
A Novel Single-Stranded RNA-Based Adjuvant Improves the Immunogenicity of the SARS-CoV-2 Recombinant Protein Vaccine

Abstract
:1. Introduction
2. Materials and Methods
2.1. Biosafety and Ethics Statement
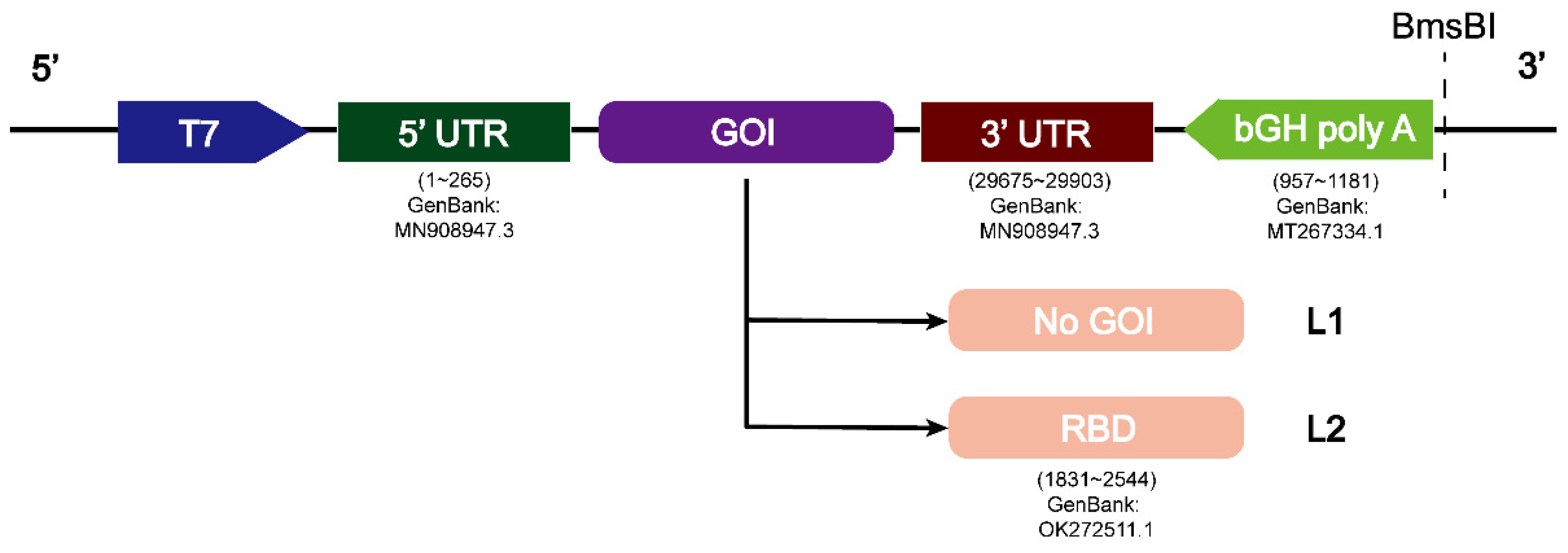
2.2. Preparation of RNA L1 (L1) and RNA L2 (L2)
2.3. Transfection
2.4. Luciferase Reporter Assay
2.5. Real-time Quantitative PCR (qPCR)
2.6. Western Blot (WB) Analysis
2.7. Vaccine Preparation
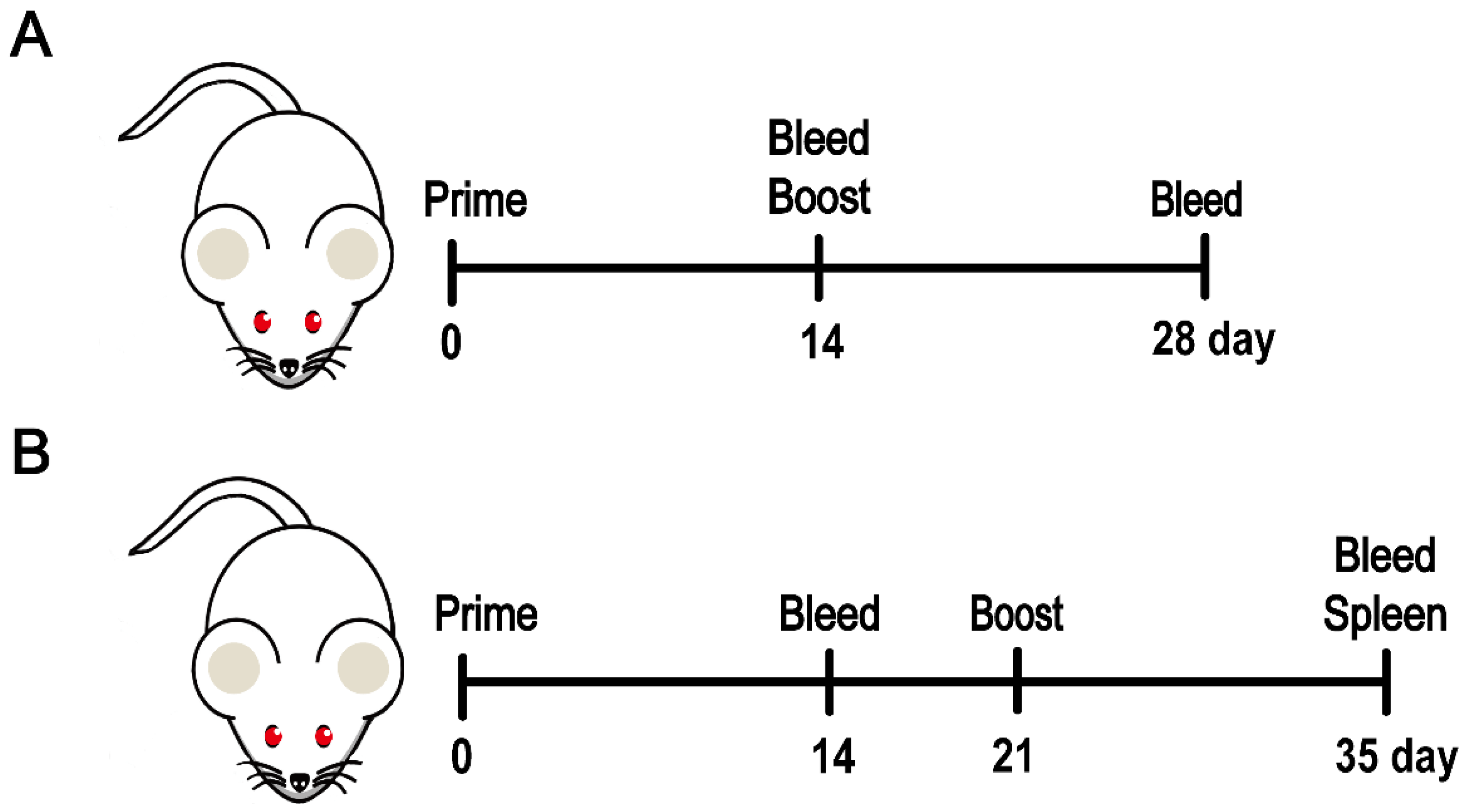
2.8. Animal Studies
2.9. Measurement of RBD-Specific IgG Titers Using ELISA
2.10. Serum Neutralization Assay
2.11. IFN-γ ELISPOT Assay
2.12. Cytokine Assay
2.13. Statistical Analyses
3. Results
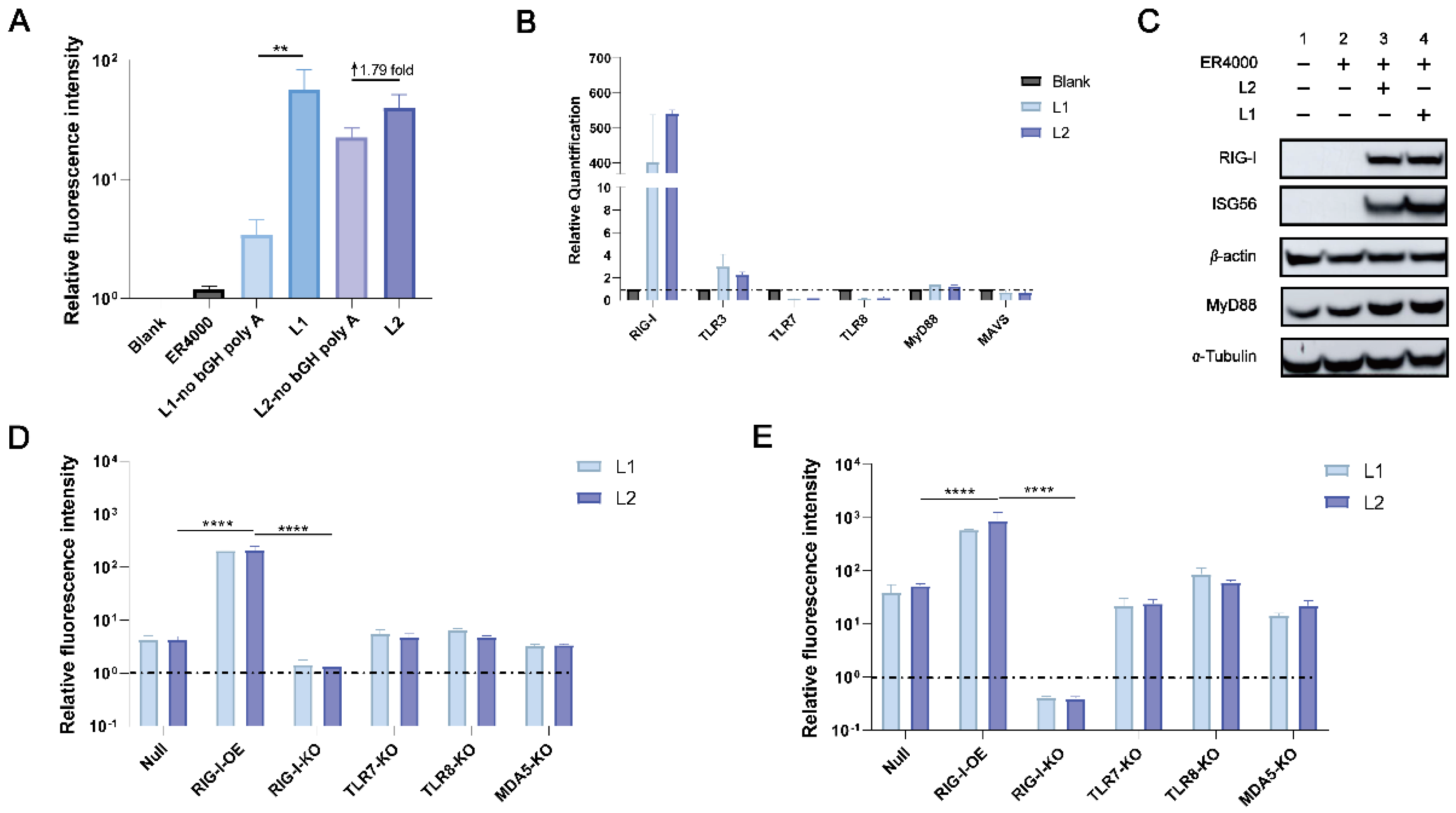
3.1. L1 and L2 Activated the RIG-I Signaling Pathways
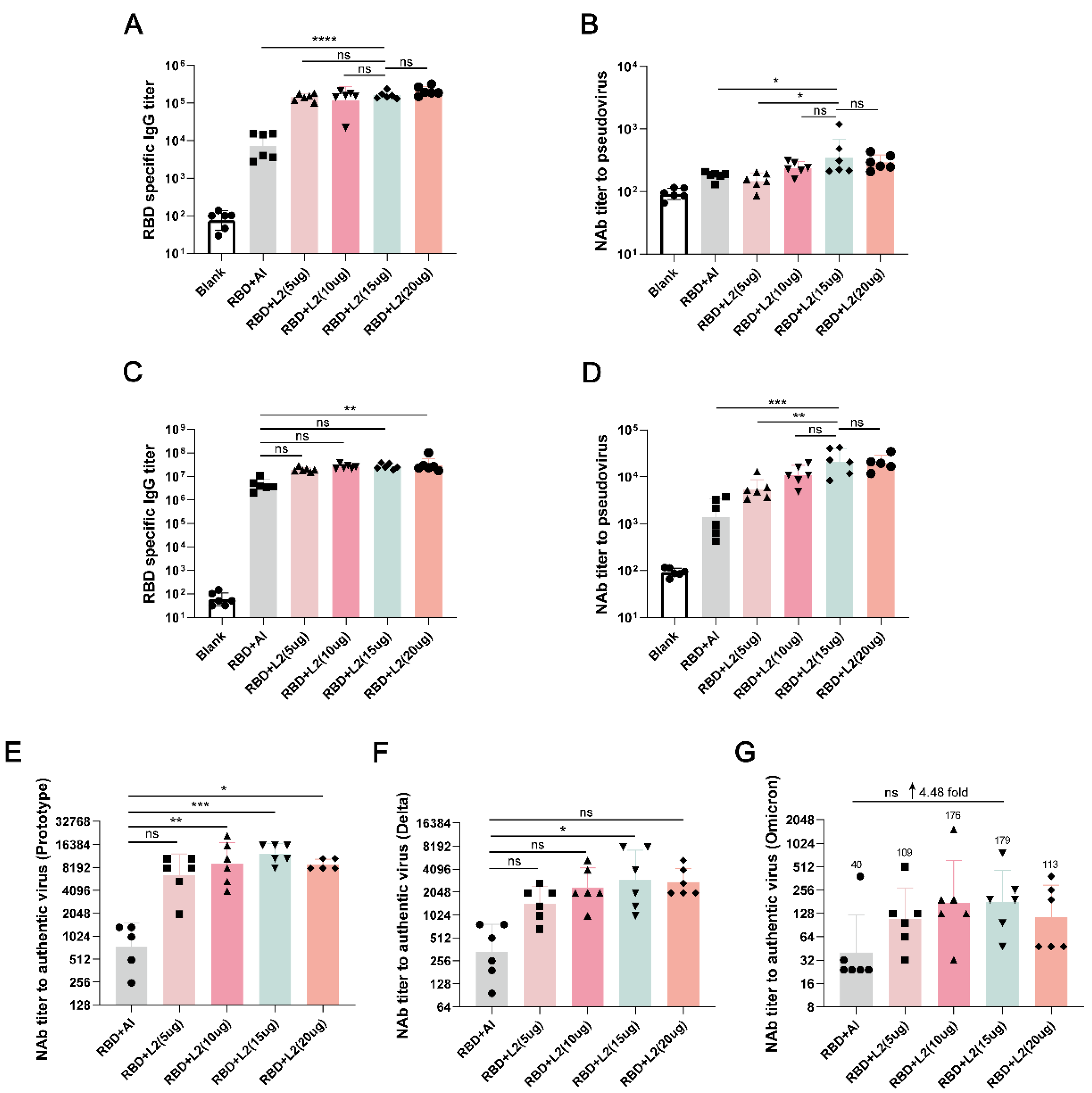
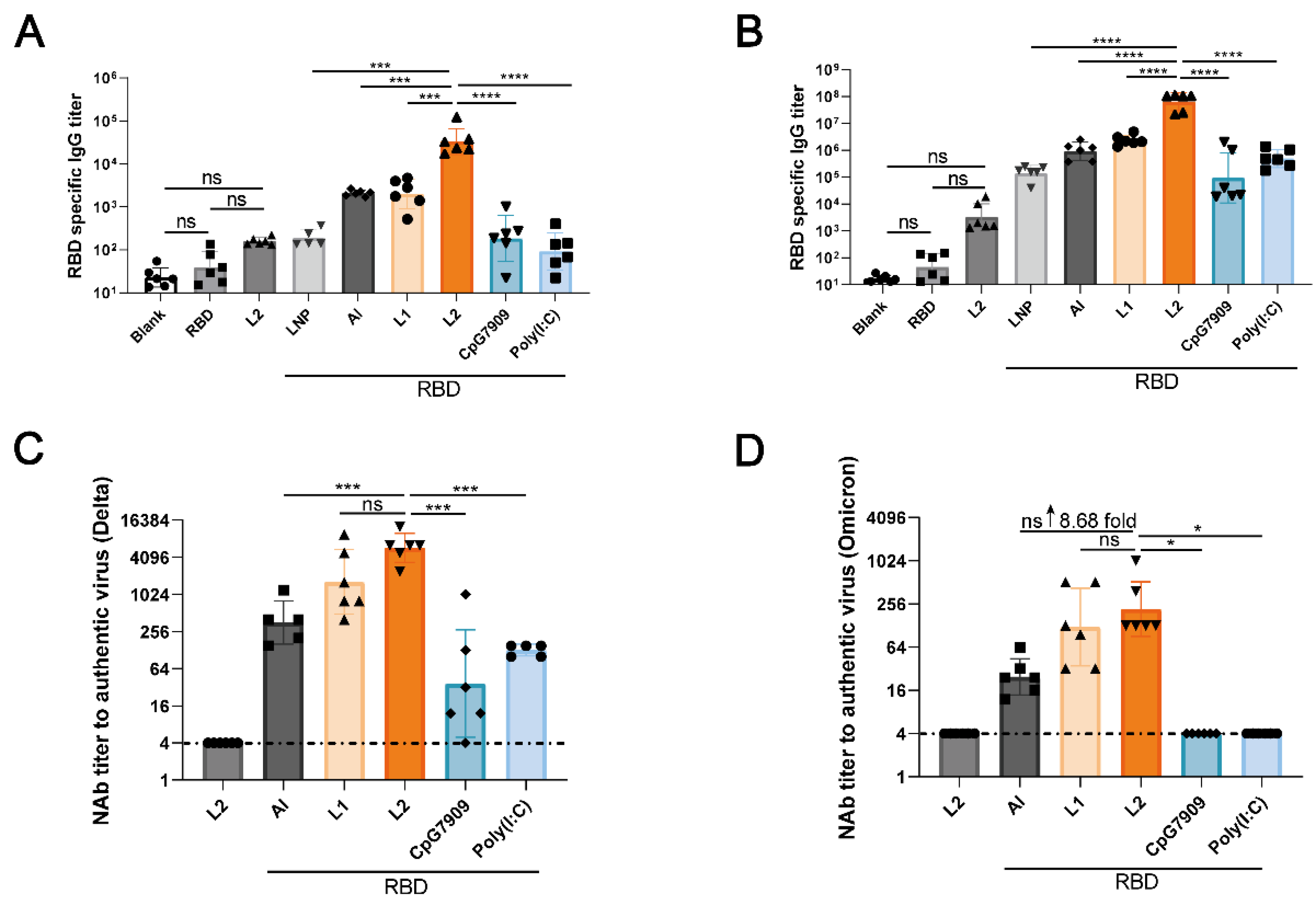
3.2. L2 Improved the Immunogenicity of RBD Compared with Al
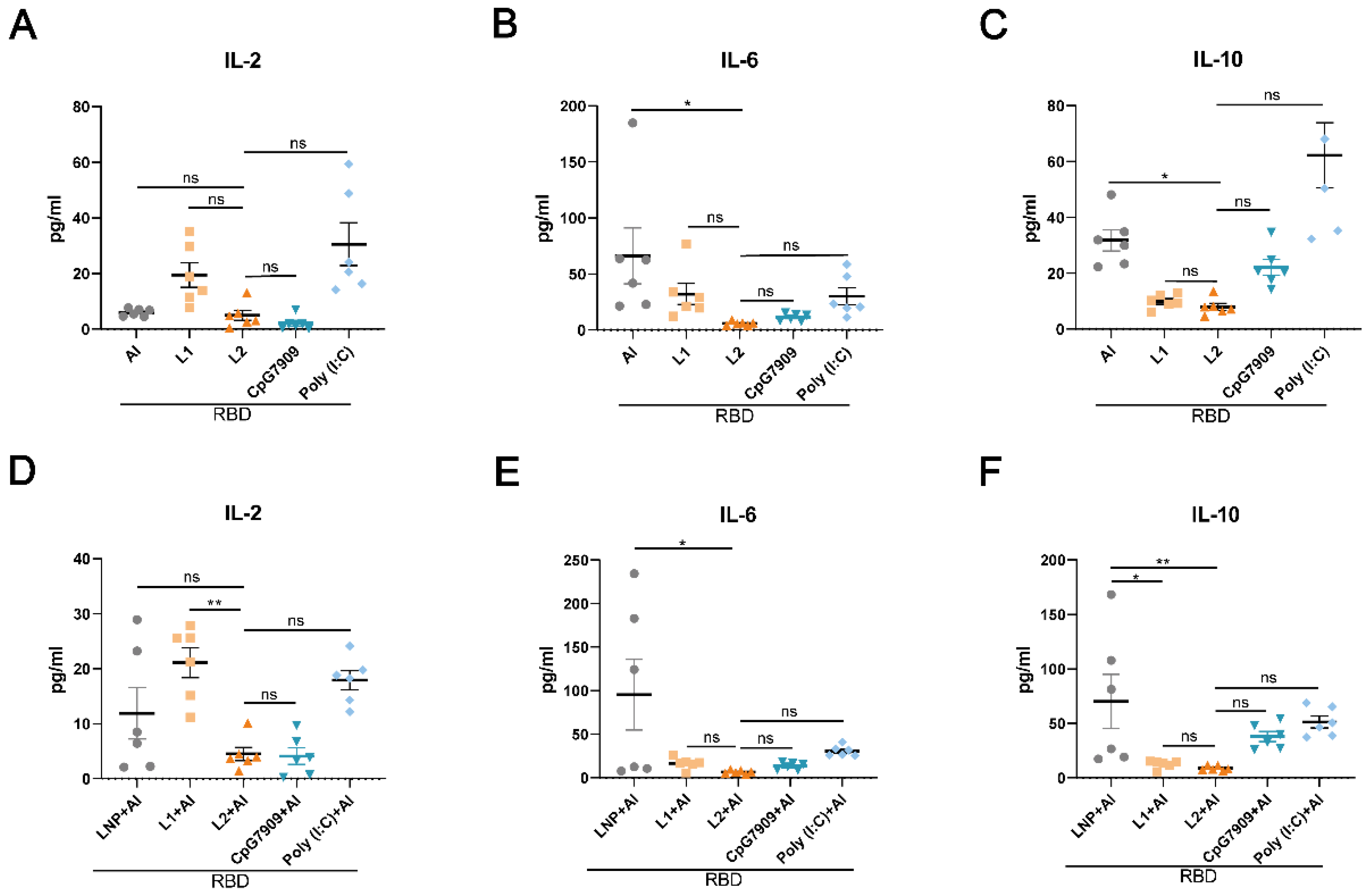
3.3. L2 Improved the Immunogenicity of RBD Compared with CpG 7909 and Poly(I:C)
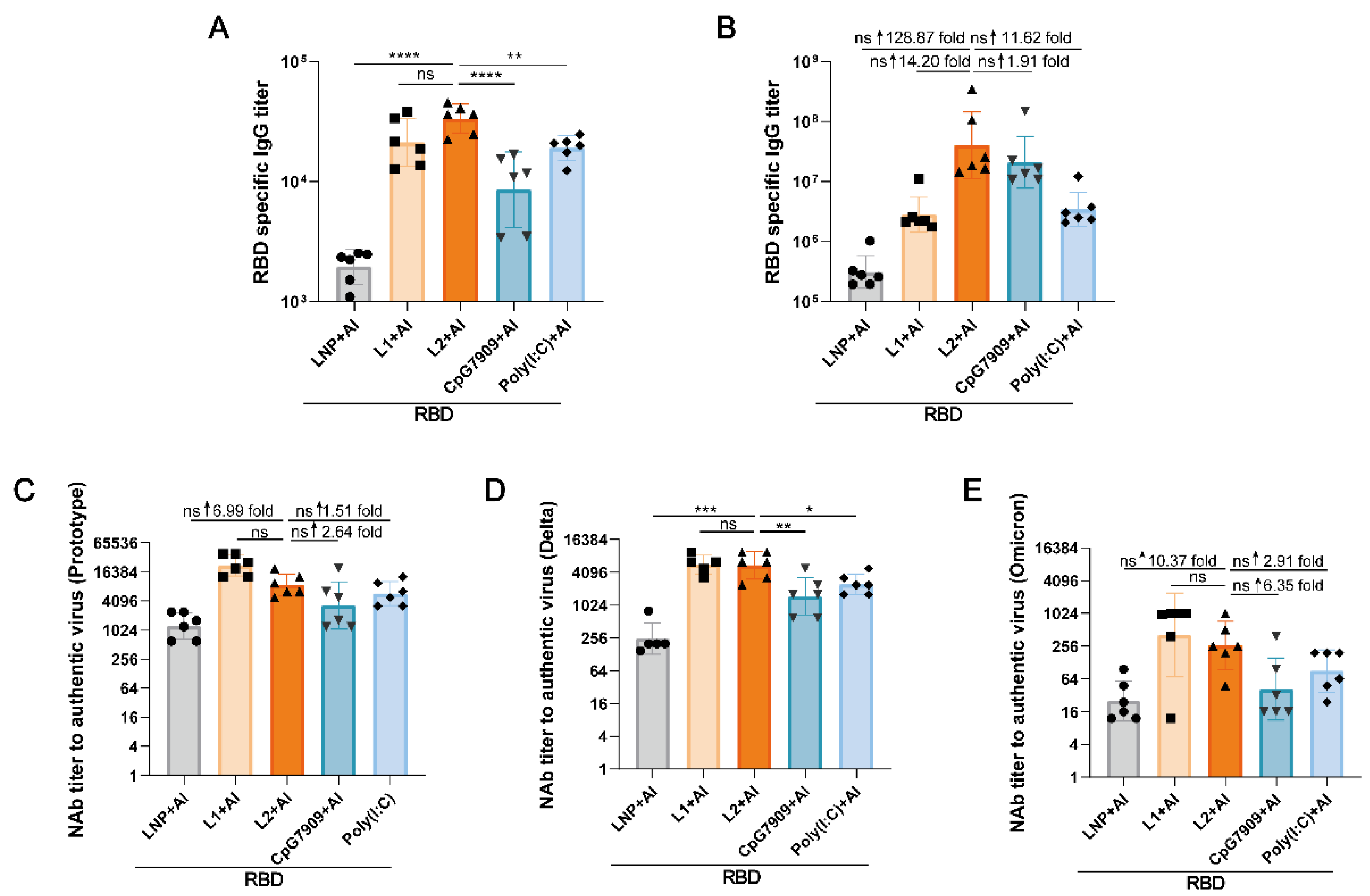
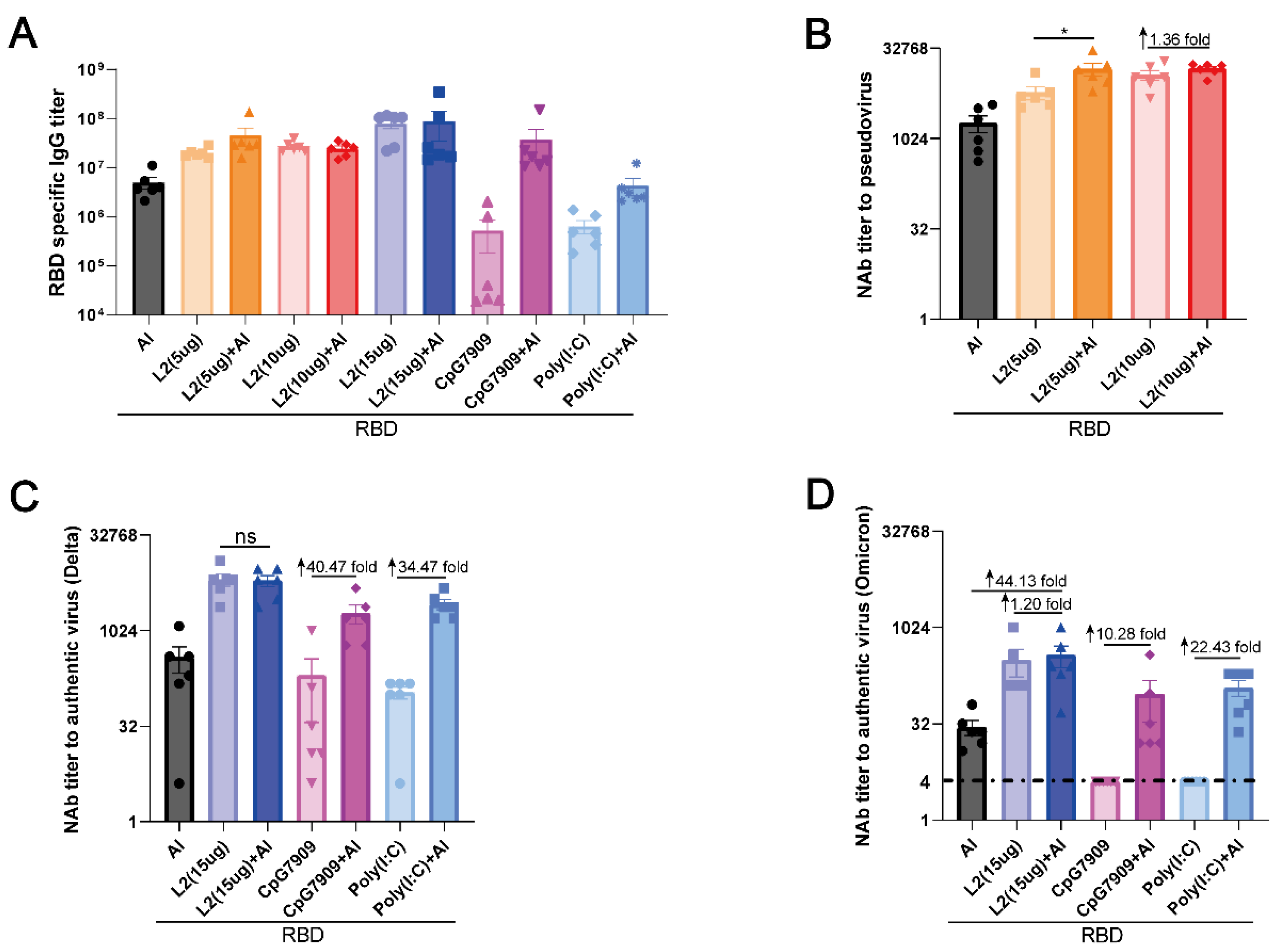
3.4. L2 plus Al Improved the Immunogenicity of RBD Compared with CpG 7909 plus Al and Poly(I:C) plus Al
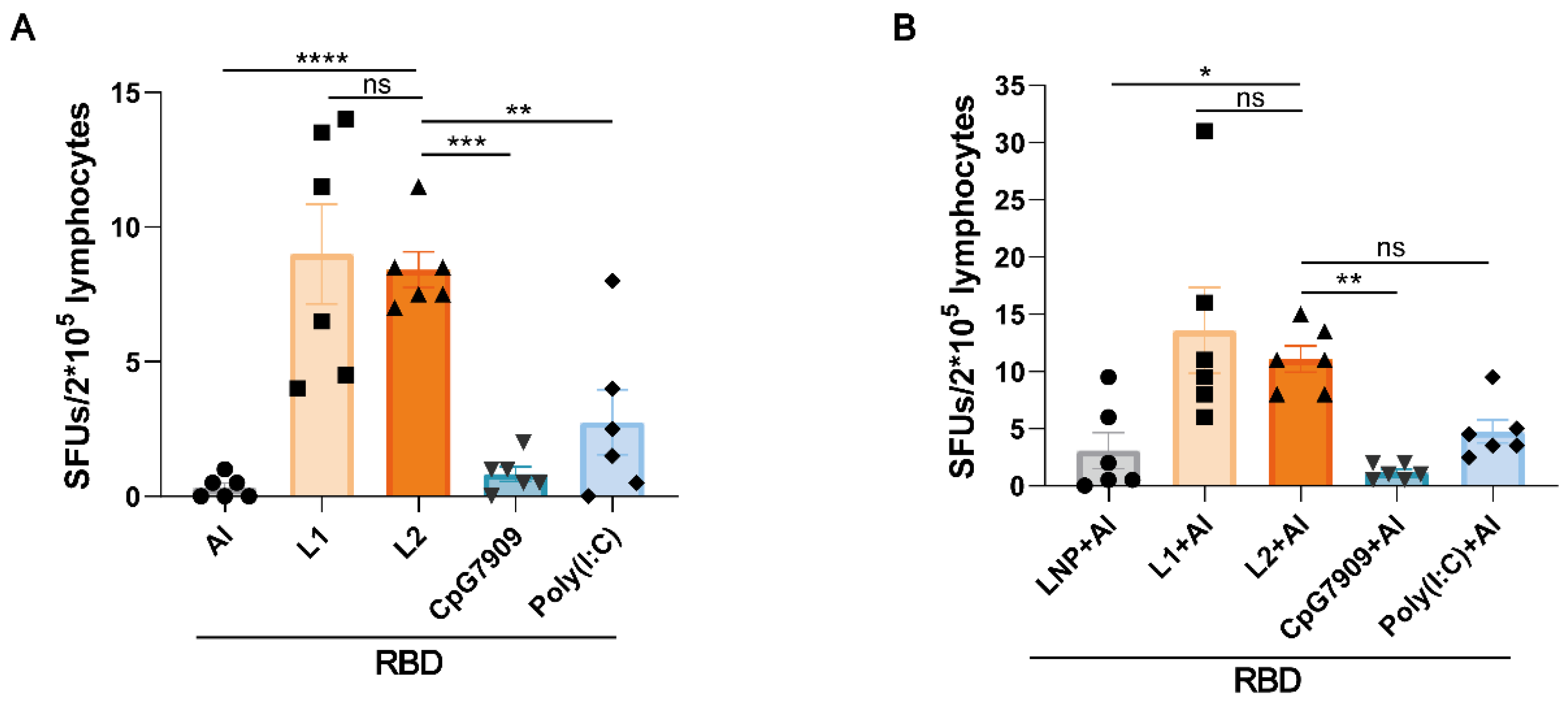
3.5. L2 and L2 plus Al Elevated the Level of RBD-Specific Th1 Cell Immune Response Represented by IFN-γ Secretion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://www.who.int/redirect-pages/page/novel-coronavirus-(covid-19)-situation-dashboard (accessed on 25 June 2022).
- National Health Commission of the Peoples’ Republic of China. Novel Coronavirus Vaccination Status in Chinese Mainland. Available online: http://www.nhc.gov.cn/xcs/yqfkdt/202206/84b28c26fc5b4859a14034fc8fd85c4b.shtml (accessed on 26 June 2022).
- Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 25 June 2022).
- An, Y.; Li, S.; Jin, X.; Han, J.B.; Xu, K.; Xu, S.; Han, Y.; Liu, C.; Zheng, T.; Liu, M.; et al. A tandem-repeat dimeric RBD protein-based covid-19 vaccine zf2001 protects mice and nonhuman primates. Emerg. Microbes Infect. 2022, 11, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Zheng, T.; Xu, K.; Han, Y.; Xu, L.; Huang, E.; An, Y.; Cheng, Y.; Li, S.; Liu, M.; et al. A Universal Design of Betacoronavirus Vaccines against COVID-19, MERS, and SARS. Cell 2020, 182, 722–733.e711. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Gao, P.; Liu, S.; Lu, S.; Lei, W.; Zheng, T.; Liu, X.; Xie, Y.; Zhao, Z.; Guo, S.; et al. Protective prototype-Beta and Delta-Omicron chimeric RBD-dimer vaccines against SARS-CoV-2. Cell 2022, 13, 2265–2278. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; P, S.A.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Mascola, J.R.; Fauci, A.S. Novel vaccine technologies for the 21st century. Nat. Rev. Immunol 2020, 20, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R.; Peachman, K.K.; Matyas, G.R.; Rao, M.; Beck, Z. Army Liposome Formulation (ALF) family of vaccine adjuvants. Expert Rev. Vaccines 2020, 19, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R.R.; Rasheed, M.A.; Bhaumik, S.K.; Ranjan, P.; Cao, W.; Davis, C.; Marisetti, K.; Thomas, S.; Gangappa, S.; Sambhara, S.; et al. Activation of the RIG-I pathway during influenza vaccination enhances the germinal center reaction, promotes T follicular helper cell induction, and provides a dose-sparing effect and protective immunity. J. Virol. 2014, 88, 13990–14001. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Singh, V.K.; Mishra, A.; Soudani, E.; Bakhru, P.; Singh, C.R.; Zhang, D.; Canaday, D.H.; Sheri, A.; Padmanabhan, S.; et al. NOD2/RIG-I Activating Inarigivir Adjuvant Enhances the Efficacy of BCG Vaccine Against Tuberculosis in Mice. Front. Immunol. 2020, 11, 592333. [Google Scholar] [CrossRef]
- Chiang, C.; Beljanski, V.; Yin, K.; Olagnier, D.; Ben, Y.F.; Steel, C.; Goulet, M.L.; DeFilippis, V.R.; Streblow, D.N.; Haddad, E.K.; et al. Sequence-Specific Modifications Enhance the Broad-Spectrum Antiviral Response Activated by RIG-I Agonists. J. Virol. 2015, 89, 8011–8025. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Zhang, S.; Yang, Z.; Lin, H.; Zhu, J.; Liu, L.; Wang, W.; Liu, S.; Liu, W.; Ma, Y.; et al. Self-Recognition of an Inducible Host lncRNA by RIG-I Feedback Restricts Innate Immune Response. Cell 2018, 173, 906–919.e913. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.L.; Angel, J.B.; Seguin, I.; Davis, H.L.; Cameron, D.W. CPG 7909 adjuvant plus hepatitis B virus vaccination in HIV-infected adults achieves long-term seroprotection for up to 5 years. Clin. Infect. Dis 2008, 46, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Bang, Y.J.; Park, H.J.; Li Ko, H.; Park, S.I.; Hwang, K.A.; Kim, H.; Nam, J.H. Inactivated influenza vaccine formulated with single-stranded RNA-based adjuvant confers mucosal immunity and cross-protection against influenza virus infection. Vaccine 2020, 38, 6141–6152. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Bang, E.K.; Hong, J.J.; Lee, S.M.; Ko, H.L.; Kwak, H.W.; Park, H.; Kang, K.W.; Kim, R.H.; Ryu, S.R.; et al. Nanoformulated Single-Stranded RNA-Based Adjuvant with a Coordinative Amphiphile as an Effective Stabilizer: Inducing Humoral Immune Response by Activation of Antigen-Presenting Cells. Angew. Chem. Int. Ed. Engl. 2020, 59, 11540–11549. [Google Scholar] [CrossRef] [PubMed]
- Bang, Y.J.; Hong, S.H.; Park, H.J.; Kwak, H.W.; Lee, Y.S.; Kim, J.Y.; Park, H.J.; Bae, S.H.; Kim, H.J.; Kim, Y.H.; et al. Effective inactivated influenza vaccine for the elderly using a single-stranded RNA-based adjuvant. Sci Rep. 2021, 11, 11981. [Google Scholar] [CrossRef] [PubMed]
- Thorne, L.G.; Reuschl, A.K.; Zuliani-Alvarez, L.; Whelan, M.V.X.; Turner, J.; Noursadeghi, M.; Jolly, C.; Towers, G.J. SARS-CoV-2 sensing by RIG-I and MDA5 links epithelial infection to macrophage inflammation. EMBO J. 2021, 40, e107826. [Google Scholar] [CrossRef]
- Kouwaki, T.; Nishimura, T.; Wang, G.; Oshiumi, H. RIG-I-Like Receptor-Mediated Recognition of Viral Genomic RNA of Severe Acute Respiratory Syndrome Coronavirus-2 and Viral Escape From the Host Innate Immune Responses. Front. Immunol. 2021, 12, 700926. [Google Scholar] [CrossRef]
- Mao, T.; Israelow, B.; Lucas, C.; Vogels, C.B.F.; Gomez-Calvo, M.L.; Fedorova, O.; Breban, M.I.; Menasche, B.L.; Dong, H.; Linehan, M.; et al. A stem-loop RNA RIG-I agonist protects against acute and chronic SARS-CoV-2 infection in mice. J. Exp. Med. 2022, 219, e20211818. [Google Scholar] [CrossRef]
- Apostolico, J.S.; Lunardelli, V.A.S.; Yamamoto, M.M.; Cunha-Neto, E.; Boscardin, S.B.; Rosa, D.S. Poly(I:C) Potentiates T Cell Immunity to a Dendritic Cell Targeted HIV-Multiepitope Vaccine. Front. Immunol 2019, 10, 843. [Google Scholar] [CrossRef]
- Hu, S.; Chen, H.; Ma, J.; Chen, Q.; Deng, H.; Gong, F.; Huang, H.; Shi, C. CpG7909 adjuvant enhanced immunogenicity efficacy in mice immunized with ESAT6-Ag85A fusion protein, but does not confer significant protection against Mycobacterium tuberculosis infection. J. Appl. Microbiol. 2013, 115, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Autumn Smiley, M.; Sanford, D.C.; Triplett, C.A.; Callahan, D.; Frolov, V.; Look, J.; Ruiz, C.; Reece, J.J.; Miles, A.; Ruiz, E.; et al. Comparative immunogenicity and efficacy of thermostable (lyophilized) and liquid formulation of anthrax vaccine candidate AV7909. Vaccine 2019, 37, 6356–6361. [Google Scholar] [CrossRef]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Song, Z.; Yang, J.; He, Q.; Mao, Q.; Bai, Y.; Liu, J.; An, C.; Yan, X.; Cui, B.; et al. Transcriptomic analysis of the innate immune signatures of a SARS-CoV-2 protein subunit vaccine ZF2001 and an mRNA vaccine RRV. Emerg. Microbes Infect. 2022, 11, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Mao, Q.; Peng, X.; He, Z.; Lu, S.; Zhang, J.; Gao, F.; Bian, L.; An, C.; Yu, W.; et al. Immunogenicity and protective efficacy of a recombinant protein subunit vaccine and an inactivated vaccine against SARS-CoV-2 variants in non-human primates. Signal. Transduct. Target. Ther. 2022, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, Q.; An, C.; Mao, Q.; Gao, F.; Bian, L.; Wu, X.; Wang, Q.; Liu, P.; Song, L.; et al. Boosting with heterologous vaccines effectively improves protective immune responses of the inactivated SARS-CoV-2 vaccine. Emerg. Microbes Infect. 2021, 10, 1598–1608. [Google Scholar] [CrossRef]
- Bian, L.; Gao, F.; Zhang, J.; He, Q.; Mao, Q.; Xu, M.; Liang, Z. Effects of SARS-CoV-2 variants on vaccine efficacy and response strategies. Expert Rev. Vaccines 2021, 20, 365–373. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, Q.; Liu, M.; Bian, L.; Liu, J.; Gao, F.; Mao, Q.; Wang, Z.; Wu, X.; Xu, M.; et al. The next major emergent infectious disease: Reflections on vaccine emergency development strategies. Expert Rev. Vaccines 2022, 21, 471–481. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, J.; Xu, W.; Deng, W.; Wang, Y.; Wang, M.; Wang, Q.; Hsieh, M.; Dong, J.; Wang, X.; et al. A novel STING agonist-adjuvanted pan-sarbecovirus vaccine elicits potent and durable neutralizing antibody and T cell responses in mice, rabbits and NHPs. Cell Res. 2022, 32, 269–287. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Sadahira, T.; Ueki, H.; Kinoshita, R.; Murata, H.; Yamamoto, K.I.; Futami, J.; Nasu, Y.; Ochiai, K.; Kumon, H.; et al. Robust cancer-specific gene expression by a novel cassette with hTERT and CMV promoter elements. Oncol. Rep. 2017, 38, 1108–1114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.N.; Li, X.F.; Deng, Y.Q.; Zhao, H.; Huang, Y.J.; Yang, G.; Huang, W.J.; Gao, P.; Zhou, C.; Zhang, R.R.; et al. A Thermostable mRNA Vaccine against COVID-19. Cell 2020, 182, 1271–1283.e1216. [Google Scholar] [CrossRef]
- Tai, W.; Zhang, X.; Drelich, A.; Shi, J.; Hsu, J.C.; Luchsinger, L.; Hillyer, C.D.; Tseng, C.K.; Jiang, S.; Du, L. A novel receptor-binding domain (RBD)-based mRNA vaccine against SARS-CoV-2. Cell Res. 2020, 30, 932–935. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E., Jr.; Lisa, A.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; et al. An infectious SARS-CoV-2 B.1.1.529 Omicron virus escapes neutralization by several therapeutic monoclonal antibodies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Bian, L.; Bai, Y.; Gao, F.; Liu, M.; He, Q.; Wu, X.; Mao, Q.; Xu, M.; Liang, Z. Effective protection of ZF2001 against the SARS-CoV-2 Delta variant in lethal K18-hACE2 mice. Virol. J. 2022, 19, 1–5. [Google Scholar] [CrossRef]
- Sabbaghi, A.; Malek, M.; Abdolahi, S.; Miri, S.M.; Alizadeh, L.; Samadi, M.; Mohebbi, S.R.; Ghaemi, A. A formulated poly (I:C)/CCL21 as an effective mucosal adjuvant for gamma-irradiated influenza vaccine. Virol. J. 2021, 18, 201. [Google Scholar] [CrossRef]
- Nanishi, E.; Borriello, F.; O’Meara, T.R.; McGrath, M.E.; Saito, Y.; Haupt, R.E.; Seo, H.S.; van Haren, S.D.; Cavazzoni, C.B.; Brook, B.; et al. An aluminum hydroxide:CpG adjuvant enhances protection elicited by a SARS-CoV-2 receptor-binding domain vaccine in aged mice. Sci. Transl. Med. 2021, 14, eabj5305. [Google Scholar] [CrossRef]
- Zhao, X.; Li, D.; Ruan, W.; Chen, Z.; Zhang, R.; Zheng, A.; Qiao, S.; Zheng, X.; Zhao, Y.; Dai, L.; et al. Effects of a Prolonged Booster Interval on Neutralization of Omicron Variant. N. Engl. J. Med. 2022, 386, 894–896. [Google Scholar] [CrossRef]
- Song, S.; Zhou, B.; Cheng, L.; Liu, W.; Fan, Q.; Ge, X.; Peng, H.; Fu, Y.X.; Ju, B.; Zhang, Z. Sequential immunization with SARS-CoV-2 RBD vaccine induces potent and broad neutralization against variants in mice. Virol J. 2022, 19, 2. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e114. [Google Scholar] [CrossRef]
- Kingstad-Bakke, B.; Lee, W.; Chandrasekar, S.S.; Gasper, D.J.; Salas-Quinchucua, C.; Cleven, T.; Sullivan, J.A.; Talaat, A.; Osorio, J.E.; Suresh, M. Vaccine-induced systemic and mucosal T cell immunity to SARS-CoV-2 viral variants. Proc. Natl. Acad. Sci. USA 2022, 119, e2118312119. [Google Scholar] [CrossRef]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1-2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Sequence (5′-3′) |
|---|---|---|
| RIG-I | Forward Reverse | GATTATATCCGGAAGACCCTG ATACTGCACCTCTTCCTCC |
| TLR-3 | Forward Reverse | TCATCCAACAGAATCATGAGAC CTAACAGTGCACTTGGTGG |
| TLR-7 | Forward Reverse | CAAGAAAGTTGATGCTATTGGG GTGTCCACATTGGAAACAC |
| TLR-8 | Forward Reverse | AGAAACATGGTTCTCTTGACAC TAGTCTCCTTTCCCAGGCT |
| MyD88 | Forward Reverse | CAGCATTGAGGAGGATTGC GGGACACTGCTGTCTACAG |
| MAVS | Forward Reverse | CAGCAGAAATGAGGAGACC CTATTCTCAGAGCTGCTGTC |
| GAPDH | Forward Reverse | GGAGCGAGATCCCTCCAAAAT GGCTGTTGTCATACTTCTCATGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; An, C.; Bai, Y.; Li, K.; Liu, J.; Wang, Q.; He, Q.; Song, Z.; Zhang, J.; Song, L.; et al. A Novel Single-Stranded RNA-Based Adjuvant Improves the Immunogenicity of the SARS-CoV-2 Recombinant Protein Vaccine. Viruses 2022, 14, 1854. https://doi.org/10.3390/v14091854
Liu D, An C, Bai Y, Li K, Liu J, Wang Q, He Q, Song Z, Zhang J, Song L, et al. A Novel Single-Stranded RNA-Based Adjuvant Improves the Immunogenicity of the SARS-CoV-2 Recombinant Protein Vaccine. Viruses. 2022; 14(9):1854. https://doi.org/10.3390/v14091854
Chicago/Turabian StyleLiu, Dong, Chaoqiang An, Yu Bai, Kelei Li, Jianyang Liu, Qian Wang, Qian He, Ziyang Song, Jialu Zhang, Lifang Song, and et al. 2022. "A Novel Single-Stranded RNA-Based Adjuvant Improves the Immunogenicity of the SARS-CoV-2 Recombinant Protein Vaccine" Viruses 14, no. 9: 1854. https://doi.org/10.3390/v14091854