
Nanomechanical and Vibrational Signature of Chikungunya Viral Particles

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Culture and Inactivation
2.2. Atomic Force Microscopy (AFM)
2.3. Raman Spectroscopy (RS)
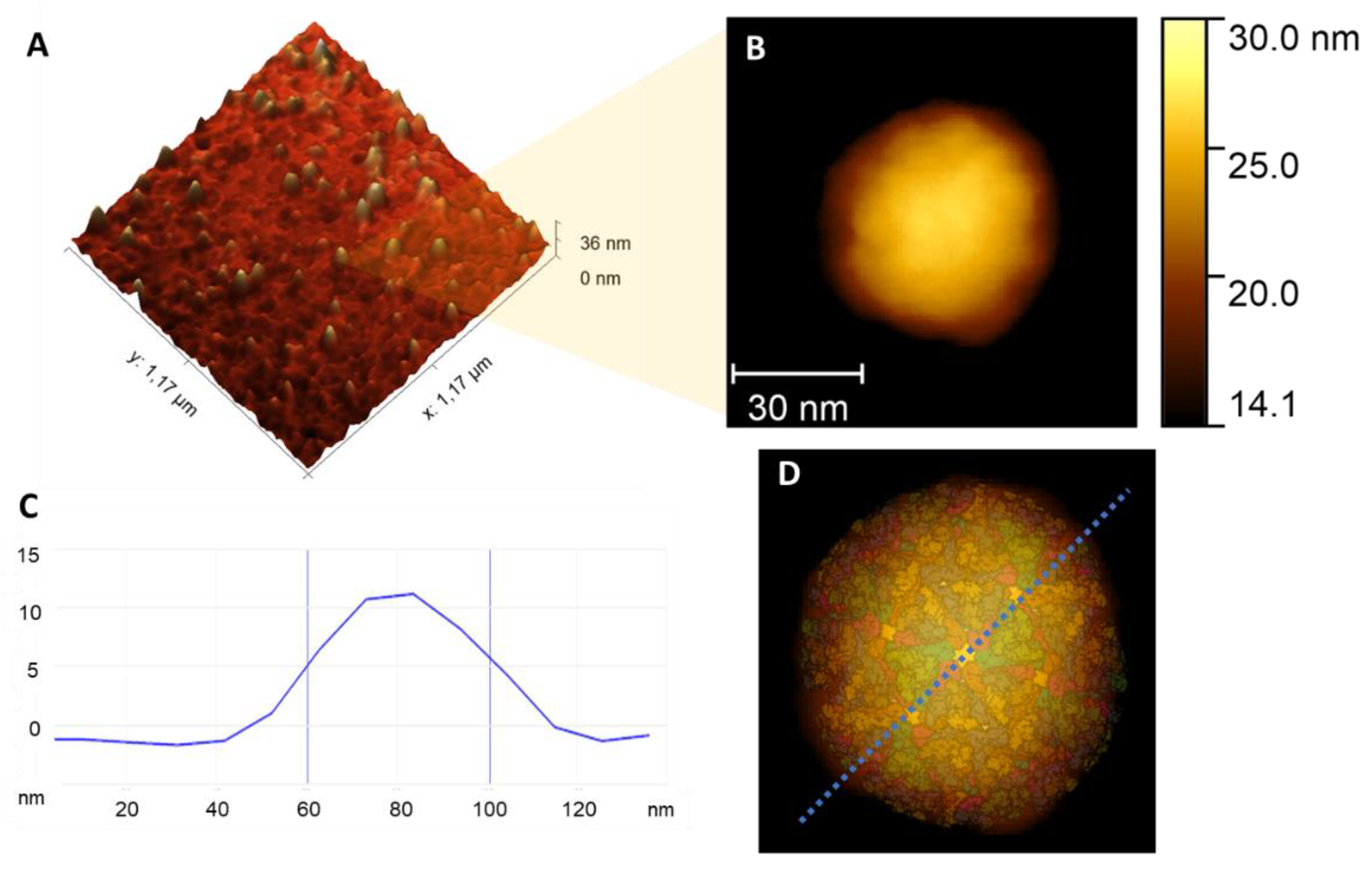
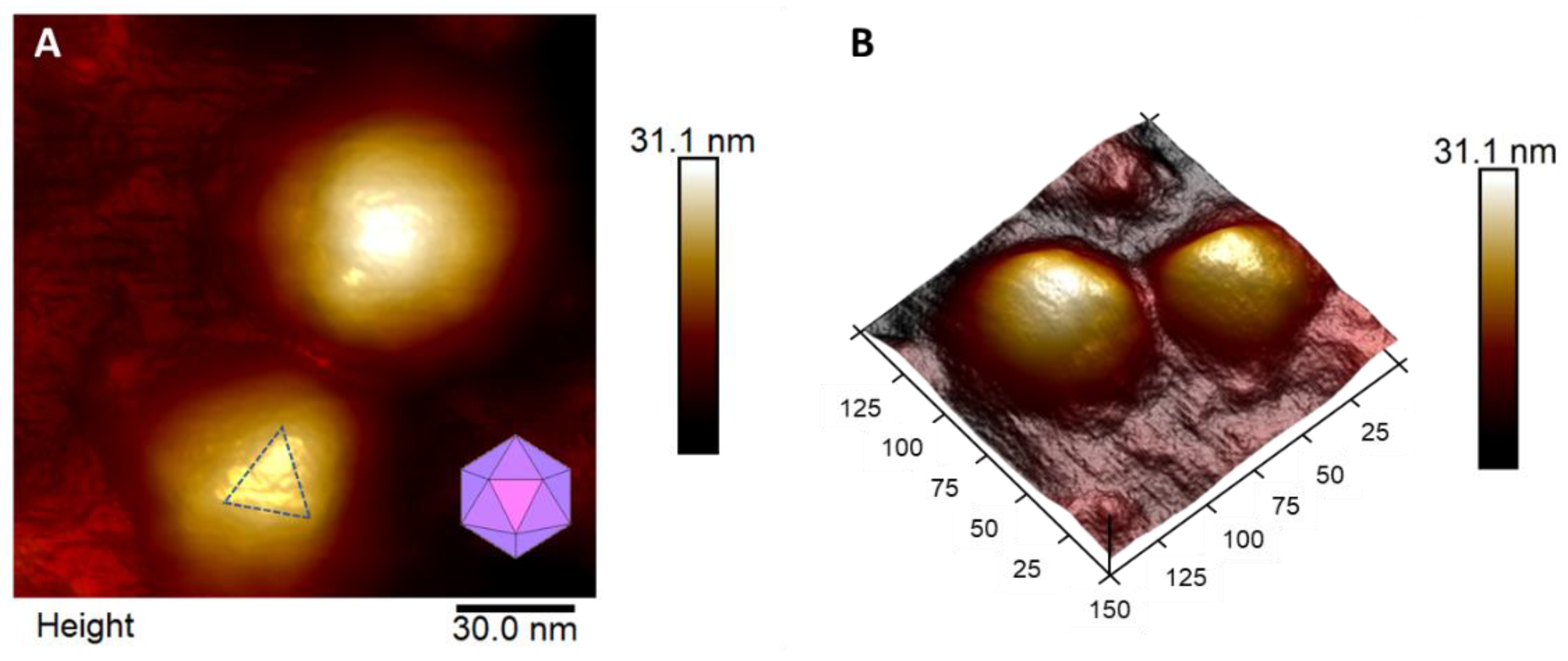
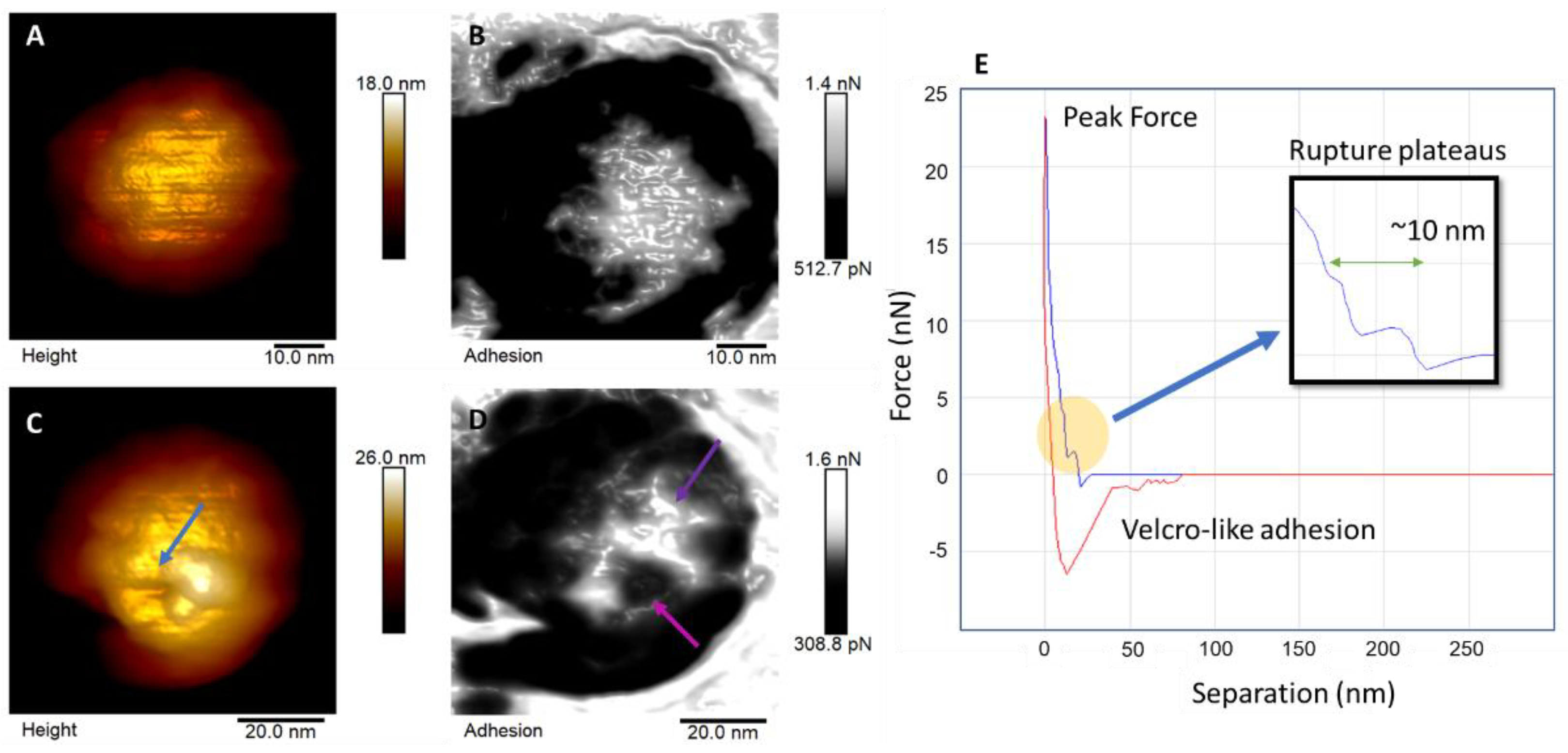
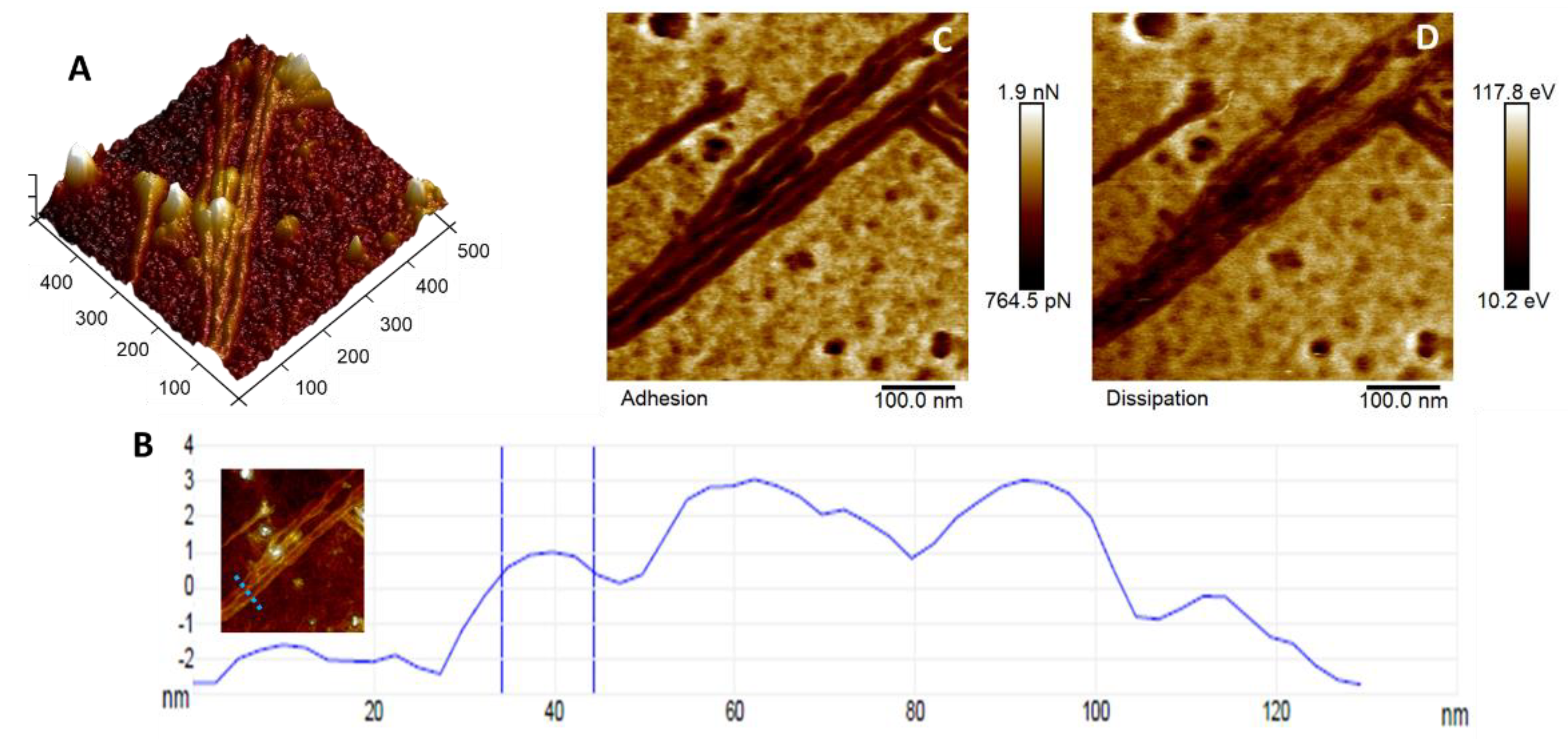
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.-N.; Deng, C.-L.; Li, J.-Q.; Li, N.; Zhang, Q.-Y.; Ye, H.-Q.; Yuan, Z.-M.; Zhang, B. Infectious Chikungunya Virus (CHIKV) with a Complete Capsid Deletion: A New Approach for a CHIKV Vaccine. J. Virol. 2019, 93, e00504-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kril, V.; Aïqui-Reboul-Paviet, O.; Briant, L.; Amara, A. New Insights into Chikungunya Virus Infection and Pathogenesis. Annu. Rev. Virol. 2021, 8, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, K.N.; Javed, F.; Ejaz, M.; Ali, M.; Mujaddadi, N.; Khan, A.A.; Khattak, A.A.; Zaib, A.; Ahmad, I.; Saeed, W.K.; et al. The Global Emergence of Chikungunya Infection: An Integrated View. Rev. Med. Virol. 2022, 32, e2287. [Google Scholar] [CrossRef] [PubMed]
- Basore, K.; Kim, A.S.; Nelson, C.A.; Zhang, R.; Smith, B.K.; Uranga, C.; Vang, L.; Cheng, M.; Gross, M.L.; Smith, J.; et al. Cryo-EM Structure of Chikungunya Virus in Complex with the Mxra8 Receptor. Cell 2019, 177, 1725–1737.e16. [Google Scholar] [CrossRef]
- Galán-Huerta, K.A.; Rivas-Estilla, A.M.; Fernández-Salas, I.; Farfan-Ale, J.A.; Ramos-Jiménez, J. Chikungunya Virus: A General Overview. Med. Univ. 2015, 17, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Tsetsarkin, K.A.; Chen, R.; Sherman, M.B.; Weaver, S.C. Chikungunya Virus: Evolution and Genetic Determinants of Emergence. Curr. Opin. Virol. 2011, 1, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Her, Z.; Kam, Y.-W.; Lin, R.T.P.; Ng, L.F.P. Chikungunya: A Bending Reality. Microbes Infect. 2009, 11, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Natrajan, M.S.; Rojas, A.; Waggoner, J.J. Beyond Fever and Pain: Diagnostic Methods for Chikungunya Virus. J. Clin. Microbiol. 2019, 57, e00350-19. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, R.J.; Dowd, K.A.; Beth Post, C.; Pierson, T.C. Shake, Rattle, and Roll: Impact of the Dynamics of Flavivirus Particles on Their Interactions with the Host. Virology 2015, 479–480, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Lima, R.; Souza, P.F.N.; Guedes, M.I.F.; Santos-Oliveira, R.; Alencar, L.M.R. SARS-CoV-2 Unrevealed: Ultrastructural and Nanomechanical Analysis. Langmuir 2021, 37, 10762–10769. [Google Scholar] [CrossRef]
- de Araujo Dorneles, M.L.; Cardoso-Lima, R.; Souza, P.F.N.; Santoro Rosa, D.; Magne, T.M.; Santos-Oliveira, R.; Alencar, L.M.R. Zika Virus (ZIKV): A New Perspective on the Nanomechanical and Structural Properties. Viruses 2022, 14, 1727. [Google Scholar] [CrossRef] [PubMed]
- Greber, U.F. (Ed.) Physical Virology: Virus Structure and Mechanics; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2019; Volume 1215, ISBN 978-3-030-14740-2. [Google Scholar]
- Roos, W.H.; Bruinsma, R.; Wuite, G.J.L. Physical Virology. Nat. Phys. 2010, 6, 733–743. [Google Scholar] [CrossRef]
- Kol, N.; Gladnikoff, M.; Barlam, D.; Shneck, R.Z.; Rein, A.; Rousso, I. Mechanical Properties of Murine Leukemia Virus Particles: Effect of Maturation. Biophys. J. 2006, 91, 767–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockley, P.G.; Twarock, R. Emerging Topics in Physical Virology; Imperial College Press: London, UK, 2010; ISBN 978-1-84816-464-2. [Google Scholar]
- Pablo, P.J. de The Application of Atomic Force Microscopy for Viruses and Protein Shells: Imaging and Spectroscopy, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 105, ISBN 978-0-12-818456-1. [Google Scholar]
- Asghari-Khiavi, M.; Wood, B.R.; Mechler, A.; Bambery, K.R.; Buckingham, D.W.; Cooke, B.M.; McNaughton, D. Correlation of Atomic Force Microscopy and Raman Micro-Spectroscopy to Study the Effects of Ex Vivo Treatment Procedures on Human Red Blood Cells. Analyst 2010, 135, 525–530. [Google Scholar] [CrossRef]
- Němeček, D.; Thomas, G.J. Chapter 16—Raman Spectroscopy of Viruses and Viral Proteins. In Frontiers of Molecular Spectroscopy; Laane, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 553–595. ISBN 978-0-444-53175-9. [Google Scholar]
- Roos, W.H.; Gertsman, I.; May, E.R.; Brooks, C.L.; Johnson, J.E.; Wuite, G.J.L. Mechanics of Bacteriophage Maturation. Proc. Natl. Acad. Sci. United States Am. 2012, 109, 2342–2347. [Google Scholar] [CrossRef] [Green Version]
- Krieg, M.; Fläschner, G.; Alsteens, D.; Gaub, B.M.; Roos, W.H.; Wuite, G.J.L.; Gaub, H.E.; Gerber, C.; Dufrêne, Y.F.; Müller, D.J. Atomic Force Microscopy-Based Mechanobiology. Nat. Rev. Phys. 2019, 1, 41–57. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-C.; Chiang, Y.-W.; Yang, C.-H.; Liou, J.-W. Atomic Force Microscopy in Biology and Biomedicine. Tzu Chi Med. J. 2012, 24, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.J.H.; Ang, S.K. (Eds.) Chikungunya Virus: Methods and Protocols; Methods in Molecular Biology; Springer: New York, NY, USA, 2016; Volume 1426, ISBN 978-1-4939-3616-8. [Google Scholar]
- Yadav, S.; Sadique, M.A.; Ranjan, P.; Kumar, N.; Singhal, A.; Srivastava, A.K.; Khan, R. SERS Based Lateral Flow Immunoassay for Point-of-Care Detection of SARS-CoV-2 in Clinical Samples. ACS Appl. Biol. Mater. 2021, 4, 2974–2995. [Google Scholar] [CrossRef]
- Han, X.X.; Rodriguez, R.S.; Haynes, C.L.; Ozaki, Y.; Zhao, B. Surface-Enhanced Raman Spectroscopy. Nat. Rev. Methods Primers 2022, 1, 1–17. [Google Scholar] [CrossRef]
- Bank, R.P.D. RCSB PDB—2XFB: Chikungunya E1 E2 Envelope Glycoproteins Fitted in Sindbis Virus Cryo-EM Map. Available online: https://www.rcsb.org/structure/2XFB (accessed on 22 November 2022).
- Metz, S.W.; Pijlman, G.P. Function of Chikungunya Virus Structural Proteins. In Chikungunya Virus; Okeoma, C.M., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 63–74. ISBN 978-3-319-42956-4. [Google Scholar]
- Moreno-Cerrada, D.; Rodríguez, C.; Moreno-Madrid, F.; Selivanovitch, E.; Douglas, T.; Pablo, P.J.; de Silván, M.M. Loading the Dice: The Orientation of Virus-like Particles Adsorbed on Titanate Assisted Organosilanized Surfaces. Biointerphases 2019, 14, 011001. [Google Scholar] [CrossRef]
- Shields, P.A.; Farrah, S.R. Characterization of Virus Adsorption by Using DEAE-Sepharose and Octyl-Sepharose. Appl. Environ. Microbiol. 2002, 68, 3965–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, T.; Fratini, M.; Frey, F.; Yserentant, K.; Liu, Y.; Weber, E.; Galior, K.; Ohmes, J.; Braun, F.; Herten, D.-P.; et al. Forces during Cellular Uptake of Viruses and Nanoparticles at the Ventral Side. Nat. Commun. 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozden, S.; Lucas-Hourani, M.; Ceccaldi, P.-E.; Basak, A.; Valentine, M.; Benjannet, S.; Hamelin, J.; Jacob, Y.; Mamchaoui, K.; Mouly, V.; et al. Inhibition of Chikungunya Virus Infection in Cultured Human Muscle Cells by Furin Inhibitors: Impairment of the maturation of the E2 surface glycoprotein. J. Biol. Chem. 2008, 283, 21899–21908. [Google Scholar] [CrossRef] [Green Version]
- Butt, H.J.; Cappella, B.; Kappl, M. Force Measurements with the Atomic Force Microscope: Technique, Interpretation and Applications. Surf. Sci. Rep. 2005, 59, 1–152. [Google Scholar] [CrossRef] [Green Version]
- Myshkin, N.; Kovalev, A. Adhesion and Surface Forces in Polymer Tribology—A Review. Friction 2018, 6, 143–155. [Google Scholar] [CrossRef]
- Israelachvili, J.N. Intermolecular and Surface Forces, 3rd ed.; Academic Press: Cambridge, MA, USA, 2011; ISBN 978-0-12-391927-4. [Google Scholar]
- Leite, F.L.; Bueno, C.C.; Da Róz, A.L.; Ziemath, E.C.; Oliveira, O.N. Theoretical Models for Surface Forces and Adhesion and Their Measurement Using Atomic Force Microscopy. Int. J. Mol. Sci. 2012, 13, 12773–12856. [Google Scholar] [CrossRef]
- Snijder, J.; Ivanovska, I.L.; Baclayon, M.; Roos, W.H.; Wuite, G.J.L. Probing the Impact of Loading Rate on the Mechanical Properties of Viral Nanoparticles. Micron 2012, 43, 1343–1350. [Google Scholar] [CrossRef]
- Ogharandukun, E.; Tewolde, W.; Damtae, E.; Wang, S.; Ivanov, A.; Kumari, N.; Nekhai, S.; Chandran, P.L. Velcro-like Mannose and Slime-like Sialic Acid Interactions Guide Self-Adhesion and Aggregation of Virus N-Glycan Shields. bioRxiv 2020, 3, 1–31. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, S. Designer Nanomaterials Using Chiral Self-Assembling Peptide Systems and Their Emerging Benefit for Society. Chem. Soc. Rev. 2012, 41, 4736–4754. [Google Scholar] [CrossRef]
- Ko, S.H.; Su, M.; Zhang, C.; Ribbe, A.E.; Jiang, W.; Mao, C. Synergistic Self-Assembly of RNA and DNA Molecules. Nat. Chem. 2010, 2, 1050–1055. [Google Scholar] [CrossRef]
- Jacobson, D.R.; Saleh, O.A. Counting the Ions Surrounding Nucleic Acids. Nucleic Acids Res. 2017, 45, 1596–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rates, E.R.D.; Almeida, C.D.; Costa, E.d.P.F.; Farias, R.J.d.M.; Santos-Oliveira, R.; Alencar, L.M.R. Layer-by-Layer Investigation of Ultrastructures and Biomechanics of Human Cornea. Int. J. Mol. Sci. 2022, 23, 7833. [Google Scholar] [CrossRef]
- Dudha, N.; Rana, J.; Rajasekharan, S.; Gabrani, R.; Gupta, A.; Chaudhary, V.K.; Gupta, S. Host–Pathogen Interactome Analysis of Chikungunya Virus Envelope Proteins E1 and E2. Virus Genes 2015, 50, 200–209. [Google Scholar] [CrossRef]
- Lakshmi, R.J.; Kartha, V.B.; Murali Krishna, C.; R Solomon, J.G.; Ullas, G.; Uma Devi, P. Tissue Raman Spectroscopy for the Study of Radiation Damage: Brain Irradiation of Mice. Radiat. Res. 2002, 157, 175–182. [Google Scholar] [CrossRef]
- Delfino, I.; Ricciardi, V.; Manti, L.; Lasalvia, M.; Lepore, M. Multivariate Analysis of Difference Raman Spectra of the Irradiated Nucleus and Cytoplasm Region of SH-SY5Y Human Neuroblastoma Cells. Sensors 2019, 19, 3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2007, 42, 493–541. [Google Scholar] [CrossRef]
- De Gelder, J.; De Gussem, K.; Vandenabeele, P.; Moens, L. Reference Database of Raman Spectra of Biological Molecules. J. Raman Spectrosc. 2007, 38, 1133–1147. [Google Scholar] [CrossRef]
- Madzharova, F.; Heiner, Z.; Gühlke, M.; Kneipp, J. Surface-Enhanced Hyper-Raman Spectra of Adenine, Guanine, Cytosine, Thymine, and Uracil. J. Phys. Chem. C 2016, 120, 15415–15423. [Google Scholar] [CrossRef] [Green Version]
- Rafiq, S.; Majeed, M.I.; Nawaz, H.; Rashid, N.; Yaqoob, U.; Batool, F.; Bashir, S.; Akbar, S.; Abubakar, M.; Ahmad, S.; et al. Surface-Enhanced Raman Spectroscopy for Analysis of PCR Products of Viral RNA of Hepatitis C Patients. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 259, 119908. [Google Scholar] [CrossRef]
- Czamara, K.; Majzner, K.; Pacia, M.Z.; Kochan, K.; Kaczor, A.; Baranska, M. Raman Spectroscopy of Lipids: A Review. J. Raman Spectrosc. 2015, 46, 4–20. [Google Scholar] [CrossRef]
- Ke, W.; Zhou, D.; Wu, J.; Ji, K. Surface-Enhanced Raman Spectra of Calf Thymus DNA Adsorbed on Concentrated Silver Colloid. Appl. Spectrosc. 2005, 59, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Kurouski, D.; Postiglione, T.; Deckert-Gaudig, T.; Deckert, V.; Lednev, I.K. Amide I Vibrational Mode Suppression in Surface (SERS) and Tip (TERS) Enhanced Raman Spectra of Protein Specimens. Analyst 2013, 138, 1665–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, T.; Li, Z.; Zhang, J.; Evilevitch, A.; Kurouski, D. Nanoscale Structural Characterization of Individual Viral Particles Using Atomic Force Microscopy Infrared Spectroscopy (AFM-IR) and Tip-Enhanced Raman Spectroscopy (TERS). Anal. Chem. 2020, 92, 11297–11304. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavenumber (cm−1) | RNA related | Amino acid/ Protein | Lipid/ Carbohydrate | Reference |
|---|---|---|---|---|
| 793 | O-P-O | [42] | ||
| 812 | O-P-O | [43] | ||
| 851 | Tyrosine | [43,44] | ||
| 837 | Proline | [44] | ||
| 950 | Polysaccharides | [42,44] | ||
| 974 | Ribose | [44] | ||
| 991 | Cytosine | [45] | ||
| 1030 | Adenine | [46] | ||
| 1104 | Uracil | [46] | ||
| 1124 | Lipids | [44] | ||
| 1140 | Adenine | [46] | ||
| 1200 | L-Tyrosine | [45] | ||
| 1210 | Guanine | [46] | ||
| 1230–1260 | Amide III | [43] | ||
| 1288 | Cytosine | [47] | ||
| 1305 | Lipids | [48] | ||
| 1325 | Guanine | [43] | ||
| 1334 | L-Proline | [45] | ||
| 1351 | Guanine | [46] | ||
| 1383 | Guanine | [46] | ||
| 1394 | Uracil | [45] | ||
| 1420–1430 | Adenine, Guanine | [43] | ||
| 1445–1456 | Phospholipids, Lipids | [42,44] | ||
| 1512 | Adenine | [49] | ||
| 1563 | Guanine | [46] | ||
| 1583 | Guanine | [43,47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso-Lima, R.; Filho, J.F.S.D.; de Araujo Dorneles, M.L.; Gaspar, R.S.; Souza, P.F.N.; Costa dos Santos, C.; Santoro Rosa, D.; Santos-Oliveira, R.; Alencar, L.M.R. Nanomechanical and Vibrational Signature of Chikungunya Viral Particles. Viruses 2022, 14, 2821. https://doi.org/10.3390/v14122821
Cardoso-Lima R, Filho JFSD, de Araujo Dorneles ML, Gaspar RS, Souza PFN, Costa dos Santos C, Santoro Rosa D, Santos-Oliveira R, Alencar LMR. Nanomechanical and Vibrational Signature of Chikungunya Viral Particles. Viruses. 2022; 14(12):2821. https://doi.org/10.3390/v14122821
Chicago/Turabian StyleCardoso-Lima, Ruana, Joel Félix Silva Diniz Filho, Maria Luiza de Araujo Dorneles, Renato Simões Gaspar, Pedro Filho Noronha Souza, Clenilton Costa dos Santos, Daniela Santoro Rosa, Ralph Santos-Oliveira, and Luciana Magalhães Rebelo Alencar. 2022. "Nanomechanical and Vibrational Signature of Chikungunya Viral Particles" Viruses 14, no. 12: 2821. https://doi.org/10.3390/v14122821