
Halorubrum pleomorphic virus-6 Membrane Fusion Is Triggered by an S-Layer Component of Its Haloarchaeal Host




Abstract
:1. Introduction
2. Materials and Methods
2.1. Haloarchaeal Host Strains and Viruses and Growth Conditions
2.2. HRPV-6 Propagation and Purification
2.3. S-Layer Extraction
2.4. SDS-PAGE
2.5. LC-MS/MS
2.6. Liposome Production
2.7. R18-Labeling of HRPV-6 and Liposomes
2.8. Liposome Flotation Assay
2.9. Transmission Electron Microscopy (TEM)
2.10. Virus-Cell and Virus-Liposome Fusion Assay (Lipid Mixing Assay)
2.11. Statistical Analysis
3. Results
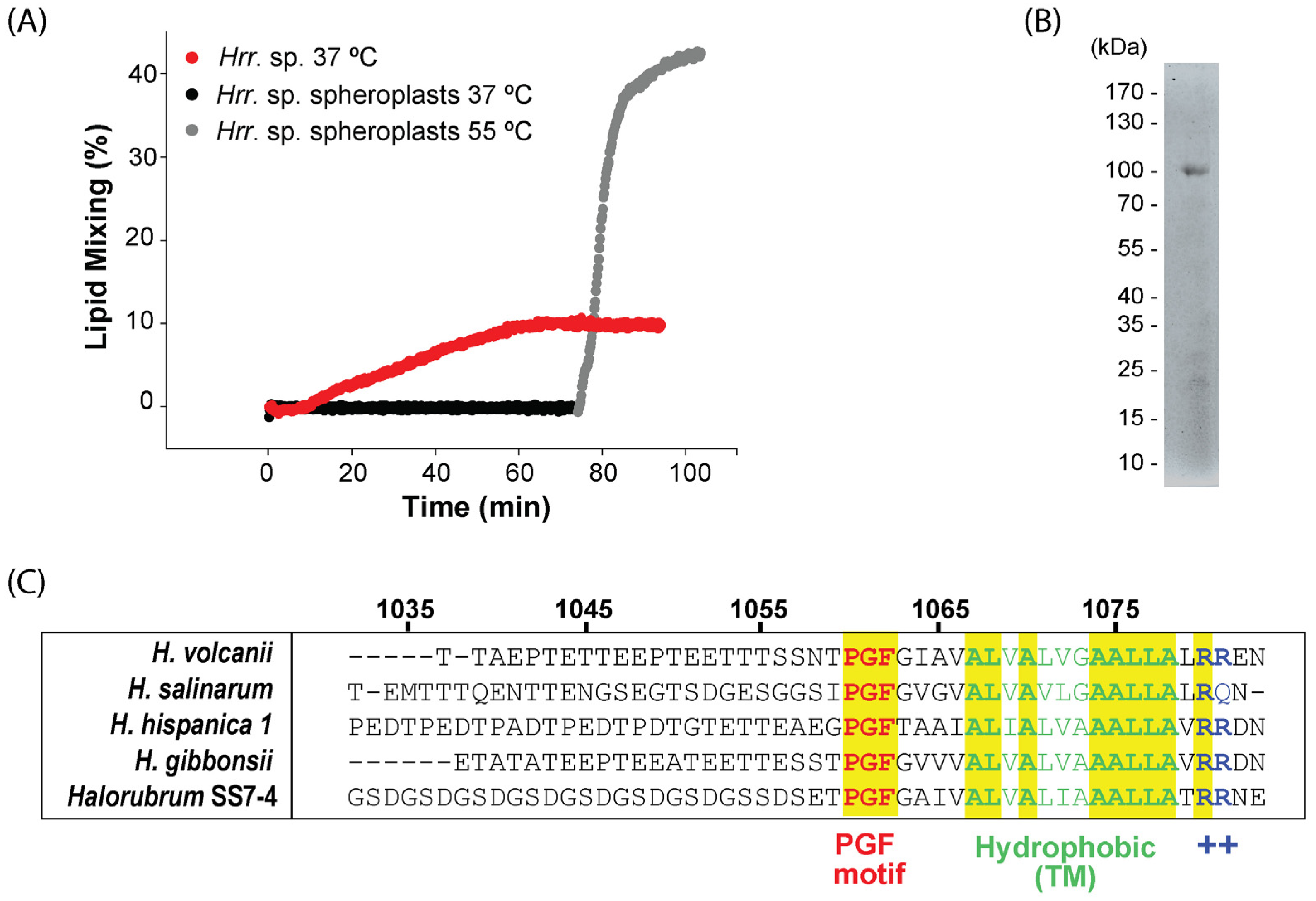
3.1. The Extraction of the Host Cell S-Layer Abrogates HRPV-6 Membrane Fusion
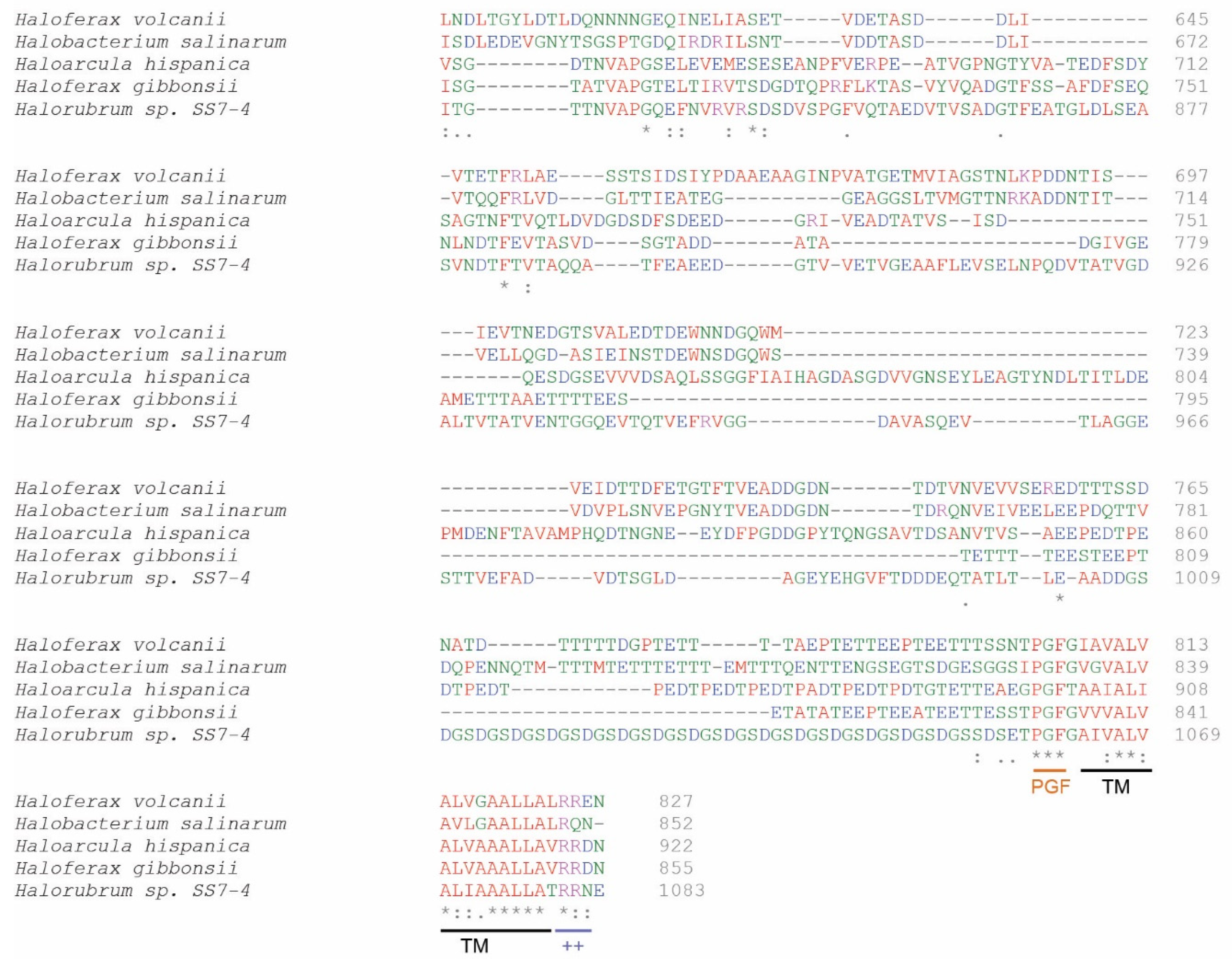
3.2. Characterization of the Host Cell S-Layer Extract
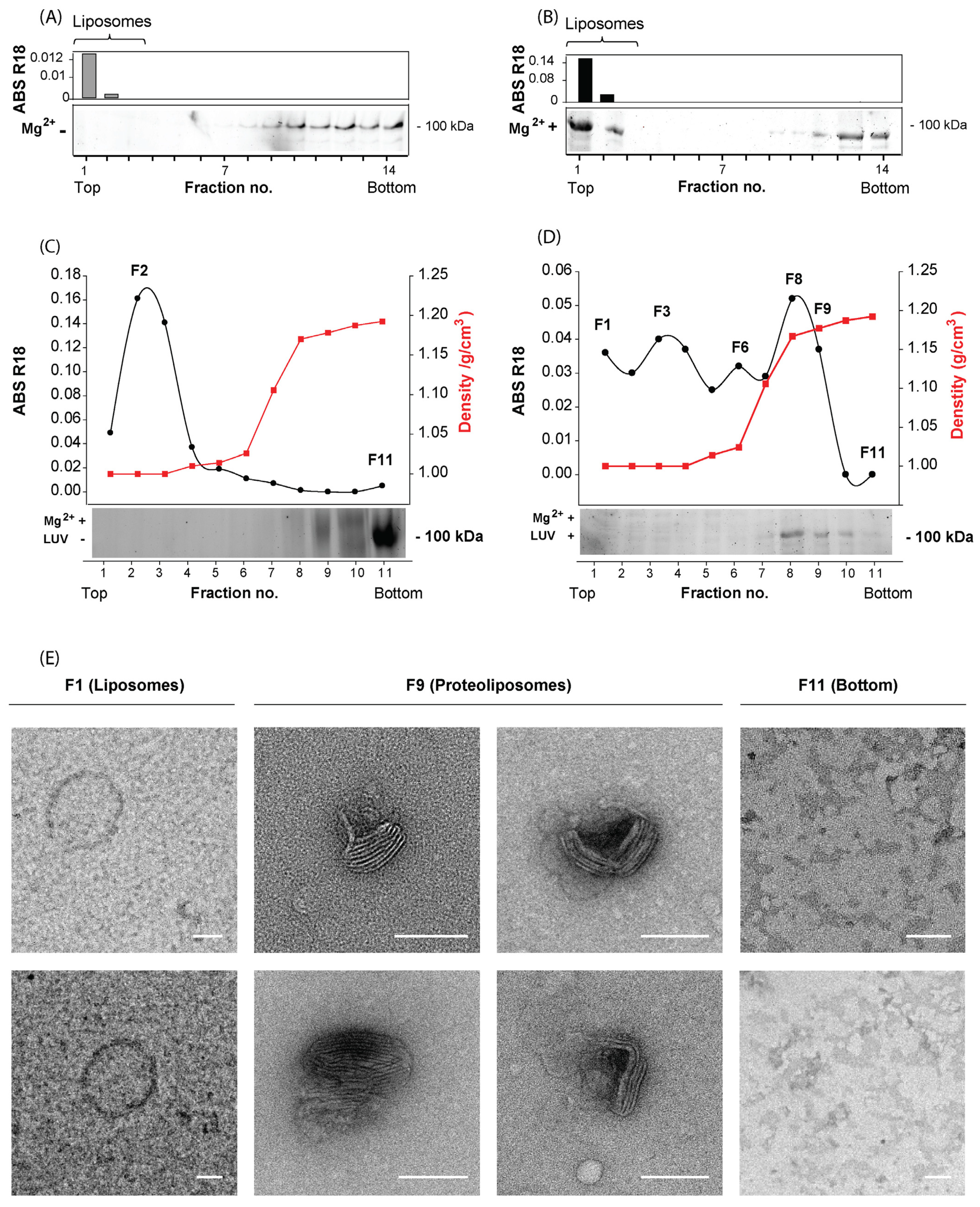
3.3. Characterization of the Host Cell S-Layer Reconstituted onto Proteoliposomes
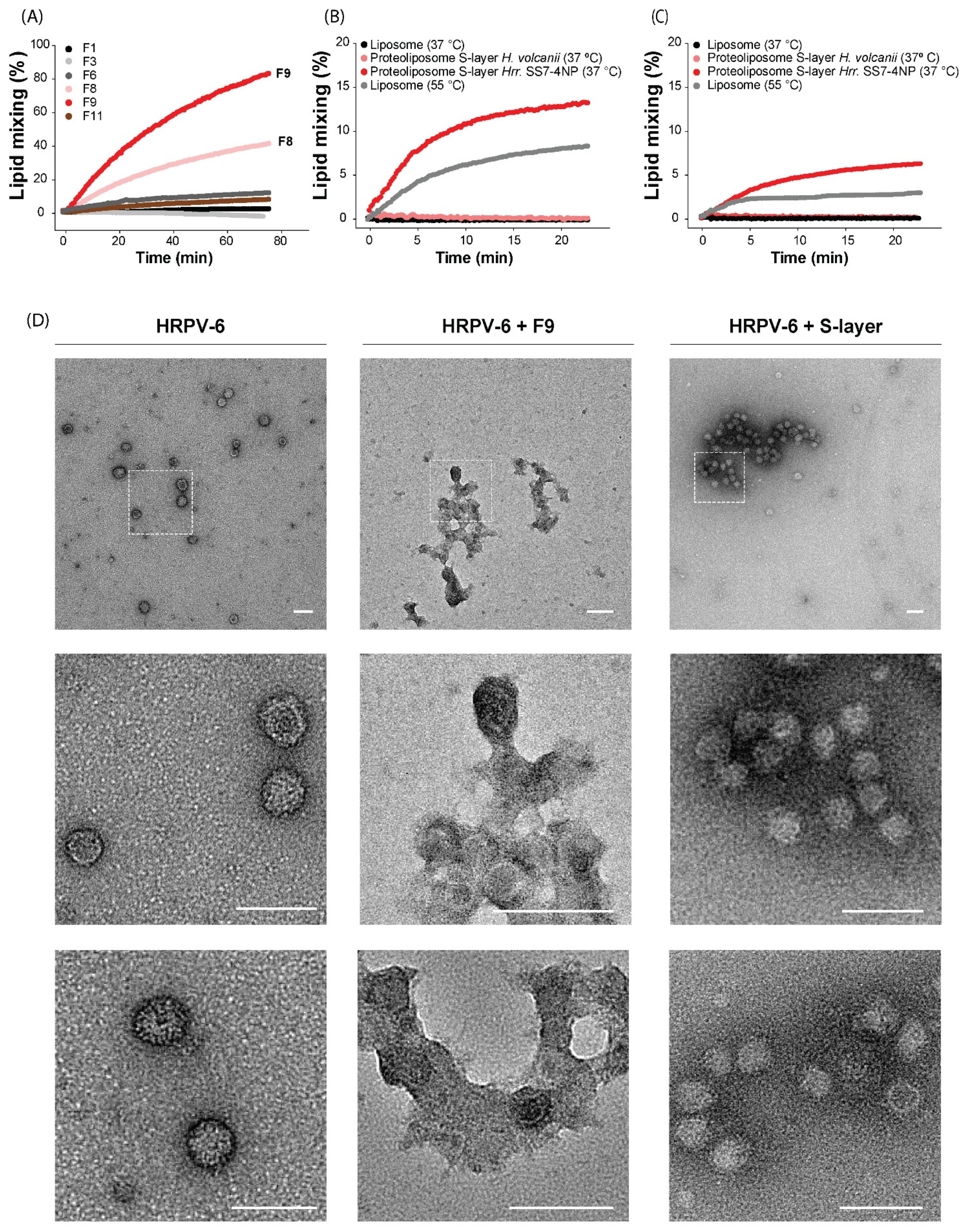
3.4. The S-Layer Extract of Hrr. sp. SS7-4NP Triggers HRPV-6 Membrane Fusion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Pietilä, M.K.; Demina, T.; Atanasova, N.; Oksanen, H.; Bamford, D.H. Archaeal viruses and bacteriophages: Comparisons and contrasts. Trends Microbiol. 2014, 22, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Cvirkaite-Krupovic, V.; Iranzo, J.; Prangishvili, D.; Koonin, E.V. Viruses of archaea: Structural, functional, environmental and evolutionary genomics. Virus Res. 2017, 244, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Prangishvili, D. The Wonderful World of Archaeal Viruses. Annu. Rev. Microbiol. 2013, 67, 565–585. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Roine, E.; Sencilo, A.; Bamford, D.; Oksanen, H.M. Pleolipoviridae, a newly proposed family comprising archaeal pleomorphic viruses with single-stranded or double-stranded DNA genomes. Arch. Virol. 2015, 161, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietila, M.K.; Atanasova, N.S.; Manole, V.; Liljeroos, L.; Butcher, S.J.; Oksanen, H.M.; Bamford, D.H. Virion Architecture Unifies Globally Distributed Pleolipoviruses Infecting Halophilic Archaea. J. Virol. 2012, 86, 5067–5079. [Google Scholar] [CrossRef] [Green Version]
- Luk, A.W.S.; Williams, T.J.; Erdmann, S.; Papke, R.T.; Cavicchioli, R. Viruses of Haloarchaea. Life 2014, 4, 681–715. [Google Scholar] [CrossRef] [Green Version]
- Bamford, D.H.; Pietilä, M.K.; Roine, E.; Atanasova, N.; Dienstbier, A.; Oksanen, H.M. ICTV Report Consortium ICTV Virus Taxonomy Profile: Pleolipoviridae. J. Gen. Virol. 2017, 98, 2916–2917. [Google Scholar] [CrossRef]
- Oren, A. Diversity of halophilic microorganisms: Environments, phylogeny, physiology, and applications. J. Ind. Microbiol. Biotechnol. 2002, 28, 56–63. [Google Scholar] [CrossRef]
- Tittes, C.; Schwarzer, S.; Quax, T.E.F. Viral Hijack of Filamentous Surface Structures in Archaea and Bacteria. Viruses 2021, 13, 164. [Google Scholar] [CrossRef]
- Quemin, E.; Quax, T.E.F. Archaeal viruses at the cell envelope: Entry and egress. Front. Microbiol. 2015, 6, 552. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, S.; Scheele, U.; Garrett, R.A. AAA ATPase p529 of Acidianus two-tailed virus ATV and host receptor recognition. Virology 2011, 421, 61–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.; Baranyi, U.; Rössler, N.; Greineder, B.; Scholz, H.; Witte, A. Natrialba magadii virus φCh1: First complete nucleotide sequence and functional organization of a virus infecting a haloalkaliphilic archaeon. Mol. Microbiol. 2002, 45, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.; Rössler, N.; Iro, M.; Scholz, H.; Witte, A. Haloarchaeal myovirus φCh1 harbours a phase variation system for the production of protein variants with distinct cell surface adhesion specificities. Mol. Microbiol. 2012, 83, 137–150. [Google Scholar] [CrossRef]
- Quemin, E.R.J.; Lucas, S.; Daum, B.; Quax, T.; Kühlbrandt, W.; Forterre, P.; Albers, S.-V.; Prangishvili, D.; Krupovic, M. First Insights into the Entry Process of Hyperthermophilic Archaeal Viruses. J. Virol. 2013, 87, 13379–13385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sleytr, U. Self-assembly of the hexagonally and tetragonally arranged subunits of bacterial surface layers and their reattachment to cell walls. J. Ultrastruct. Res. 1976, 55, 360–377. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.-M.; Pum, D. S-layers: Principles and applications. FEMS Microbiol. Rev. 2014, 38, 823–864. [Google Scholar] [CrossRef]
- Rodrigues, T.; Belmok, A.; Vasconcellos, D.; Schuster, B.; Kyaw, C.M. Archaeal S-Layers: Overview and Current State of the Art. Front. Microbiol. 2017, 8, 2597. [Google Scholar] [CrossRef] [Green Version]
- Albers, S.-V.; Meyer, B.H. The archaeal cell envelope. Nat. Rev. Microbiol. 2011, 9, 414–426. [Google Scholar] [CrossRef]
- Sára, M.; Sleytr, U.B. S-Layer Proteins. J. Bacteriol. 2000, 182, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Bharat, T.A.; von Kügelgen, A.; Alva, V. Molecular Logic of Prokaryotic Surface Layer Structures. Trends Microbiol. 2020, 29, 405–415. [Google Scholar] [CrossRef]
- von Kügelgen, A.; Alva, V.; Bharat, T.A. Complete atomic structure of a native archaeal cell surface. Cell Rep. 2021, 37. [Google Scholar] [CrossRef] [PubMed]
- Kandiba, L.; Guan, Z.; Eichler, J. Lipid modification gives rise to two distinct Haloferax volcanii S-layer glycoprotein populations. Biochim. Biophys. Acta (BBA)-Biomembr. 2012, 1828, 938–943. [Google Scholar] [CrossRef] [Green Version]
- Haft, D.H.; Payne, S.; Selengut, J.D. Archaeosortases and Exosortases Are Widely Distributed Systems Linking Membrane Transit with Posttranslational Modification. J. Bacteriol. 2011, 194, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halim, M.F.A.; Pfeiffer, F.; Zou, J.; Frisch, A.; Haft, D.; Wu, S.; Tolić, N.; Brewer, H.; Payne, S.; Paša-Tolić, L.; et al. Haloferax volcaniiarchaeosortase is required for motility, mating, and C-terminal processing of the S-layer glycoprotein. Mol. Microbiol. 2013, 88, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Halim, M.F.A.; Karch, K.R.; Zhou, Y.; Haft, D.H.; Garcia, B.A.; Pohlschroder, M. Permuting the PGF Signature Motif Blocks both Archaeosortase-Dependent C-Terminal Cleavage and Prenyl Lipid Attachment for the Haloferax volcanii S-Layer Glycoprotein. J. Bacteriol. 2016, 198, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poranen, M.M.; Daugelavičius, R.; Bamford, D.H. Common Principles in Viral Entry. Annu. Rev. Microbiol. 2002, 56, 521–538. [Google Scholar] [CrossRef]
- Zink, I.A.; Pfeifer, K.; Wimmer, E.; Sleytr, U.B.; Schuster, B.; Schleper, C. CRISPR-mediated gene silencing reveals involvement of the archaeal S-layer in cell division and virus infection. Nat. Commun. 2019, 10, 4797. [Google Scholar] [CrossRef] [Green Version]
- Demina, T.A.; Oksanen, H.M. Pleomorphic archaeal viruses: The family Pleolipoviridae is expanding by seven new species. Arch. Virol. 2020, 165, 2723–2731. [Google Scholar] [CrossRef]
- Senčilo, A.; Paulin, L.; Kellner, S.; Helm, M.; Roine, E. Related haloarchaeal pleomorphic viruses contain different genome types. Nucleic Acids Res. 2012, 40, 5523–5534. [Google Scholar] [CrossRef]
- Pietilä, M.K.; Roine, E.; Paulin, L.; Kalkkinen, N.; Bamford, D.H. An ssDNA virus infecting archaea: A new lineage of viruses with a membrane envelope. Mol. Microbiol. 2009, 72, 307–319. [Google Scholar] [CrossRef]
- Pietilä, M.K.; Laurinavičius, S.; Sund, J.; Roine, E.; Bamford, D.H. The Single-Stranded DNA Genome of Novel Archaeal Virus Halorubrum pleomorphic virus 1 Is Enclosed in the Envelope Decorated with Glycoprotein Spikes. J. Virol. 2010, 84, 788–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandiba, L.; Aitio, O.; Helin, J.; Guan, Z.; Permi, P.; Bamford, D.H.; Eichler, J.; Roine, E. Diversity in prokaryotic glycosylation: An archaeal-derived N-linked glycan contains legionaminic acid. Mol. Microbiol. 2012, 84, 578–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Omari, K.; Li, S.; Kotecha, A.; Walter, T.S.; Bignon, E.A.; Harlos, K.; Somerharju, P.; De Haas, F.; Clare, D.K.; Molin, M.; et al. The structure of a prokaryotic viral envelope protein expands the landscape of membrane fusion proteins. Nat. Commun. 2019, 10, 846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielian, M. Mechanisms of Virus Membrane Fusion Proteins. Annu. Rev. Virol. 2014, 1, 171–189. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479–480, 498–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and Mechanisms of Viral Membrane Fusion Proteins: Multiple Variations on a Common Theme. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 189–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyall-Smith, M. The Halohandbook- Protocols for Haloarchaeal Genetics. 2009. Available online: https://haloarchaea.com/wp-content/uploads/2018/10/Halohandbook_2009_v7.3mds.pdf (accessed on 30 October 2017).
- Stroud, A.; Liddell, S.; Allers, T. Genetic and Biochemical Identification of a Novel Single-Stranded DNA-Binding Complex in Haloferax volcanii. Front. Microbiol. 2012, 3, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allers, T.; Ngo, H.-P.; Mevarech, M.; Lloyd, R.G. Development of Additional Selectable Markers for the Halophilic Archaeon Haloferax volcanii Based on the leuB and trpA Genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Sumper, M.; Berg, E.; Mengele, R.; Strobel, I. Primary structure and glycosylation of the S-layer protein of Haloferax volcanii. J. Bacteriol. 1990, 172, 7111–7118. [Google Scholar] [CrossRef] [Green Version]
- Posch, A.; Kohn, J.; Oh, K.; Hammond, M.; Liu, N. V3 Stain-free Workflow for a Practical, Convenient, and Reliable Total Protein Loading Control in Western Blotting. J. Vis. Exp. 2013, 82, e50948. [Google Scholar] [CrossRef] [Green Version]
- Fullmer, M.S.; Ouellette, M.; Louyakis, A.S.; Papke, R.T.; Gogarten, J.P. The Patchy Distribution of Restriction–Modification System Genes and the Conservation of Orphan Methyltransferases in Halobacteria. Genes 2019, 10, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukashin, A.V. GeneMark.hmm: New solutions for gene finding. Nucleic Acids Res. 1998, 26, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Castile, J.D.; Taylor, K.M. Factors affecting the size distribution of liposomes produced by freeze–thaw extrusion. Int. J. Pharm. 1999, 188, 87–95. [Google Scholar] [CrossRef]
- Hoekstra, D.; De Boer, T.; Klappe, K.; Wilschut, J. Fluorescence method for measuring the kinetics of fusion between biological membranes. Biochemistry 1984, 23, 5675–5681. [Google Scholar] [CrossRef]
- Cline, S.W.; Doolittle, W.F. Transformation of members of the genus Haloarcula with shuttle vectors based on Halobacterium halobium and Haloferax volcanii plasmid replicons. J. Bacteriol. 1992, 174, 1076–1080. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2--A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Shalev, Y.; Soucy, S.M.; Papke, R.T.; Gogarten, J.P.; Eichler, J.; Gophna, U. Comparative Analysis of Surface Layer Glycoproteins and Genes Involved in Protein Glycosylation in the Genus Haloferax. Genes 2018, 9, 172. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Lü, Y.; Ren, J.; Wang, Z.; Wang, Q.; Luo, Y.; Han, J.; Xiang, H.; Du, Y.; Jin, C. Identification of the S-layer glycoproteins and their covalently linked glycans in the halophilic archaeonHaloarcula hispanica. Glycobiology 2015, 25, 1150–1162. [Google Scholar] [CrossRef]
- Halim, M.F.A.; Stoltzfus, J.D.; Schulze, S.; Hippler, M.; Pohlschroder, M. ArtA-Dependent Processing of a Tat Substrate Containing a Conserved Tripartite Structure That Is Not Localized at the C Terminus. J. Bacteriol. 2017, 199, e00802-16. [Google Scholar] [CrossRef] [Green Version]
- Schulze, S.; Pfeiffer, F.; Garcia, B.A.; Pohlschroder, M. Comprehensive glycoproteomics shines new light on the complexity and extent of glycosylation in archaea. PLoS Biol. 2021, 19, e3001277. [Google Scholar] [CrossRef] [PubMed]
- Trent, J.D.; Kagawa, H.K.; Yaoi, T.; Olle, E.; Zaluzec, N.J. Chaperonin filaments: The archaeal cytoskeleton? Proc. Natl. Acad. Sci. USA 1997, 94, 5383–5388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, S.-V.; Koning, S.M.; Konings, W.N.; Driessen, A.J. Insights into ABC Transport in Archaea. J. Bioenerg. Biomembr. 2004, 36, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, G.; Engelhard, M.; Müller, V. Bioenergetics of the Archaea. Microbiol. Mol. Biol. Rev. 1999, 63, 570–620. [Google Scholar] [CrossRef] [Green Version]
- Acuña, R.; Bignon, E.A.; Mancini, R.; Lozach, P.-Y.; Tischler, N.D. Acidification triggers Andes hantavirus membrane fusion and rearrangement of Gc into a stable post-fusion homotrimer. J. Gen. Virol. 2015, 96, 3192–3197. [Google Scholar] [CrossRef]
- Stiasny, K.; Allison, S.L.; Schalich, J.; Heinz, F.X. Membrane Interactions of the Tick-Borne Encephalitis Virus Fusion Protein E at Low pH. J. Virol. 2002, 76, 3784–3790. [Google Scholar] [CrossRef] [Green Version]
- Fischer, H.; Polikarpov, I.; Craievich, A.F. Average protein density is a molecular-weight-dependent function. Protein Sci. 2009, 13, 2825–2828. [Google Scholar] [CrossRef]
- Fernandes, S.; São-José, C. Enzymes and Mechanisms Employed by Tailed Bacteriophages to Breach the Bacterial Cell Barriers. Viruses 2018, 10, 396. [Google Scholar] [CrossRef] [Green Version]
- Mahfoud, R.; Garmy, N.; Maresca, M.; Yahi, N.; Puigserver, A.; Fantini, J. Identification of a Common Sphingolipid-binding Domain in Alzheimer, Prion, and HIV-1 Proteins. J. Biol. Chem. 2002, 277, 11292–11296. [Google Scholar] [CrossRef] [Green Version]
- Mattar, S.; Scharf, B.; Kent, S.B.; Rodewald, K.; Oesterhelt, D.; Engelhard, M. The primary structure of halocyanin, an archaeal blue copper protein, predicts a lipid anchor for membrane fixation. J. Biol. Chem. 1994, 269, 14939–14945. [Google Scholar] [CrossRef]
- Storf, S.; Pfeiffer, F.; Dilks, K.; Chen, Z.Q.; Imam, S.; Pohlschröder, M. Mutational and Bioinformatic Analysis of Haloarchaeal Lipobox-Containing Proteins. Archaea 2010, 2010, 410975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sleytr, U.B.; Huber, C.; Ilk, N.; Pum, D.; Schuster, B.; Egelseer, E.M. S-layers as a tool kit for nanobiotechnological applications. FEMS Microbiol. Lett. 2007, 267, 131–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession/Locus Tag Numbers (NCBI/UniProtKB) | Length (aa) | MW [KDa] | Score | Sequence Coverage (%) | Unique Peptides | Cell Localization |
|---|---|---|---|---|---|---|---|
| PGF-CTERM sorting domain containing protein | TKX57172/ EXE44_11260 | 1083 | 111.4 | 88.7 | 12.7 | 10 | Periplasm (this paper) |
| Thermosome subunit protein | TKX57972/ EXE44_08630 | 550 | 57.7 | 47.9 | 23.4 | 10 | Membrane/ cytoplasm [52,53] |
| Carbohydrate ABC transporter substrate-binding protein | TKX56931/ EXE44_12795 | 432 | 46.0 | 43.5 | 15.7 | 4 | Periplasm [54] |
| Class II fumarate hydratase | TKX59009/ EXE44_05545 | 469 | 49.9 | 39.8 | 24.1 | 8 | Periplasm [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bignon, E.A.; Chou, K.R.; Roine, E.; Tischler, N.D. Halorubrum pleomorphic virus-6 Membrane Fusion Is Triggered by an S-Layer Component of Its Haloarchaeal Host. Viruses 2022, 14, 254. https://doi.org/10.3390/v14020254
Bignon EA, Chou KR, Roine E, Tischler ND. Halorubrum pleomorphic virus-6 Membrane Fusion Is Triggered by an S-Layer Component of Its Haloarchaeal Host. Viruses. 2022; 14(2):254. https://doi.org/10.3390/v14020254
Chicago/Turabian StyleBignon, Eduardo A., Kevin R. Chou, Elina Roine, and Nicole D. Tischler. 2022. "Halorubrum pleomorphic virus-6 Membrane Fusion Is Triggered by an S-Layer Component of Its Haloarchaeal Host" Viruses 14, no. 2: 254. https://doi.org/10.3390/v14020254