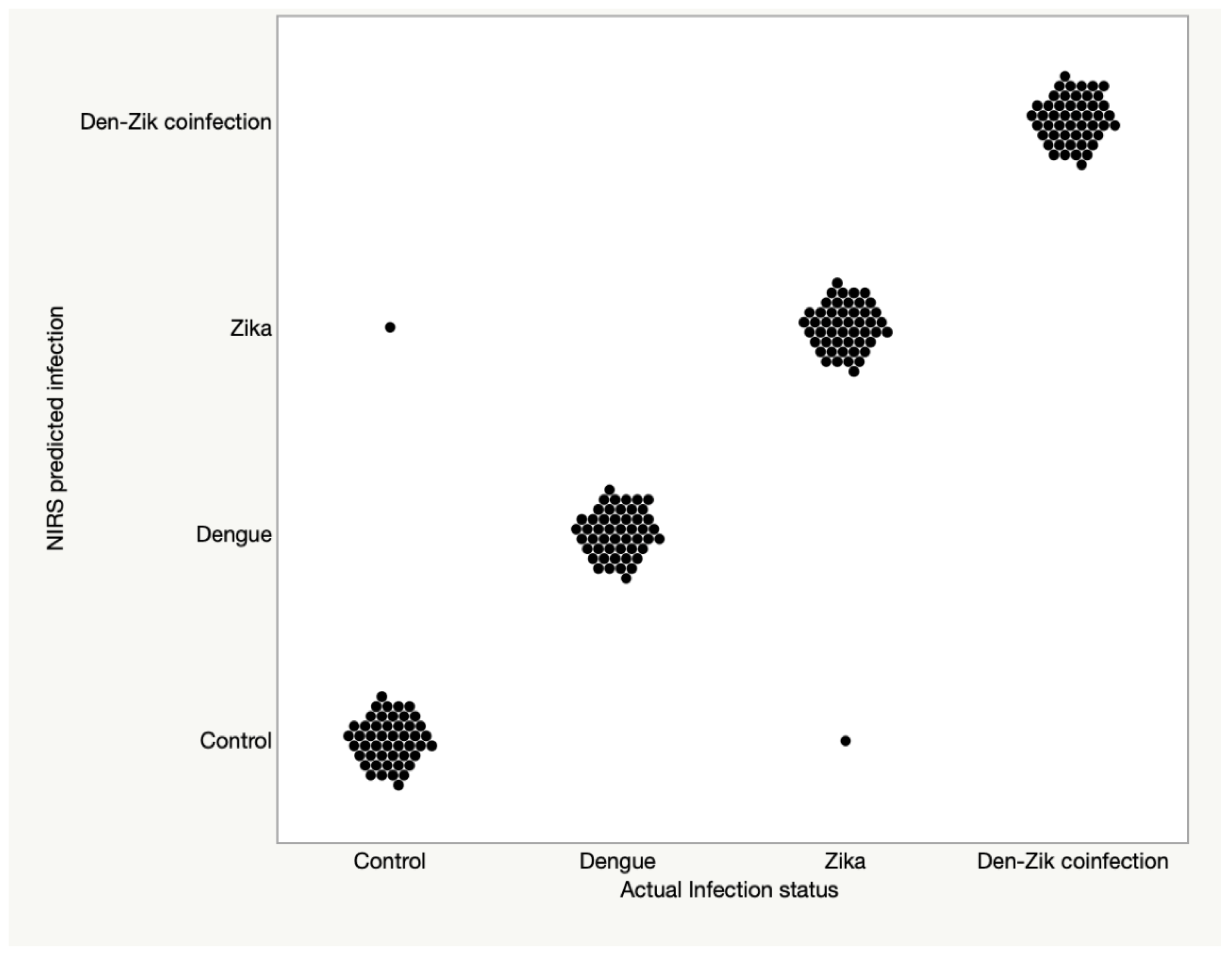
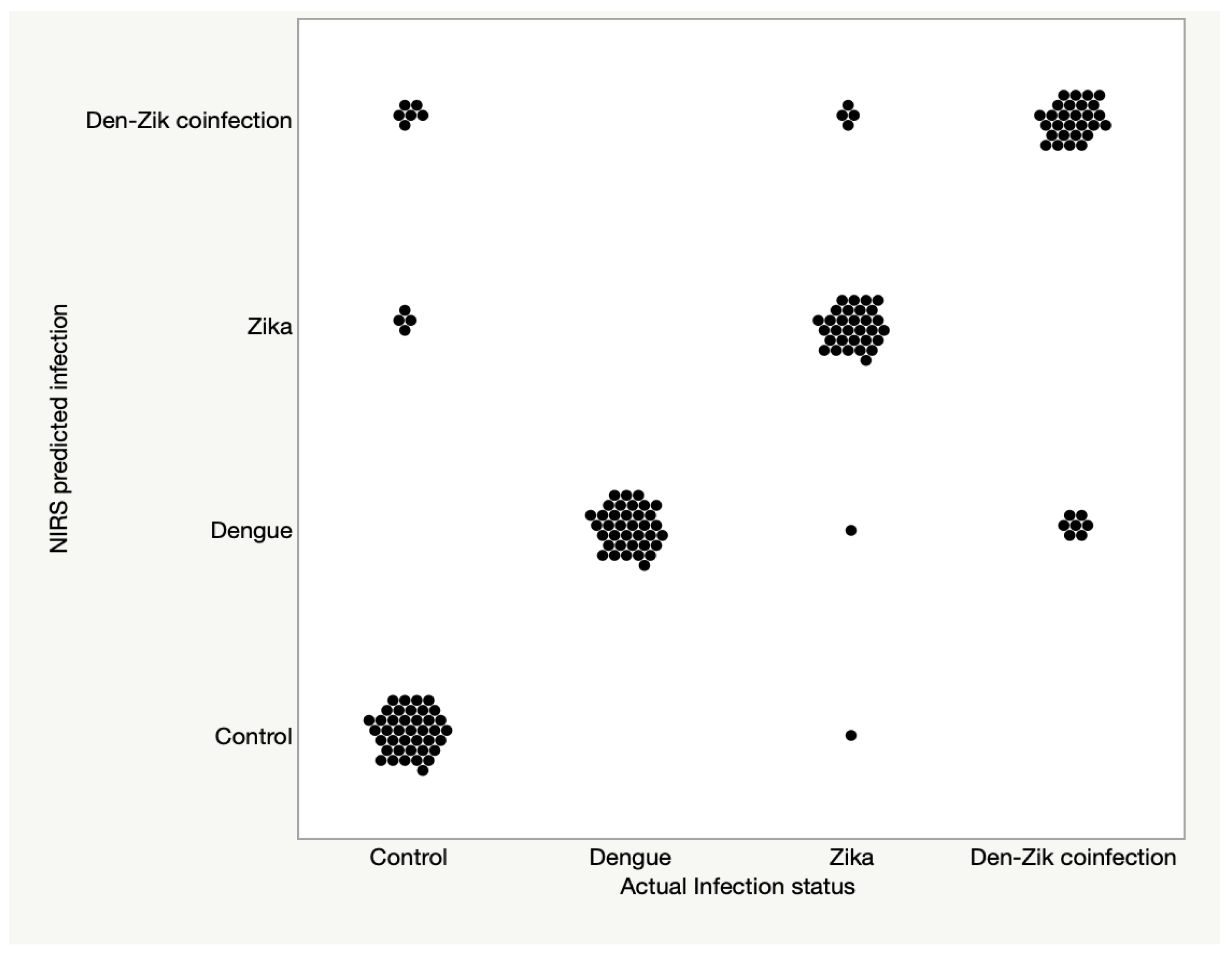
3.2. Differentiation between DENV, ZIKV, and ZIKV + DENV Coinfections in Ae. aegypti
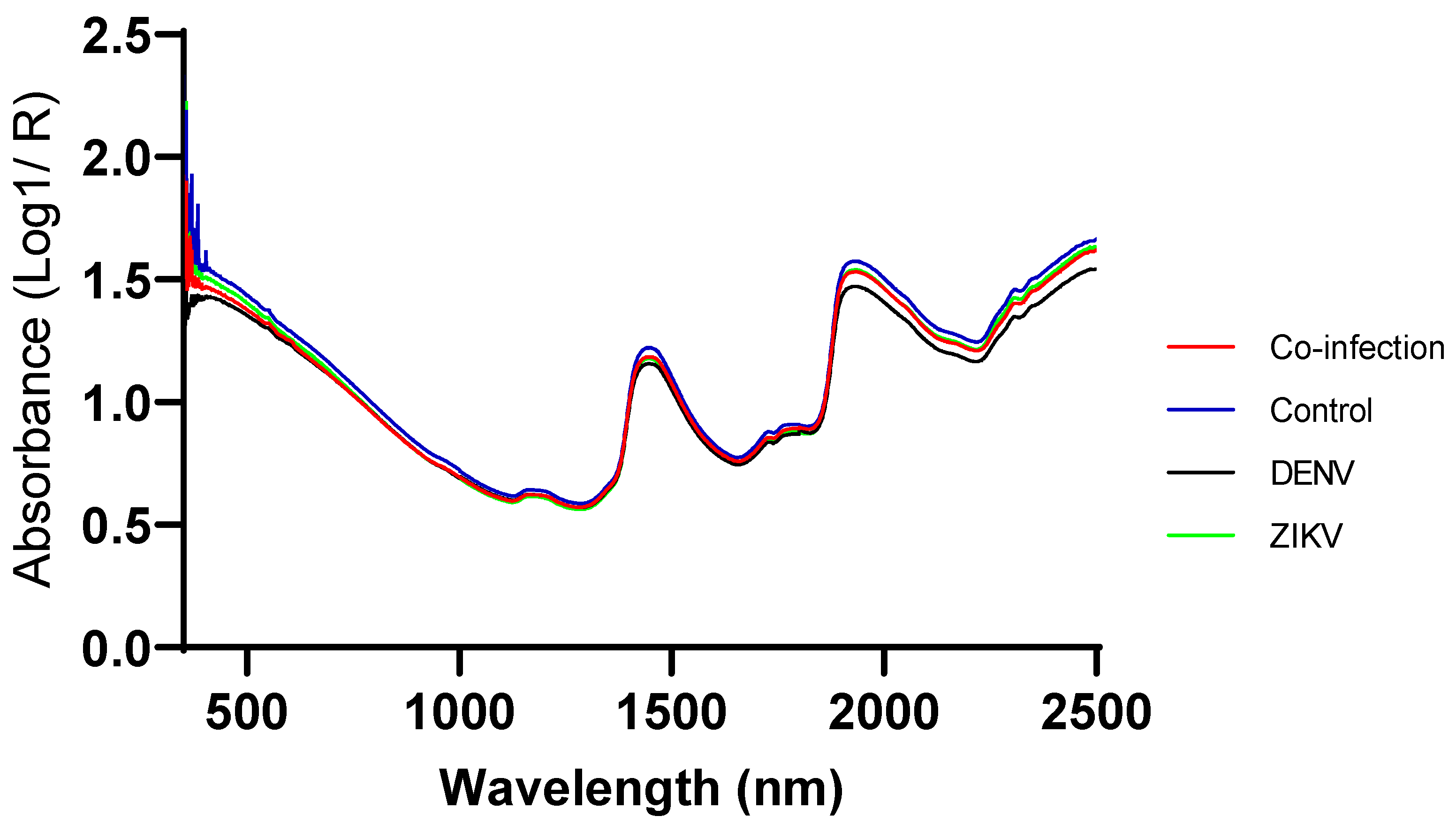
3.3. Second Derivative Figure of Infected and Uninfected Mosquitoes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How Aedes Aegypti Mosquitoes Enabled the First Pandemic Arbovirus. Annu. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhralia, S.; Verma, M.; Gopirajan, S.; Dhanaraj, P.S.; Lal, R.; Mehla, N.; Kant, C.R. From Dengue to Zika: The Wide Spread of Mosquito-Borne Arboviruses. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Chala, B.; Hamde, F. Emerging and Re-Emerging Vector-Borne Infectious Diseases and the Challenges for Control: A Review. Front. Public Health 2021, 9, 715759. [Google Scholar] [CrossRef]
- Edman, J.D.; Strickman, D.; Kittayapong, P.; Scott, T.W. Female Aedes Aegypti (Diptera: Culicidae) in Thailand Rarely Feed on Sugar. J. Med. Entomol. 1992, 29, 1035–1038. [Google Scholar] [CrossRef]
- David, M.R.; Dantas, E.S.; Maciel-de-Freitas, R.; Codeço, C.T.; Prast, A.E.; Lourenço-de-Oliveira, R. Influence of Larval Habitat Environmental Characteristics on Culicidae Immature Abundance and Body Size of Adult Aedes Aegypti. Front. Ecol. Evol. 2021, 9, 626757. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Marques, W.A.; Peres, R.C.; Cunha, S.P.; Lourenço-de-Oliveira, R. Variation in Aedes Aegypti(Diptera: Culicidae) Container Productivity in a Slum and a Suburban District of Rio de Janeiro during Dry and Wet Seasons. Mem. Inst. Oswaldo Cruz 2007, 102, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Abílio, A.P.; Abudasse, G.; Kampango, A.; Candrinho, B.; Sitoi, S.; Luciano, J.; Tembisse, D.; Sibindy, S.; de Almeida, A.P.G.; Garcia, G.A.; et al. Distribution and Breeding Sites of Aedes Aegypti and Aedes Albopictus in 32 Urban/Peri-Urban Districts of Mozambique: Implication for Assessing the Risk of Arbovirus Outbreaks. PLoS Negl. Trop. Dis. 2018, 12, e0006692. [Google Scholar] [CrossRef] [Green Version]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.-M.; Lourenco-De-Oliveira, R.; Caro, V.; Lambrechts, L.; Failloux, A.-B. Three-Way Interactions between Mosquito Population, Viral Strain and Temperature Underlying Chikungunya Virus Transmission Potential. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141078. [Google Scholar] [CrossRef]
- González, M.A.; Pavan, M.G.; Fernandes, R.S.; Busquets, N.; David, M.R.; Lourenço-Oliveira, R.; García-Pérez, A.L.; Maciel-de-Freitas, R. Limited Risk of Zika Virus Transmission by Five Aedes Albopictus Populations from Spain. Parasit. Vectors 2019, 12, 150. [Google Scholar] [CrossRef]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenço-de-Oliveira, R.; Failloux, A.-B. Differential Susceptibilities of Aedes Aegypti and Aedes Albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- De Filipps, A.M.B.; Vazeille, M.; Lourenço-de-Oliveira, R.; Failloux, A.-B. Large Genetic Differentiation and Low Variation in Vector Competence for Dengue and Yellow Fever Viruses of Aedes Albopictus from Brazil, the United States, and the Cayman Islands. Am. J. Trop. Med. Hyg. 2003, 69, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Pereira dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Aedes Albopictus as a Bridge Vector for Enzootic Pathogens at the Urban-Forest Interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes Management for the Control of Aedes-Borne Diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef] [Green Version]
- Olliaro, P.; Fouque, F.; Kroeger, A.; Bowman, L.; Velayudhan, R.; Santelli, A.C.; Garcia, D.; Skewes Ramm, R.; Sulaiman, L.H.; Tejeda, G.S.; et al. Improved Tools and Strategies for the Prevention and Control of Arboviral Diseases: A Research-to-Policy Forum. PLoS Negl. Trop. Dis. 2018, 12, e0005967. [Google Scholar] [CrossRef]
- Ramírez, A.L.; van den Hurk, A.F.; Meyer, D.B.; Ritchie, S.A. Searching for the Proverbial Needle in a Haystack: Advances in Mosquito-Borne Arbovirus Surveillance. Parasit. Vectors 2018, 11, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Leandro, A.S.; Lopes, R.D.; Martins, C.A.; Rivas, A.V.; da Silva, I.; Galvão, S.R.; Maciel-de-Freitas, R. The Adoption of the One Health Approach to Improve Surveillance of Venomous Animal Injury, Vector-Borne and Zoonotic Diseases in Foz Do Iguaçu, Brazil. PLoS Negl. Trop. Dis. 2021, 15, e0009109. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.S.; de Castro, W.A.C.; Lopes, R.D.; Delai, R.M.; Villela, D.A.M.; De-Freitas, R.M. Citywide Integrated Aedes Aegypti Mosquito Surveillance as Early Warning System for Arbovirus Transmission, Brazil. Emerg. Infect. Dis. 2022, 28, 701–706. [Google Scholar] [CrossRef]
- Zanluca, C.; De Melo, V.C.A.; Mosimann, A.L.P.; Dos Santos, G.I.V.; dos Santos, C.N.D.; Luz, K. First Report of Autochthonous Transmission of Zika Virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Nunes, M.R.T.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.G.; de Oliveira, L.F.; Azevedo, R.d.S.d.S.; da Silva, D.E.A.; da Silva, E.V.P.; da Silva, S.P.; et al. Emergence and Potential for Spread of Chikungunya Virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Novak, R.J. Short Report: Detection Probability of Arbovirus Infection in Mosquitopopulations. Am. J. Trop. Med. Hyg. 2004, 71, 636–638. [Google Scholar] [CrossRef]
- Juliano, S.A.; Ribeiro, G.S.; Maciel-de-Freitas, R.; Castro, M.G.; Codeço, C.; Lourenço-de-Oliveira, R.; Lounibos, L.P. She’s a Femme Fatale: Low-Density Larval Development Produces Good Disease Vectors. Mem. Inst. Oswaldo Cruz 2014, 109, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-de-Brito, A.; Ribeiro, I.P.; de Miranda, R.M.; Fernandes, R.S.; Campos, S.S.; da Silva, K.A.B.; de Castro, M.G.; Bonaldo, M.C.; Brasil, P.; Lourenço-de-Oliveira, R. First Detection of Natural Infection of Aedes Aegypti with Zika Virus in Brazil and throughout South America. Mem. Inst. Oswaldo Cruz 2016, 111, 655–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciel-de-Freitas, R.; Avendanho, F.C.; Santos, R.; Sylvestre, G.; Araújo, S.C.; Lima, J.B.P.; Martins, A.J.; Coelho, G.E.; Valle, D. Undesirable Consequences of Insecticide Resistance Following Aedes Aegypti Control Activities Due to a Dengue Outbreak. PLoS ONE 2014, 9, e92424. [Google Scholar] [CrossRef]
- Garcia-Rejon, J.; Loroño-Pino, M.A.; Farfan-Ale, J.A.; Flores-Flores, L.; Rosado-Paredes, E.D.P.; Rivero-Cardenas, N.; Najera-Vazquez, R.; Gomez-Carro, S.; Lira-Zumbardo, V.; Gonzalez-Martinez, P.; et al. Dengue Virus–Infected Aedes Aegypti in the Home Environment. Am. J. Trop. Med. Hyg. 2008, 79, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.; Ching, K.; Soares Magalhães, R.J.; Ciocchetta, S.; Edstein, M.D.; Maciel-de-Freitas, R.; Sikulu-Lord, M.T. The Application of Spectroscopy Techniques for Diagnosis of Malaria Parasites and Arboviruses and Surveillance of Mosquito Vectors: A Systematic Review and Critical Appraisal of Evidence. PLoS Negl. Trop. Dis. 2021, 15, e0009218. [Google Scholar] [CrossRef]
- Mayagaya, V.S.; Michel, K.; Benedict, M.Q.; Killeen, G.F.; Wirtz, R.A.; Ferguson, H.M.; Dowell, F.E. Non-Destructive Determination of Age and Species of Anopheles Gambiae s.l. Using Near-Infrared Spectroscopy. Am. J. Trop. Med. Hyg. 2009, 81, 622–630. [Google Scholar] [CrossRef]
- Sikulu, M.; Killeen, G.F.; Hugo, L.E.; Ryan, P.A.; Dowell, K.M.; Wirtz, R.A.; Moore, S.J.; Dowell, F.E. Near-Infrared Spectroscopy as a Complementary Age Grading and Species Identification Tool for African Malaria Vectors. Parasit. Vectors 2010, 3, 49. [Google Scholar] [CrossRef] [Green Version]
- Ntamatungiro, A.J.; Mayagaya, V.S.; Rieben, S.; Moore, S.J.; Dowell, F.E.; Maia, M.F. The Influence of Physiological Status on Age Prediction of Anopheles Arabiensis Using near Infra-Red Spectroscopy. Parasit. Vectors 2013, 6, 298. [Google Scholar] [CrossRef] [Green Version]
- Sikulu, M.; Dowell, K.M.; Hugo, L.E.; Wirtz, R.A.; Michel, K.; Peiris, K.H.; Moore, S.; Killeen, G.F.; Dowell, F.E. Evaluating RNAlater® as a Preservative for Using Near-Infrared Spectroscopy to Predict Anopheles Gambiae Age and Species. Malar. J. 2011, 10, 186. [Google Scholar] [CrossRef]
- Sikulu, M.T.; Majambere, S.; Khatib, B.O.; Ali, A.S.; Hugo, L.E.; Dowell, F.E. Using a Near-Infrared Spectrometer to Estimate the Age of Anopheles Mosquitoes Exposed to Pyrethroids. PLoS ONE 2014, 9, e90657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.N.; dos Santos, L.M.B.; Chouin-Carneiro, T.; Pavan, M.G.; Garcia, G.A.; David, M.R.; Beier, J.C.; Dowell, F.E.; Maciel-de-Freitas, R.; Sikulu-Lord, M.T. Rapid, Noninvasive Detection of Zika Virus in Aedes Aegypti Mosquitoes by near-Infrared Spectroscopy. Sci. Adv. 2018, 4, eaat0496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milali, M.P.; Sikulu-Lord, M.T.; Kiware, S.S.; Dowell, F.E.; Povinelli, R.J.; Corliss, G.F. Do NIR Spectra Collected from Laboratory-Reared Mosquitoes Differ from Those Collected from Wild Mosquitoes? PLoS ONE 2018, 13, e0198245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milali, M.P.; Sikulu-Lord, M.T.; Kiware, S.S.; Dowell, F.E.; Corliss, G.F.; Povinelli, R.J. Age Grading An. Gambiae and An. Arabiensis Using near Infrared Spectra and Artificial Neural Networks. PLoS ONE 2019, 14, e0209451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikulu-Lord, M.T.; Milali, M.P.; Henry, M.; Wirtz, R.A.; Hugo, L.E.; Dowell, F.E.; Devine, G.J. Near-Infrared Spectroscopy, a Rapid Method for Predicting the Age of Male and Female Wild-Type and Wolbachia Infected Aedes Aegypti. PLoS Negl. Trop. Dis. 2016, 10, e0005040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikulu-Lord, M.T.; Maia, M.F.; Milali, M.P.; Henry, M.; Mkandawile, G.; Kho, E.A.; Wirtz, R.A.; Hugo, L.E.; Dowell, F.E.; Devine, G.J. Rapid and Non-Destructive Detection and Identification of Two Strains of Wolbachia in Aedes Aegypti by Near-Infrared Spectroscopy. PLoS Negl. Trop. Dis. 2016, 10, e0004759. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.M.B.; Mutsaers, M.; Garcia, G.A.; David, M.R.; Pavan, M.G.; Petersen, M.T.; Corrêa-Antônio, J.; Couto-Lima, D.; Maes, L.; Dowell, F.; et al. High Throughput Estimates of Wolbachia, Zika and Chikungunya Infection in Aedes Aegypti by near-Infrared Spectroscopy to Improve Arbovirus Surveillance. Commun. Biol. 2021, 4, 67. [Google Scholar] [CrossRef]
- Esperança, P.M.; Blagborough, A.M.; Da, D.F.; Dowell, F.E.; Churcher, T.S. Detection of Plasmodium Berghei Infected Anopheles Stephensi Using Near-Infrared Spectroscopy. Parasit. Vectors 2018, 11, 377. [Google Scholar] [CrossRef] [Green Version]
- Da, D.F.; McCabe, R.; Somé, B.M.; Esperança, P.M.; Sala, K.A.; Blight, J.; Blagborough, A.M.; Dowell, F.; Yerbanga, S.R.; Lefèvre, T.; et al. Detection of Plasmodium Falciparum in Laboratory-Reared and Naturally Infected Wild Mosquitoes Using near-Infrared Spectroscopy. Sci. Rep. 2021, 11, 10289. [Google Scholar] [CrossRef]
- Maia, M.F.; Kapulu, M.; Muthui, M.; Wagah, M.G.; Ferguson, H.M.; Dowell, F.E.; Baldini, F.; Ranford-Cartwright, L. Detection of Plasmodium Falciparum Infected Anopheles Gambiae Using Near-Infrared Spectroscopy. Malar. J. 2019, 18, 85. [Google Scholar] [CrossRef]
- Dowell, F.E.; Noutcha, A.E.M.; Michel, K. The Effect of Preservation Methods on Predicting Mosquito Age by Near Infrared Spectroscopy. Am. J. Trop. Med. Hyg. 2011, 85, 1093–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayagaya, V.; Ntamatungiro, A.; Moore, S.; Wirtz, R.; Dowell, F.; Maia, M. Evaluating Preservation Methods for Identifying Anopheles Gambiae s.s. and Anopheles Arabiensis Complex Mosquitoes Species Using near Infra-Red Spectroscopy. Parasit. Vectors 2015, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaldo, M.C.; Ribeiro, I.P.; Lima, N.S.; dos Santos, A.A.C.; Menezes, L.S.R.; da Cruz, S.O.D.; de Mello, I.S.; Furtado, N.D.; de Moura, E.E.; Damasceno, L.; et al. Isolation of Infective Zika Virus from Urine and Saliva of Patients in Brazil. PLoS Negl. Trop. Dis. 2016, 10, e0004816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caragata, E.P.; Rocha, M.N.; Pereira, T.N.; Mansur, S.B.; Dutra, H.L.C.; Moreira, L.A. Pathogen Blocking in Wolbachia-Infected Aedes Aegypti Is Not Affected by Zika and Dengue Virus Co-Infection. PLoS Negl. Trop. Dis. 2019, 13, e0007443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, D.M.; Burke, D.S.; Harrison, B.A.; Whitmire, R.E.; Nisalak, A. Effect of Temperature on the Vector Efficiency of Aedes Aegypti for Dengue 2 Virus. Am. J. Trop. Med. Hyg. 1987, 36, 143–152. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Santos, M.C.D.; Viana, J.L.S.; Monteiro, J.D.; Freire, R.C.M.; Freitas, D.L.D.; Câmara, I.M.; Silva, G.J.S.; Gama, R.A.; Araújo, J.M.G.; Lima, K.M.G. Infrared spectroscopy (NIRS and ATR-FTIR) together with multivariate classification for non-destructive differentiation between female mosquitoes of Aedes aegypti recently infected with dengue vs. uninfected females. Acta Trop 2022, 235, 106633. [Google Scholar] [CrossRef]
- Lowe, R.; Barcellos, C.; Brasil, P.; Cruz, O.; Honório, N.; Kuper, H.; Carvalho, M. The Zika Virus Epidemic in Brazil: From Discovery to Future Implications. Int. J. Environ. Res. Public Health 2018, 15, 96. [Google Scholar] [CrossRef] [Green Version]
- Kazazian, L.; Lima Neto, A.S.; Sousa, G.S.; do Nascimento, O.J.; Castro, M.C. Spatiotemporal Transmission Dynamics of Co-Circulating Dengue, Zika, and Chikungunya Viruses in Fortaleza, Brazil: 2011–2017. PLoS Negl. Trop. Dis. 2020, 14, e0008760. [Google Scholar] [CrossRef]
- Nunes, P.C.G.; Daumas, R.P.; Sánchez-Arcila, J.C.; Nogueira, R.M.R.; Horta, M.A.P.; dos Santos, F.B. 30 Years of Fatal Dengue Cases in Brazil: A Review. BMC Public Health 2019, 19, 329. [Google Scholar] [CrossRef]
- Ciota, A.T. The Role of Co-Infection and Swarm Dynamics in Arbovirus Transmission. Virus Res. 2019, 265, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.; Paupy, C.; Grard, G.; Becquart, P.; Mombo, I.; Nso, B.B.B.; Kassa Kassa, F.; Nkoghe, D.; Leroy, E.M. Recent Introduction and Rapid Dissemination of Chikungunya Virus and Dengue Virus Serotype 2 Associated with Human and Mosquito Coinfections in Gabon, Central Africa. Clin. Infect. Dis. 2012, 55, e45–e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito Co-Infection with Zika and Chikungunya Virus Allows Simultaneous Transmission without Affecting Vector Competence of Aedes Aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of Simultaneous Exposure to Arboviruses on Infection and Transmission by Aedes Aegypti Mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [Green Version]
- Kantor, A.M.; Lin, J.; Wang, A.; Thompson, D.C.; Franz, A.W.E. Infection Pattern of Mayaro Virus in Aedes Aegypti (Diptera: Culicidae) and Transmission Potential of the Virus in Mixed Infections with Chikungunya Virus. J. Med. Entomol. 2019, 56, 832–843. [Google Scholar] [CrossRef]
- Le Coupanec, A.; Tchankouo-Nguetcheu, S.; Roux, P.; Khun, H.; Huerre, M.; Morales-Vargas, R.; Enguehard, M.; Lavillette, D.; Missé, D.; Choumet, V. Co-Infection of Mosquitoes with Chikungunya and Dengue Viruses Reveals Modulation of the Replication of Both Viruses in Midguts and Salivary Glands of Aedes Aegypti Mosquitoes. Int. J. Mol. Sci. 2017, 18, 1708. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-H.; Huber, R.G.; Bond, P.J.; Grad, Y.H.; Camerini, D.; Maurer-Stroh, S.; Lipsitch, M. Systematic Analysis of Protein Identity between Zika Virus and Other Arthropod-Borne Viruses. Bull. World Health Organ. 2017, 95, 517–525I. [Google Scholar] [CrossRef]
- Angleró-Rodríguez, Y.I.; MacLeod, H.J.; Kang, S.; Carlson, J.S.; Jupatanakul, N.; Dimopoulos, G. Aedes Aegypti Molecular Responses to Zika Virus: Modulation of Infection by the Toll and Jak/Stat Immune Pathways and Virus Host Factors. Front. Microbiol. 2017, 8, 2050. [Google Scholar] [CrossRef] [Green Version]
- Murrieta, R.A.; Garcia-Luna, S.M.; Murrieta, D.J.; Halladay, G.; Young, M.C.; Fauver, J.R.; Gendernalik, A.; Weger-Lucarelli, J.; Rückert, C.; Ebel, G.D. Impact of Extrinsic Incubation Temperature on Natural Selection during Zika Virus Infection of Aedes Aegypti and Aedes Albopictus. PLOS Pathog. 2021, 17, e1009433. [Google Scholar] [CrossRef]
- Aw, W.C.; Dowell, F.E.; Ballard, J.W.O. Using Near-Infrared Spectroscopy to Resolve the Species, Gender, Age, and the Presence of Wolbachia Infection in Laboratory-Reared Drosophila. G3 Genes|Genomes|Genet. 2012, 2, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.T.; da Silveira, I.D.; Tátila-Ferreira, A.; David, M.R.; Chouin-Carneiro, T.; Van den Wouwer, L.; Maes, L.; Maciel-de-Freitas, R. The Impact of the Age of First Blood Meal and Zika Virus Infection on Aedes Aegypti Egg Production and Longevity. PLoS ONE 2018, 13, e0200766. [Google Scholar] [CrossRef] [PubMed]
- Maciel-de-Freitas, R.; Sylvestre, G.; Gandini, M.; Koella, J.C. The Influence of Dengue Virus Serotype-2 Infection on Aedes Aegypti (Diptera: Culicidae) Motivation and Avidity to Blood Feed. PLoS ONE 2013, 8, e65252. [Google Scholar] [CrossRef] [PubMed]
- Maciel-de-Freitas, R.; Koella, J.C.; Lourenço-de-Oliveira, R. Lower Survival Rate, Longevity and Fecundity of Aedes Aegypti (Diptera: Culicidae) Females Orally Challenged with Dengue Virus Serotype 2. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 452–458. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, I.D.; Petersen, M.T.; Sylvestre, G.; Garcia, G.A.; David, M.R.; Pavan, M.G.; Maciel-de-Freitas, R. Zika Virus Infection Produces a Reduction on Aedes Aegypti Lifespan but No Effects on Mosquito Fecundity and Oviposition Success. Front. Microbiol. 2018, 9, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, N.; Bhatti, A.; Paradkar, P.N. Advances in Understanding Vector Behavioural Traits after Infection. Pathogens 2021, 10, 1376. [Google Scholar] [CrossRef]
- Etebari, K.; Hegde, S.; Saldaña, M.A.; Widen, S.G.; Wood, T.G.; Asgari, S.; Hughes, G.L. Global Transcriptome Analysis of Aedes Aegypti Mosquitoes in Response to Zika Virus Infection. mSphere 2017, 2, 1648–1659. [Google Scholar] [CrossRef] [Green Version]
- Ramos, L.F.C.; Martins, M.; Murillo, J.R.; Domont, G.B.; de Oliveira, D.M.P.; Nogueira, F.C.S.; Maciel-de-Freitas, R.; Junqueira, M. Interspecies Isobaric Labeling-Based Quantitative Proteomics Reveals Protein Changes in the Ovary of Aedes Aegypti Coinfected with ZIKV and Wolbachia. Front. Cell. Infect. Microbiol. 2022, 12, 900608. [Google Scholar] [CrossRef]
- Martins, M.; Ramos, L.F.C.; Murillo, J.R.; Torres, A.; de Carvalho, S.S.; Domont, G.B.; de Oliveira, D.M.P.; Mesquita, R.D.; Nogueira, F.C.S.; Maciel-de-Freitas, R.; et al. Comprehensive Quantitative Proteome Analysis of Aedes Aegypti Identifies Proteins and Pathways Involved in Wolbachia Pipientis and Zika Virus Interference Phenomenon. Front. Physiol. 2021, 12, 642237. [Google Scholar] [CrossRef]
- Ayllón, T.; Campos, R.D.M.; Brasil, P.; Morone, F.C.; Câmara, D.C.P.; Meira, G.L.S.; Tannich, E.; Yamamoto, K.A.; Carvalho, M.S.; Pedro, R.S.; et al. Early Evidence for Zika Virus Circulation among Aedes Aegypti Mosquitoes, Rio de Janeiro, Brazil. Emerg. Infect. Dis. 2017, 23, 1411–1412. [Google Scholar] [CrossRef] [Green Version]
- dos Reis, I.C.; Gibson, G.; Ayllón, T.; de Medeiros Tavares, A.; de Araújo, J.M.G.; da Silva Monteiro, E.; Rodrigues Aguiar, A.; de Oliveira, J.V.; de Paiva, A.A.P.; Wana Bezerra Pereira, H.; et al. Entomo-Virological Surveillance Strategy for Dengue, Zika and Chikungunya Arboviruses in Field-Caught Aedes Mosquitoes in an Endemic Urban Area of the Northeast of Brazil. Acta Trop. 2019, 197, 105061. [Google Scholar] [CrossRef]
- Singh, J.; Balakrishnan, N.; Bhardwaj, M.; Amuthadevi, P.; George, E.G.; Subramani, K.; Soundararajan, K.; Appavoo, N.C.; Jain, D.C.; Ichhpujani, R.L.; et al. Silent Spread of Dengue and Dengue Haemorrhagic Fever to Coimbatore and Erode Districts in Tamil Nadu, India, 1998: Need for Effective Surveillance to Monitor and Control the Disease. Epidemiol. Infect. 2000, 125, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.S.; Lopes, R.D.; Amaral Martins, C.; Delai, R.M.; Villela, D.A.M.; Maciel-de-Freitas, R. Entomo-virological surveillance followed by serological active survey of symptomatic individuals is helpful to identify hotspots of early arbovirus transmission. Front. Public Health 2022, 10, 1024187. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | |||
|---|---|---|---|
| ZIKV-Infected | DENV-Infected | ZIKV/DENV Coinfected | |
| Infection rate | 90.62% (n = 87/96) | 90.62% (n = 87/96) | 96.6% (n = 85/88) |
| Training and Validation Set | Test Set | |||||||
|---|---|---|---|---|---|---|---|---|
| % Spec (n) | % Sensitivity (n) | % Spec (n) | % Sensitivity (n) | |||||
| Predicted Group | Control | DENV | ZIKV | Co-Infection ZIKV/DENV | Control | DENV | ZIKV | Co-Infection ZIKV/DENV |
| Predicted into actual infection group | 98 (n = 50) | 100 (n = 50) | 98 (n = 50) | 100 (n = 50) | 80 (n = 50) | 100 (n = 37) | 84 (n = 37) | 80 (n = 35) |
| Predicted as infected | 2 (n = 50) | 100 (n = 50) | 98 (n = 50) | 100 (n = 50) | 20 (n = 50) | 100 (n = 37) | 97 (n = 37) | 100 (n = 35) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, G.A.; Lord, A.R.; Santos, L.M.B.; Kariyawasam, T.N.; David, M.R.; Couto-Lima, D.; Tátila-Ferreira, A.; Pavan, M.G.; Sikulu-Lord, M.T.; Maciel-de-Freitas, R. Rapid and Non-Invasive Detection of Aedes aegypti Co-Infected with Zika and Dengue Viruses Using Near Infrared Spectroscopy. Viruses 2023, 15, 11. https://doi.org/10.3390/v15010011
Garcia GA, Lord AR, Santos LMB, Kariyawasam TN, David MR, Couto-Lima D, Tátila-Ferreira A, Pavan MG, Sikulu-Lord MT, Maciel-de-Freitas R. Rapid and Non-Invasive Detection of Aedes aegypti Co-Infected with Zika and Dengue Viruses Using Near Infrared Spectroscopy. Viruses. 2023; 15(1):11. https://doi.org/10.3390/v15010011
Chicago/Turabian StyleGarcia, Gabriela A., Anton R. Lord, Lilha M. B. Santos, Tharanga N. Kariyawasam, Mariana R. David, Dinair Couto-Lima, Aline Tátila-Ferreira, Márcio G. Pavan, Maggy T. Sikulu-Lord, and Rafael Maciel-de-Freitas. 2023. "Rapid and Non-Invasive Detection of Aedes aegypti Co-Infected with Zika and Dengue Viruses Using Near Infrared Spectroscopy" Viruses 15, no. 1: 11. https://doi.org/10.3390/v15010011