
Norovirus: An Overview of Virology and Preventative Measures


Abstract
:1. Introduction
2. Norovirus (NoV) Virology
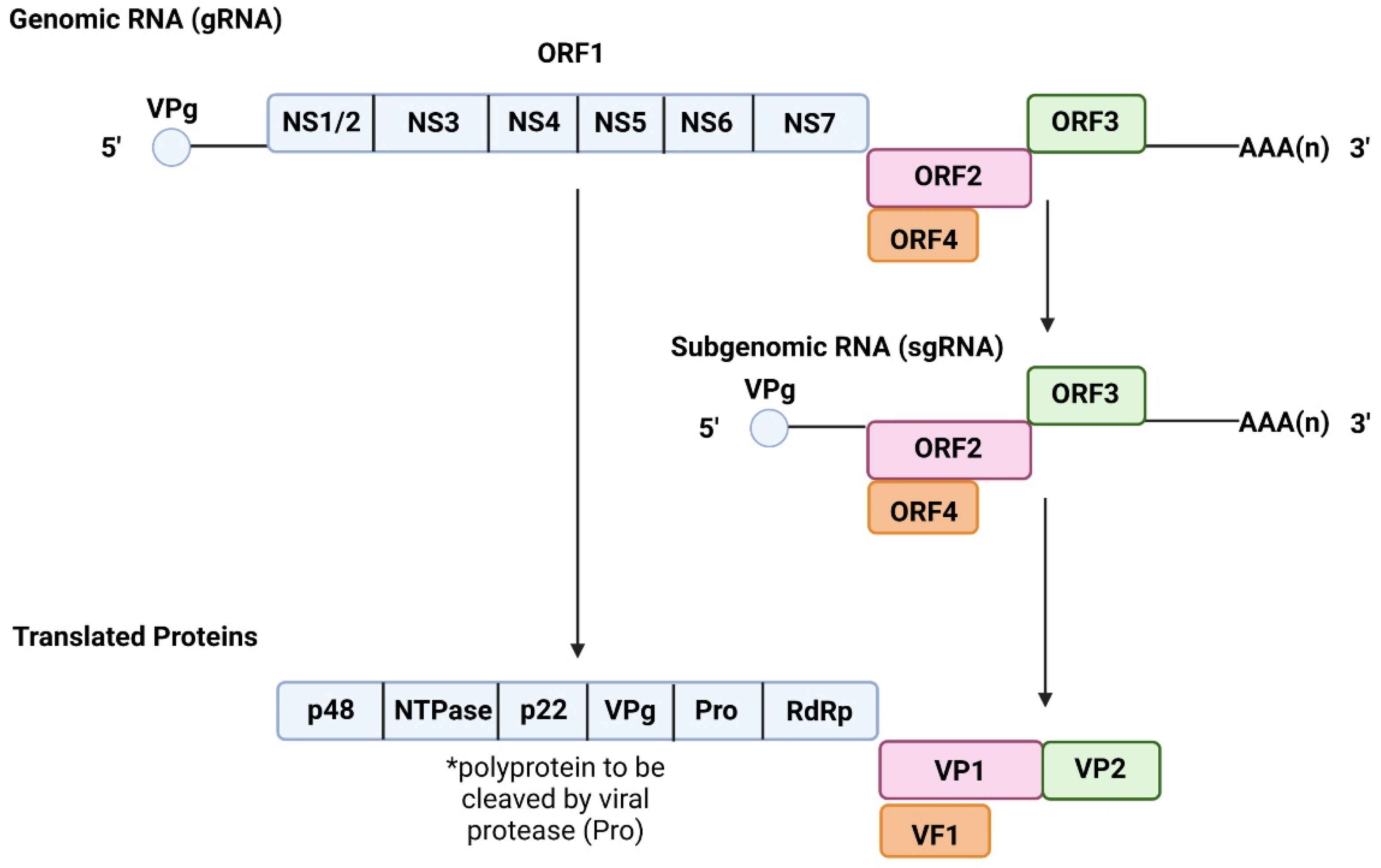
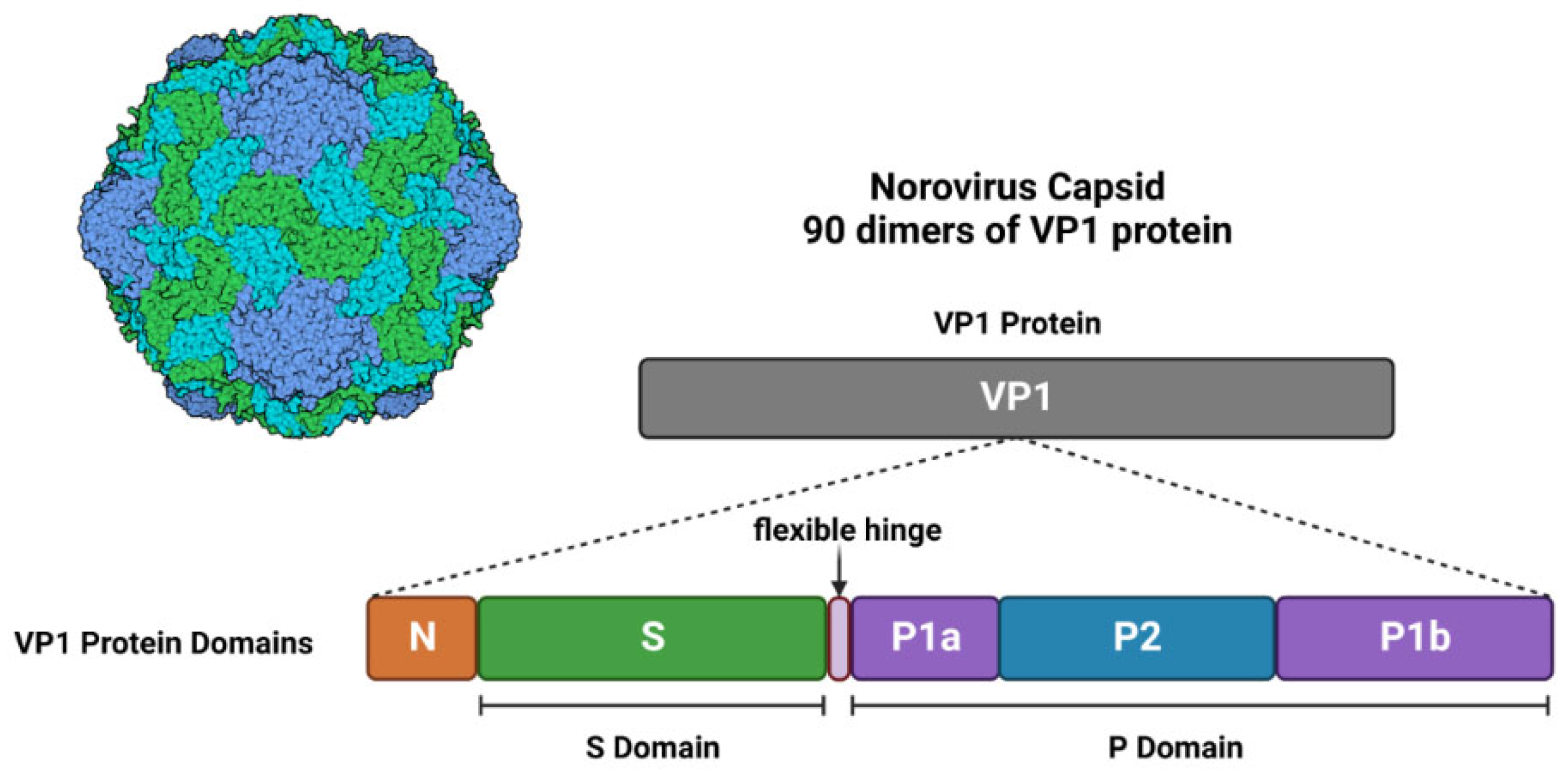
2.1. Genome and Structure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | Gene | Protein | Function | References |
|---|---|---|---|---|
| ORF1 | NS1/2 | p48 N-term Protein | Regulates RdRp activity | [12,15,22] |
| NS3 | Nucleotide Triphosphatase (NTPase) | Assists in RNA synthesis | [12,15,23] | |
| NS4 | p22 | Antagonistic for Golgi apparatus functions | [12,15,24] | |
| NS5 | Viral VPg Protein | Directs host machinery to favour viral protein synthesis | [12,15,25] | |
| ORF2 ORF3 | NS6 | Viral Protease | Processes viral polyprotein | [12,15,25] |
| NS7 | RNA-dependent RNA Polymerase | Translates viral RNA | [12,15,25,26] | |
| VP1 | Major Viral Protein 1 | Binding interface for HBGA | [12,15,26,27] | |
| VP2 | Minor Viral Protein 2 | Supports viral capsid stability | [12,15,21,26] | |
| ORF4 | VF1 | Viral Factor 1 Protein | Antagonizes innate immune responses | [14,21] |
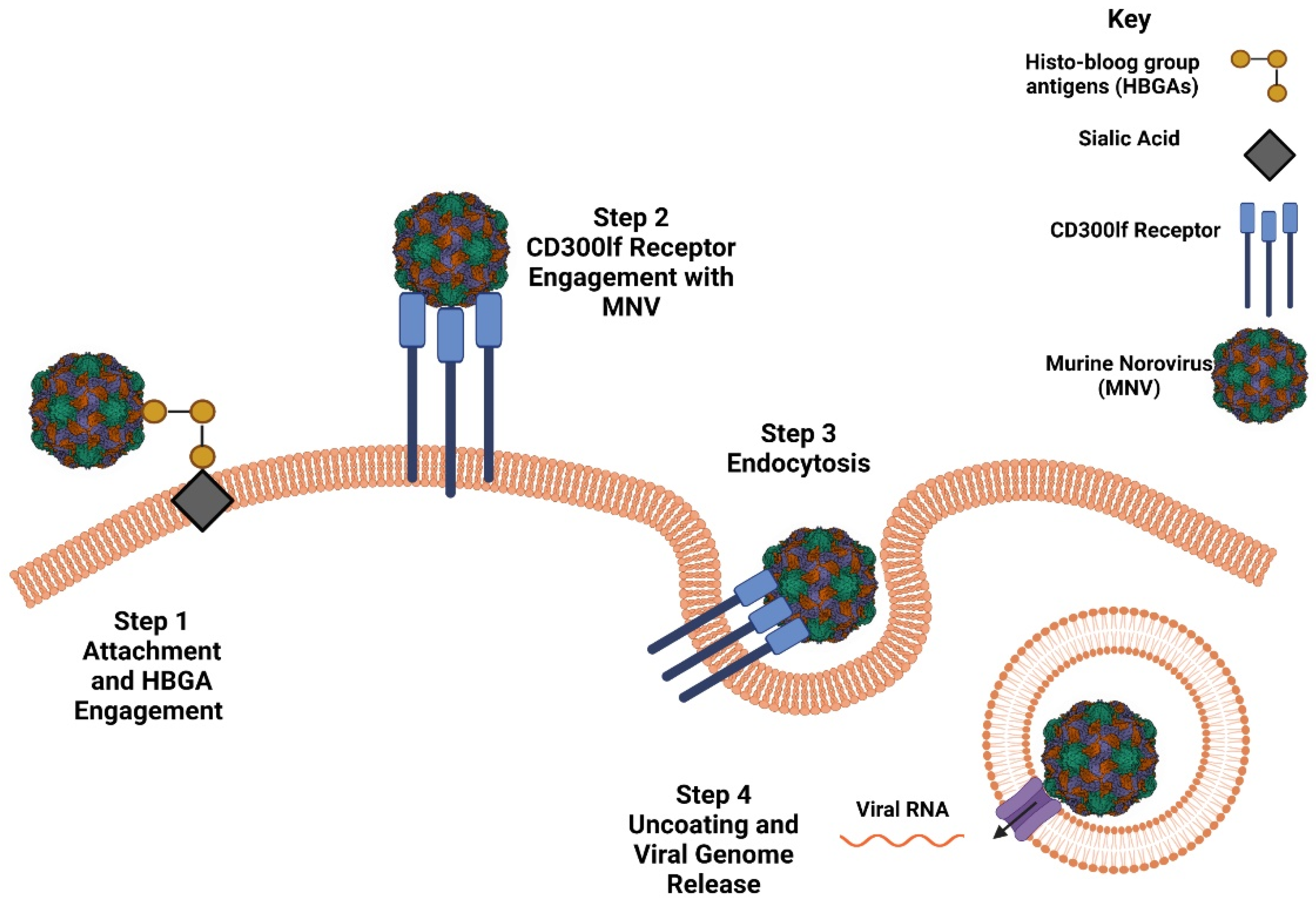
2.2. Host Susceptibility and Receptor Engagement
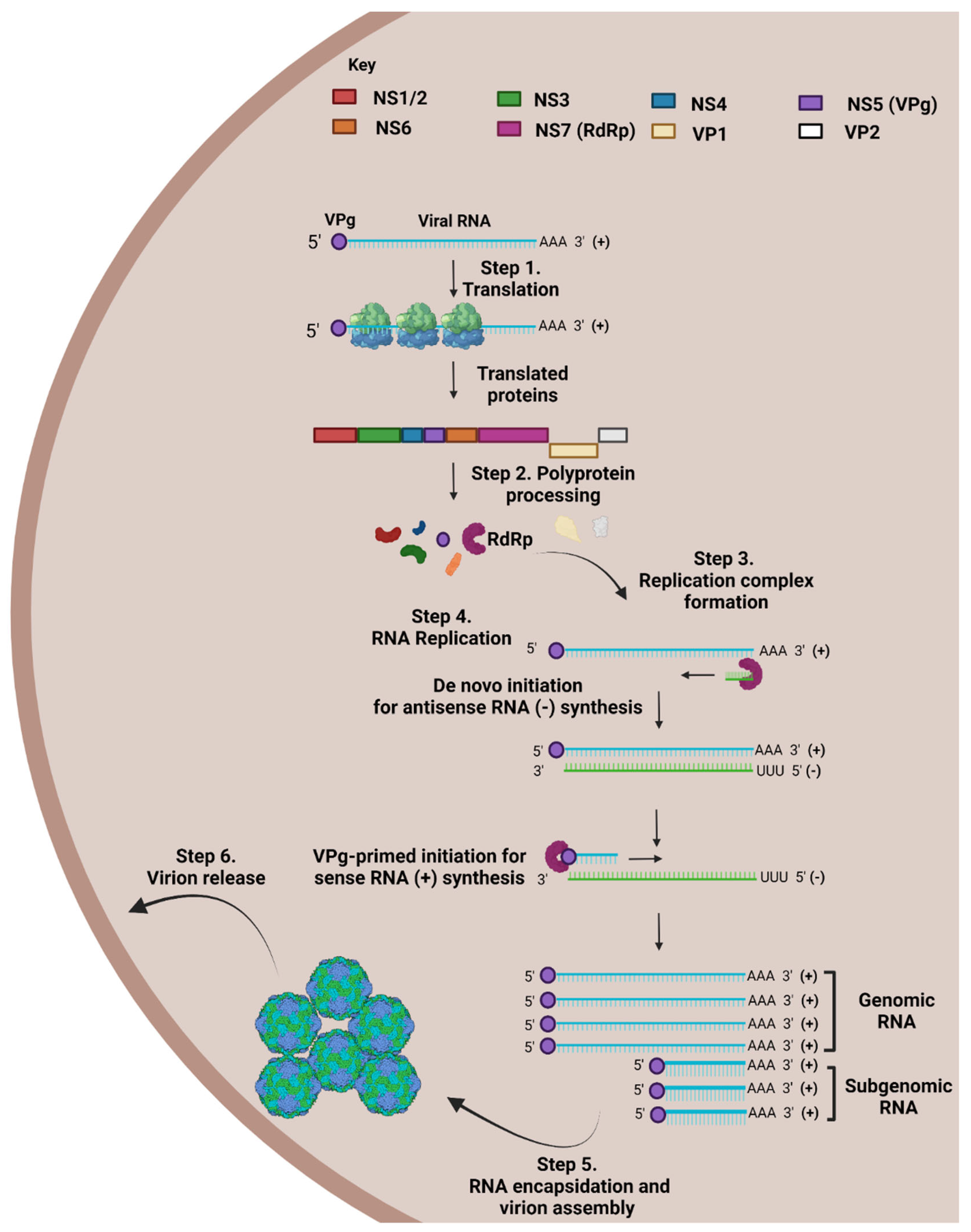
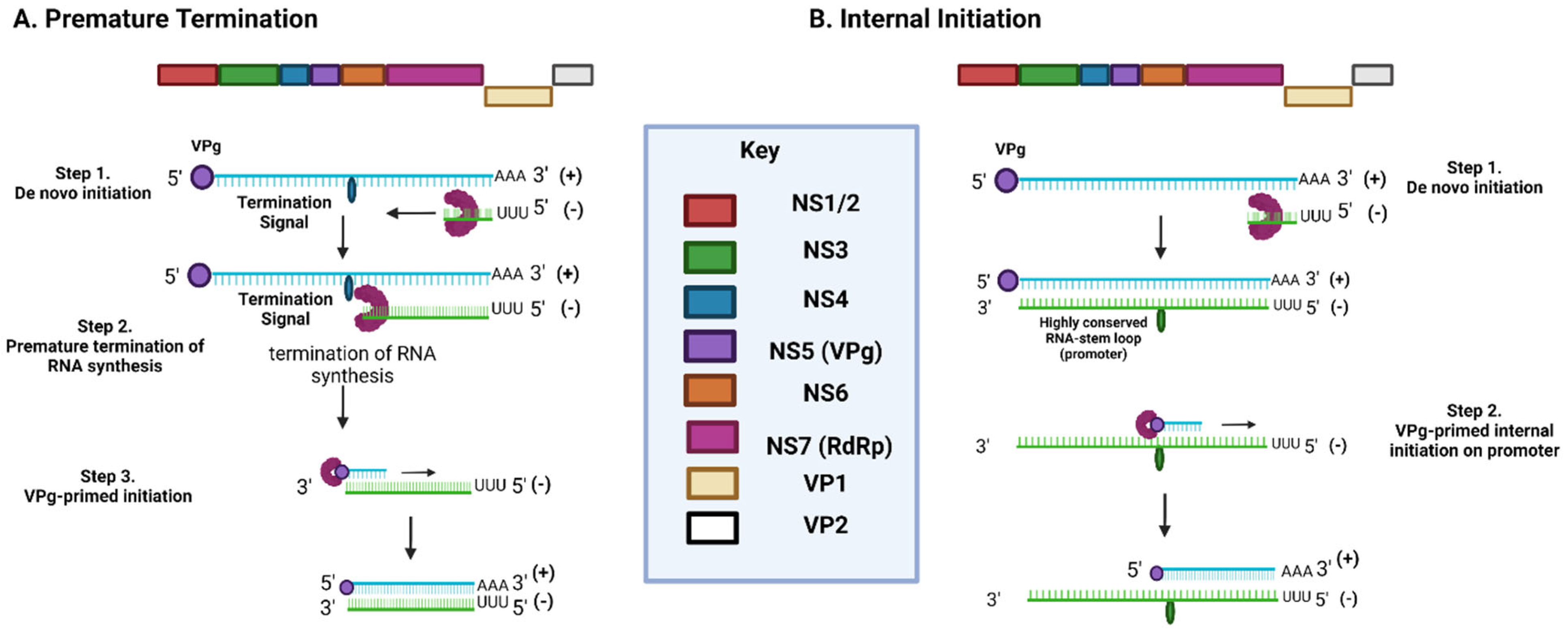
2.3. NoV Life Cycle
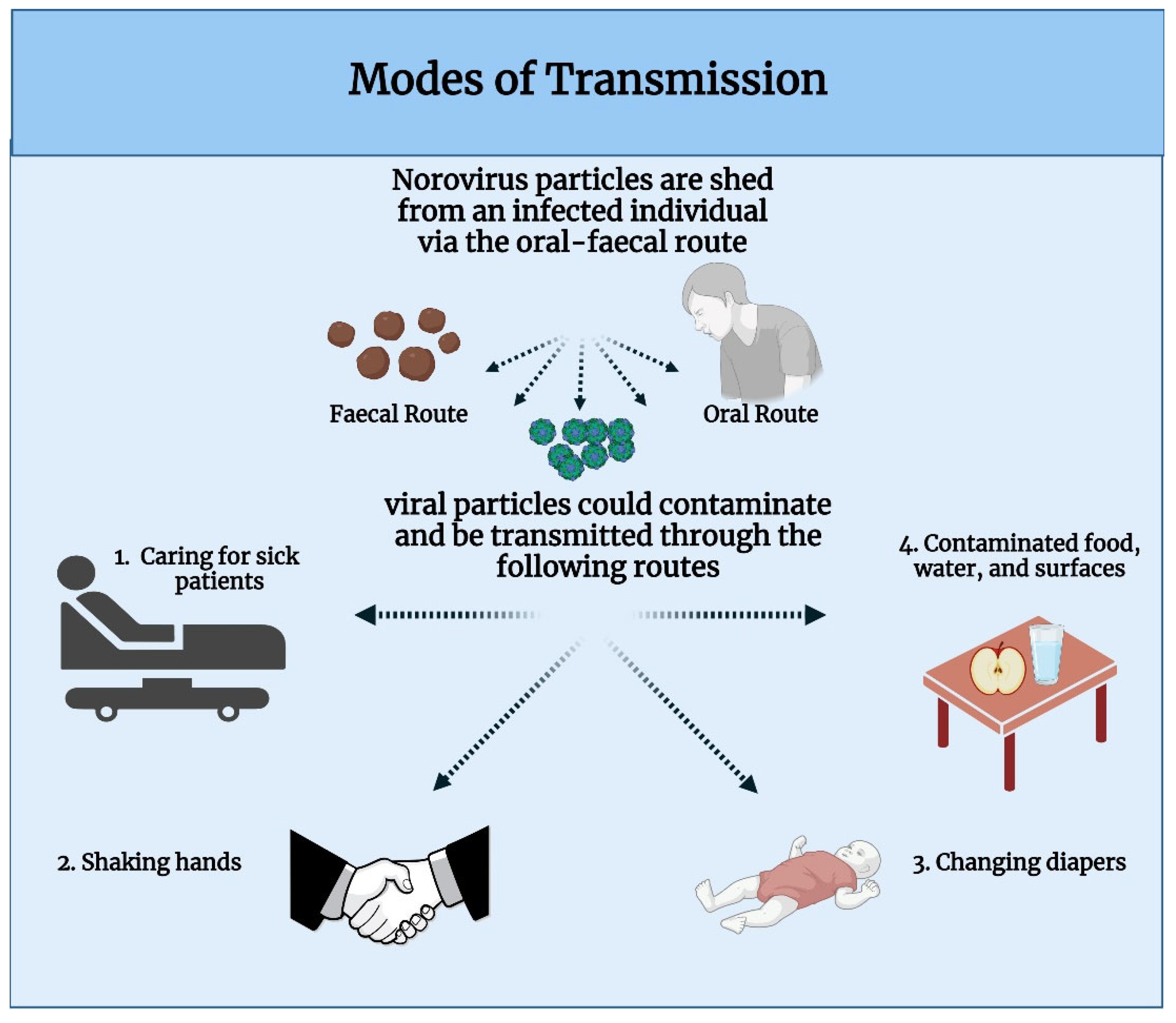
2.4. Modes of Transmission
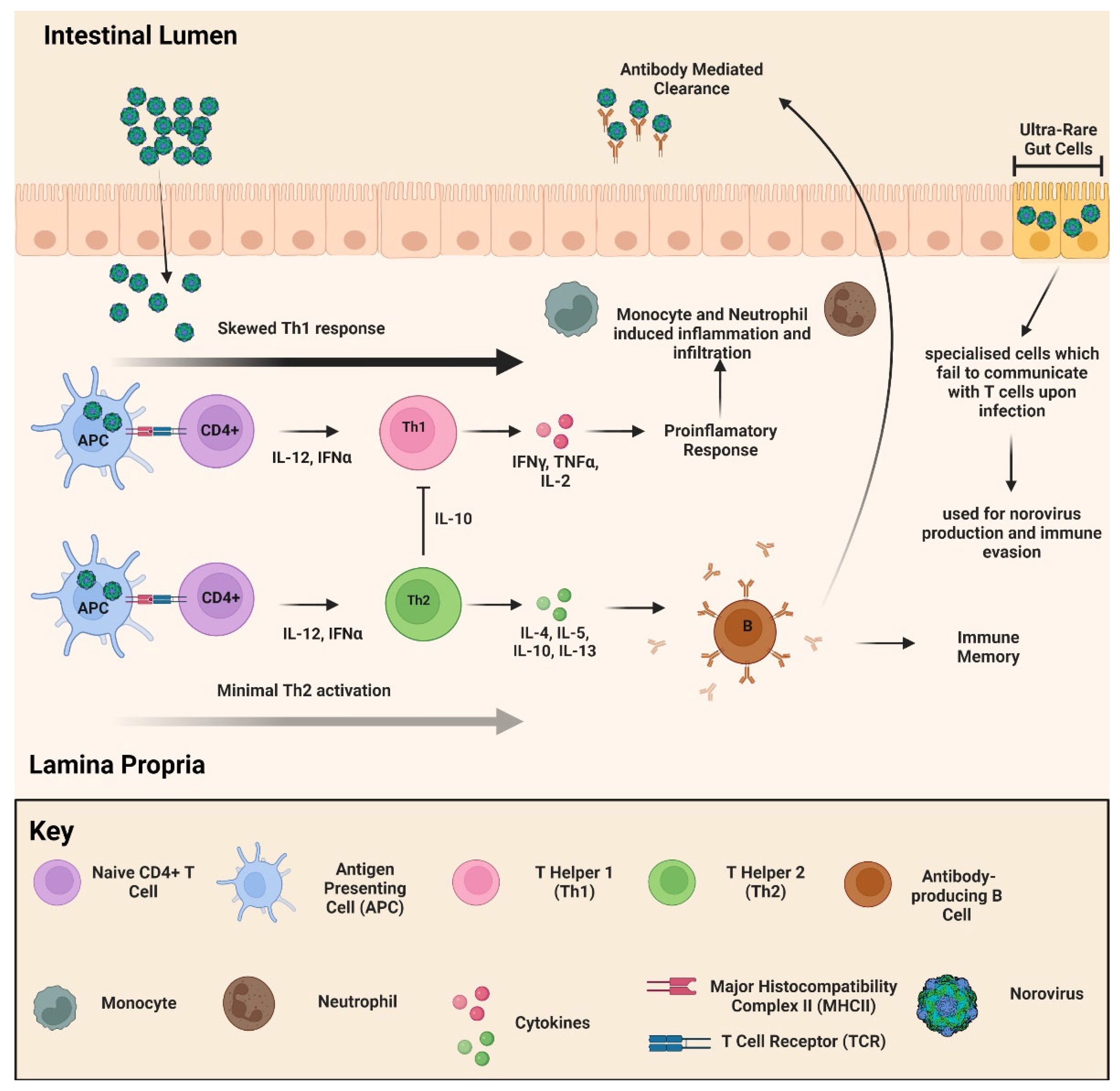
2.5. Pathophysiology and Immune Response
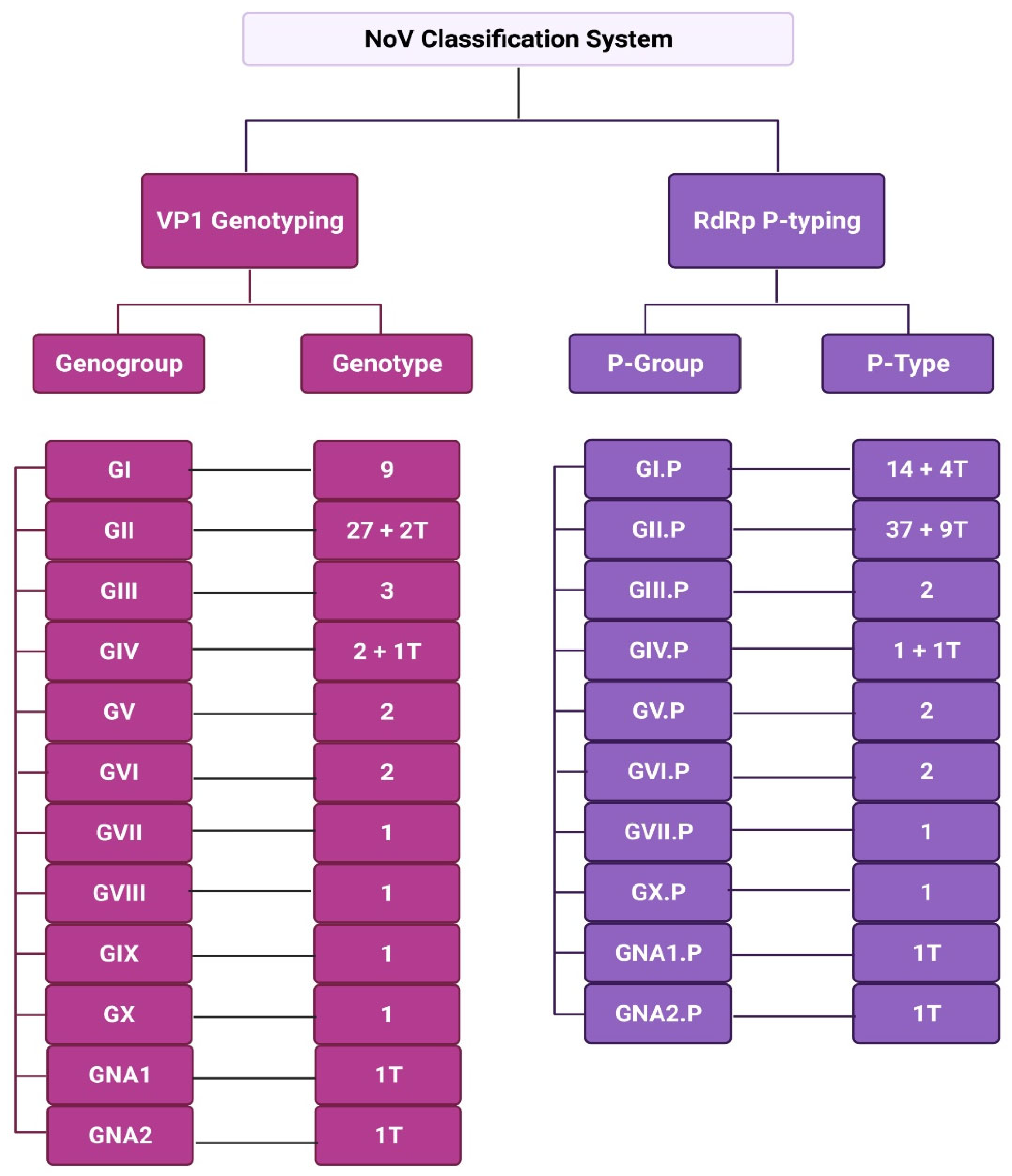
3. Classification of NoV
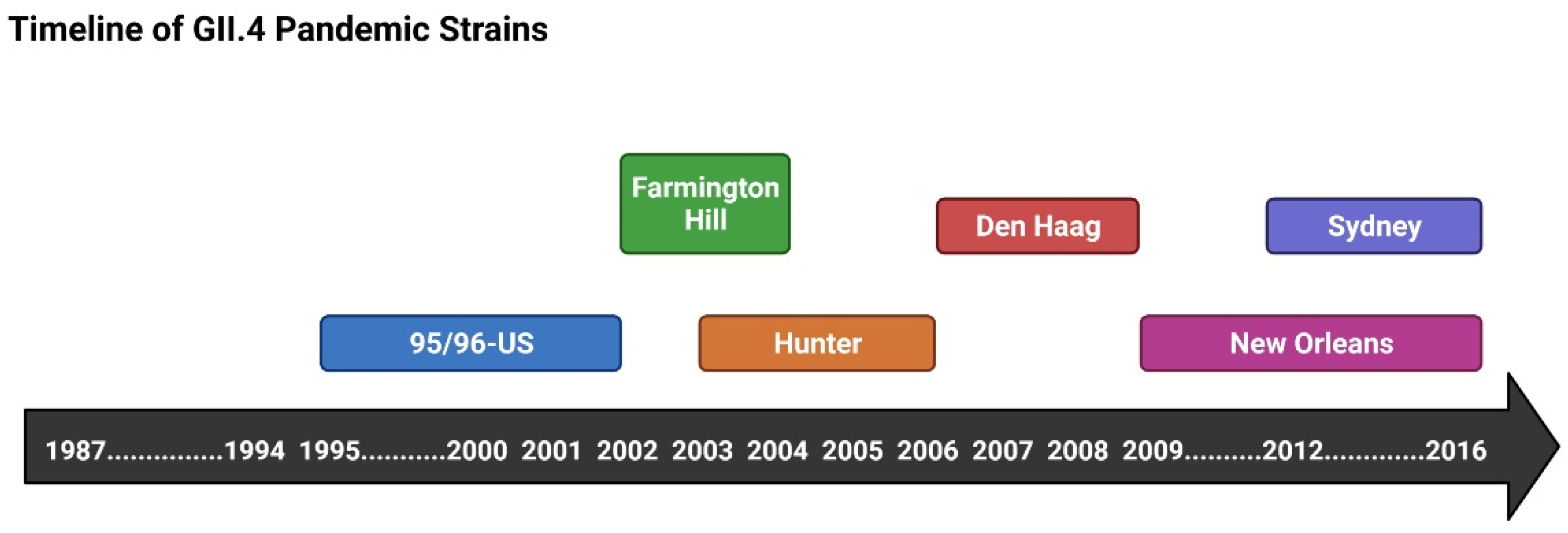
3.1. GII.4 and NoV Pandemics
3.2. Epidemiology
4. Current Hand Hygiene Methodologies for Combating NoV Transmission
5. Future of NoV Treatment
| Company | Vaccine Candidate | Administration | Antigen Form: Genotype | Clinical Trial Phase | Reference |
|---|---|---|---|---|---|
| Takeda | HIL-214 (previously TAK-214) | Intranasal, Intramuscular | NoV VLP: GI.1/GII.4 | Phase 2b | [123] |
| Vaxart | VXA-NVV-104 | Oral | Adenovirus-expressing NoV VP1NoV VLP: GI.1/GII.4 | Phase 1 | [124] |
| NVSI | Hansenulapolymorpha | Intramuscular | NoV VLP: GI.1/GII.4 | Phase 1 | [125] |
| IPS/Zhifei | Longkoma | Intramuscular | NoV VLP: GI.1/GII.3/GII.4/GII.17 | Phase 2a | [126] |
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kapikian, A.Z.; Wyatt, R.G.; Dolin, R.; Thornhill, T.S.; Kalica, A.R.; Chanock, R.M. Visualization by Immune Electron Microscopy of a 27-nm Particle Associated with Acute Infectious Nonbacterial Gastroenteritis. J. Virol. 1972, 10, 1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, M.C.; Kwan, H.S.; Chan, P.K. Structure and Genotypes of Noroviruses. Norovirus 2017, 1, 51–63. [Google Scholar]
- Lopman, B.; Armstrong, B.; Atchison, C.; Gray, J.J. Host, Weather and Virological Factors Drive Norovirus Epidemiology: Time-Series Analysis of Laboratory Surveillance Data in England and Wales. PLoS ONE 2009, 4, 6671. [Google Scholar] [CrossRef] [PubMed]
- Cubitt, W.D.; McSwiggan, D.A.; Moore, W. Winter vomiting disease caused by calicivirus. J. Clin. Pathol. 1979, 32, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Lopman, B.A.; Reacher, M.H.; Vipond, B.; Sarangi, J.; Brown, D.W. Clinical manifestation of norovirus gastroenteritis in health care settings. Clin. Infect. Dis. Off. Publ. IDSA 2004, 39, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Sandmann, F.G.; Shallcross, L.; Adams, N.; Allen, D.J.; Coen, P.G.; Jeanes, A.; Kozlakidis, Z.; Larkin, L.; Wurie, F.; Robotham, J.V.; et al. Estimating the Hospital Burden of Norovirus-Associated Gastroenteritis in England and Its Opportunity Costs for Nonadmitted Patients. Clin. Infect. Dis. 2018, 67, 693–700. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, K.M.; Sandman, F.; Allen, D.; Jarvis, C.I.; Gimma, A.; Douglas, A.; Larkin, L.; Wong, K.L.M.; Baguelin, M.; Baric, R.S.; et al. Predicted norovirus resurgence in 2021–2022 the relaxation of nonpharmaceutical interventions associated with COVID-19 restrictions in England: A mathematical modeling study. BMC Med. 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Yasmin, F.; Ali, S.H. Ullah Norovirus outbreak amid COVID-19 in the United Kingdom; priorities for achieving control. J. Med. Virol. 2022, 94, 1232. [Google Scholar] [CrossRef]
- UKHSA. National Norovirus and Rotavirus Bulletins 2021 to 2022: Management Information. 2022. Available online: https://www.gov.uk/government/statistical-data-sets/national-norovirus-and-rotavirus-bulletin-management-information--2 (accessed on 20 October 2022).
- GOVUK. Norovirus: Guidance, Data and Analysis. 2022. Available online: https://www.gov.uk/government/collections/norovirus-guidance-data-and-analysis (accessed on 23 October 2022).
- Ausar, S.F.; Foubert, T.R.; Hudson, M.H.; Vedvick, T.S.; Middaugh, C.R. Conformational stability and disassembly of Norwalk virus-like particles. Effect of pH and temperature. J. Biol. Chem. 2006, 281, 19478–19488. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Wang, M.; Wang, K.; Estes, M.K. Sequence and genomic organization of Norwalk virus. Virology 1993, 195, 51–61. [Google Scholar] [CrossRef]
- Kroneman, A.; Vega, E.; Vennema, H.; Vinjé, J.; White, P.A.; Hansman, G.; Green, K.; Martella, V.; Katayama, K.; Koopmans, M. Proposal for a unified norovirus nomenclature and genotyping. Arch. Virol. 2013, 158, 2059–2068. [Google Scholar] [CrossRef] [PubMed]
- McFadden, N.; Bailey, D.; Carrara, G.; Benson, A.; Chaudhry, Y.; Shortland, A.; Heeney, J.; Yarovinsky, F.; Simmoeternds, P.; Macdonald, A.; et al. Norovirus regulation of the innate immune response and apoptosis occurs via the product of the alternative open reading frame 4. PLoS Pathog. 2011, 7, e1002413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, B.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray crystallographic structure of the Norwalk virus capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Hutson, A.M.; Atmar, R.L.; Marcus, D.M.; Estes, M.K. Norwalk virus-like particle hemagglutination by binding to h histo-blood group antigens. J. Virol. 2003, 77, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.C.; Donaldson, E.; Leon, J.; Moe, C.L.; Frelinger, J.A.; Johnston, R.E.; Weber, D.J.; Baric, R.S. Heterotypic Humoral and Cellular Immune Responses following Norwalk Virus Infection. J. Virol. 2010, 84, 1800–1815. [Google Scholar] [CrossRef] [Green Version]
- Schroten, H.; Hanisch, F.-G.; Hansman, G.S. Human Norovirus Interactions with Histo-Blood Group Antigens and Human Milk Oligosaccharides. J. Virol. 2016, 90, 5855. [Google Scholar] [CrossRef] [Green Version]
- Bertolotti-Ciarlet, A.; Crawford, S.E.; Hutson, A.M.; Estes, M.K. The 3′ End of Norwalk Virus mRNA Contains Determinants That Regulate the Expression and Stability of the Viral Capsid Protein VP1: A Novel Function for the VP2 Protein. J. Virol. 2003, 77, 11603–11615. [Google Scholar] [CrossRef] [Green Version]
- Vongpunsawad, S.; Venkataram Prasad, B.V.; Estes, M.K. Norwalk Virus Minor Capsid Protein VP2 Associates within the VP1 Shell Domain. J. Virol. 2013, 87, 4818–4825. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Farkas, T.; Zhong, W.; Tan, M.; Thornton, S.; Morrow, A.L.; Jiang, X. Norovirus and Histo-Blood Group Antigens: Demonstration of a Wide Spectrum of Strain Specificities and Classification of Two Major Binding Groups among Multiple Binding Patterns. J. Virol. 2005, 79, 6714. [Google Scholar] [CrossRef] [Green Version]
- Campillay-Véliz, C.P.; Carvajal, J.J.; Avellaneda, A.M.; Escobar, D.; Covián, C.; Kalergis, A.M.; Lay, M.K. Human Norovirus Proteins: Implications in the Replicative Cycle, Pathogenesis, and the Host Immune Response. Front. Immunol. 2020, 11, 961. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.N.; Karst, S.M. Norovirus Mechanisms of Immune Antagonism. Curr. Opin. Virol. 2016, 16, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, R.A.; Hansman, G.S.; Clancy, L.E.; Tanaka, M.M.; Rawlinson, W.D.; White, P.A. Norovirus recombination in ORF1/ORF2 overlap. Emerg. Infect. Dis. 2005, 11, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, M.; Graham, D.Y.; Estes, M.K. Expression, self-assembly, and antigenicity of the Norwalk virus capsid protein. J. Virol. 1992, 66, 6527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, M.L.; Lindesmith, L.C.; Graham, R.L.; Baric, R.S. GII.4 Human Norovirus: Surveying the Antigenic Landscape. Viruses 2019, 11, 177. [Google Scholar] [CrossRef] [Green Version]
- Marsh, M.; Helenius, A. Virus entry: Open sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Thorne, L.G. GoodfellowG Norovirus gene expression and replication. J. Gen. Virol. 2014, 95, 278–291. [Google Scholar] [CrossRef]
- Graziano, V.R.; Wei, J.; Wilen, C.B. Norovirus attachment and entry. Viruses 2019, 11, 495. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.; Moe, C.; Marionneau, S.; Ruvoen, N.; Jiang, X.; Lindblad, L.; Stewart, P.; Lependu, J.; Baric, R. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 2003, 9, 548–553. [Google Scholar] [CrossRef]
- Nordgren, J.; Svensson, L. Genetic Susceptibility to Human Norovirus Infection: An Update. Viruses 2019, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Rouquier, S.; Lowe, J.B.; Kelly, R.J.; Fertitta, A.L.; Lennon, G.G.; Giorgi, D. Molecular Cloning of a Human Genomic Region Containing the H Blood Group α(1,2)Fucosyltransferase Gene and Two H Locus-related DNA Restriction Fragments: Isolation of A Candidate for the Human Secretor Blood Group Locus. J. Biol. Chem. 1995, 270, 4632–4639. [Google Scholar] [CrossRef] [PubMed]
- Bhar, S.; Jones, M.K. In Vitro Replication of Human Norovirus. Viruses 2019, 11, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabelli, C.; Santos-Ferreira, N.; Gillilland, I.I.I.M.G.; Cieza, R.J.; Colacino, J.A.; Sexton, J.Z.; Neyts, J.; Taube, S.; Rocha-Pereira, J.; Wobus, C.E. Human Norovirus Efficiently Replicates in Differentiated 3D-Human Intestinal Enteroids. J. Virol. 2022, 23, e0085522. [Google Scholar] [CrossRef] [PubMed]
- Borrego, F. The CD300 molecules: An emerging family of regulators of the immune system. Blood 2013, 121, 1951–1960. [Google Scholar] [CrossRef] [Green Version]
- Kolawole, A.O.; Xia, C.; Li, M.; Gamez, M.; Yu, C.; Rippinger, C.M.; Yucha, R.; Smith, T.J.; Wobus, C.E. Newly isolated mAbs broaden the neutralizing epitope in murine norovirus. J. Gen. Virol. 2014, 95, 1958–1968. [Google Scholar] [CrossRef]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldridge, M.T.; McCune, B.T.; Lee, Y.C.; Lee, S.; Pruett-Miller, S.M.; Nelson, C.E.; Fremont, D.H. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Wobus, C.E.; Karst, S.M.; Thackray, L.B.; Chang, K.O.; Sosnovtsev, S.V.; Belliot, G.; Krug, A.; Mackenzie, J.M.; Green, K.Y.; Virgin IV, H.W. Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLoS Biol. 2004, 2, e432. [Google Scholar] [CrossRef]
- Mordechai, L.; Eisenberg, M.; Newfield, T.P.; Izdebski, A.; Kay, J.E.; Poinar, H. The Justinianic Plague: An inconsequential pandemic? Proc. Natl. Acad. Sci. USA 2019, 116, 25546–25554. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.O.; Sosnovtsev, S.V.; Belliot, G.; King, A.D.; Green, K.Y. Stable expression of a Norwalk virus RNA replicon in a human hepatoma cell line. Virol 2006, 353, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Gerondopoulos, A.; Jackson, T.; Monaghan, P.; Doyle, N.; Roberts, L.O. Murine norovirus-1 cell entry is mediated through a non-clathrin-, non-caveolae-, dynamin- And cholesterol-dependent pathway. J. Gen. Virol. 2010, 91, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.W.; Wobus, C.E. Endocytosis of Murine Norovirus 1 into Murine Macrophages Is Dependent on Dynamin II and Cholesterol. J. Virol. 2010, 84, 6163. [Google Scholar] [CrossRef] [PubMed]
- Daughenbaugh, K.F.; Fraser, C.S.; Hershey, J.W.; Hardy, M.E. The genome-linked protein VPg of the Norwalk virus binds eIF3, suggesting its role in translation initiation complex recruitment. EMBO J. 2003, 22, 2852–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodfellow, I.; Chaudhry, Y.; Gioldasi, I.; Gerondopoulos, A.; Natoni, A.; Labrie, L.; Laliberté, J.F.; Roberts, L. Calicivirus translation initiation requires an interaction between VPg and eIF4E. EMBO Rep. 2005, 6, 968–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunus, M.A.; Yunus, M.A. Molecular Mechanisms for Norovirus Genome Replication. Norovirus 2021, 23, 278–291. [Google Scholar]
- Green, K.Y.; Belliot, G.; Taylor, J.L.; Valdesuso, J.; Lew, J.F.; Kapikian, A.Z.; Lin, F.Y.C. A Predominant Role for Norwalk-like Viruses as Agents of Epidemic Gastroenteritis in Maryland Nursing Homes for the Elderly. J. Infect. Dis. 2002, 185, 133–146. [Google Scholar] [CrossRef]
- Rohayem, J.; Robel, I.; Jäger, K.; Scheffler, U.; Rudolph, W. Protein-Primed and De Novo Initiation of RNA Synthesis by Norovirus 3D pol. J. Virol. 2006, 80, 7060–7069. [Google Scholar] [CrossRef] [Green Version]
- Subba-Reddy, C.V.; Yunus, M.A.; Goodfellow, G.; Kao, C.C. Norovirus RNA synthesis is modulated by an interaction between the viral RNA-dependent RNA polymerase and the major capsid protein, VP1. J. Virol. 2012, 86, 10138–10149. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, P.; Karakasiliotis, I.; Bailey, D.; Chaudhry, Y.; Evans, D.J.; Goodfellow, G. Bioinformatic and functional analysis of RNA secondary structure elements among different genera of human and animal caliciviruses. Nucleic Acids Res. 2008, 36, 2530–2546. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, Y.; Nayak, A.; Bordeleau, M.E.; Tanaka, J.; Pelletier, J.; Belsham, G.J.; Roberts, L.O.; Goodfellow, I.G. Caliciviruses differ in their functional requirements for eIF4F components. J. Biol. Chem. 2006, 281, 25315–25325. [Google Scholar] [CrossRef] [Green Version]
- Guix, S.; Asanaka, M.; Katayama, K.; Crawford, S.E.; Neill, F.H.; Atmar, R.L.; Estes, M.K. Norwalk virus RNA is infectious in mammalian cells. J. Virol. 2007, 81, 12238–12248. [Google Scholar] [CrossRef] [Green Version]
- Subba-Reddy, C.V.; Goodfellow, I.; Kao, C.C. VPg-Primed RNA Synthesis of Norovirus RNA-Dependent RNA Polymerases by Using a Novel Cell-Based Assay. J. Virol. 2011, 85, 13027–13037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosnovtsev, S.V.; Belliot, G.; Chang, K.O.; Onwudiwe, O.; Green, K.Y. Feline Calicivirus VP2 Is Essential for the Production of Infectious Virions. J. Virol. 2005, 79, 4012. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Oviedo, J.M.; Martín-Alonso, J.M.; Díaz, E.; Boga, J.A.; Parra, F. Programmed cell death in the pathogenesis of rabbit hemorrhagic disease. Arch. Virol. 1998, 143, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Sosnovtsev, S.V.; Prikhod’ko, E.A.; Belliot, G.; Cohen, J.I.; Green, K.Y. Feline calicivirus replication induces apoptosis in cultured cells. Virus Res. 2003, 94, 1–10. [Google Scholar] [CrossRef]
- Bok, K.; Prikhodko, V.G.; Green, K.Y.; Sosnovtsev, S.V. Apoptosis in Murine Norovirus-Infected RAW264.7 Cells Is Associated with Downregulation of Survivin. J. Virol. 2009, 83, 3647–3656. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Norovirus Outbreak Linked to Raw Oysters from British Columbia—Norovirus CDC. 2022. Available online: https://www.cdc.gov/norovirus/outbreaks/index.html (accessed on 28 October 2022).
- Centres for Disease Control and Prevention. Vital Signs: Foodborne Norovirus Outbreaks—United States, 2009–2012. 2011. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm63223.htm (accessed on 1 November 2022).
- Nikitin, N.; Petrova, E.; Trifonova, E.; Karpova, O. Influenza virus aerosols in the air and their infectiousness. Adv. Virol. 2014, 2014, 859090. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.A.; Bruggink, L.D. The Dynamics of Norovirus Outbreak Epidemics: Recent Insights. Int. J. Environ. Res. Public Health 2011, 8, 1141. [Google Scholar] [CrossRef] [Green Version]
- Tian, P.; Engelbrektson, A.L.; Jiang, X.; Zhong, W.; Mandrell, R.E. Norovirus Recognizes Histo-Blood Group Antigens on Gastrointestinal Cells of Clams, Mussels, and Oysters: A Possible Mechanism of Bioaccumulation. J. Food. Prot. 2007, 70, 2140–2147. [Google Scholar] [CrossRef] [Green Version]
- CDC. When and How to Wash Your Hands|Handwashing|CDC Centers for Disease Control and Prevention. 2022. Available online: https://www.cdc.gov/handwashing/when-how-handwashing.html (accessed on 1 November 2022).
- Hassard, F.; Sharp, J.H.; Taft, H.; LeVay, L.; Harris, J.P.; McDonald, J.E.; Tuson, K.; Wilson, J.; Jones, D.L.; Malham, S.K. Critical Review on the Public Health Impact of Norovirus Contamination in Shellfish and the Environment: A UK Perspective. Food Environ. Virol. 2017, 9, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Rockx, B.; De Wit, M.; Vennema, H.; Vinjé, J.; De Bruin, E.; Van Duynhoven, Y.; Koopmans, M. Natural history of human Calicivirus infection: A prospective cohort study. Clin. Infect. Dis. 2002, 35, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Glass, R.I.; Parashar, U.D.; Estes, M.K. Norovirus Gastroenteritis. N. Engl. J. Med. 2009, 361, 1776–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robilotti, E.; Deresinski, S.; Pinsky, B.A. Norovirus. Clin. Microbiol. Rev. 2015, 28, 134–164. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Katsushima, N.; Mizuta, K.; Muraki, Y.; Hongo, S.; Matsuzaki, Y. Prolonged norovirus shedding in infants ≤6 months of age with gastroenteritis. J. Pediatr. Infect. 2007, 26, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Desai, R.; Hembree, C.D.; Handel, A.; Matthews, J.E.; Dickey, B.W.; Mcdonald, S.; Hall, A.J.; Parashar, U.D.; Leon, J.S.; Lopman, B. Severe outcomes are associated with genogroup 2 genotype 4 norovirus outbreaks: A systematic literature review. Cliln. Infect. Dis. 2012, 55, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Asten, L.; Van Den Wijngaard, C.; Van Pelt, W.; Van De Kassteele, J.; Meijer, A.; Van Der Hoek, W.; Kretzschmar, M.; Koopmans, M. Mortality attributable to 9 common infections: Significant effect of influenza A, respiratory syncytial virus, influenza B, norovirus, and parainfluenza in elderly persons. J. Infect. Dis. 2012, 206, 628–639. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.; Forde, D.; Vedio, A.; Cope, A.; Pratt, G.; Tunbridge, A. ‘It’s just a virus’—Viral illness in older people: Prevention and management. Rev. Clin. Gerontol. 2013, 23, 131–141. [Google Scholar] [CrossRef]
- Haddadin, Z.; Batarseh, E.; Hamdan, L.; Stewart, L.S.; Piya, B.; Rahman, H.; Spieker, A.J.; Chappell, J.; Wikswo, M.E.; Dunn, J.R.; et al. Characteristics of GII.4 Norovirus Versus Other Genotypes in Sporadic Pediatric Infections in Davidson County, Tennessee, USA. Clin. Infect. Dis. 2021, 73, e1525–e1531. [Google Scholar] [CrossRef]
- Newman, K.L.; Leon, J.S. Norovirus immunology: Of mice and mechanisms. Eur. J. Immunol. 2015, 45, 2742–2757. [Google Scholar] [CrossRef] [Green Version]
- Ponterio, E.; Petrizzo, A.; Di Bartolo, I.; Buonaguro, F.M.; Buonaguro, L.; Ruggeri, F.M. Pattern of activation of human antigen presenting cells by genotype GII.4 norovirus virus-like particles. J. Transl. Med. 2013, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Newman, K.L.; Moe, C.L.; Kirby, A.E.; Flanders, W.D.; Parkos, C.A.; Leon, J.S. Human norovirus infection and the acute serum cytokine response. Clin. Exp. Immunol. 2015, 182, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Ando, T.; Monroe, S.S.; Gentsch, J.R.; Jin, Q.; Lewis, D.C.; Glass, R.I. Detection and differentiation of antigenically distinct small round-structured viruses (Norwalk-like viruses) by reverse transcription-PCR and southern hybridization. J. Clin. Microbiol. 1995, 33, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinjé, J.; Green, J.; Lewis, D.C.; Gallimore, C.I.; Brown, D.W.; Koopmans, M.P. Genetic polymorphism across regions of the three open reading frames of “Norwalk-like viruses”. Arch. Virol. 2000, 145, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.P.; Ando, T.; Fankhauser, R.L.; Beard, R.S.; Glass, R.I.; Monroe, S.S. Norovirus classification and proposed strain nomenclature. Virol 2006, 346, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Vinjé, J. Advances in laboratory methods for detection and typing of norovirus. J. Clin. Microbiol. 2015, 53, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.; Green, K.; Martella, V.; Wang, Q.; White, P.E.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Wobus, C.E.; Thackray, L.B.; Herbert, W. Virgin Murine Norovirus: A Model System To Study Norovirus Biology and Pathogenesis. J. Virol. 2006, 80, 5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.L.; Barclay, L.; Collins, N.R.; Wikswo, M.E.; Castro, C.J.; Magaña, L.C.; Gregoricus, N.; Marine, R.L.; Chhabra, P.; Vinjé, J. Genetic and Epidemiologic Trends of Norovirus Outbreaks in the United States from 2013 2016 Emergence of Novel GII.4 Recombinant Viruses. J. Clin. Microbiol. 2017, 55, 2208. [Google Scholar] [CrossRef] [Green Version]
- Estienney, M.; Tarris, G.; Abou-Hamad, N.; Rouleau, A.; Boireau, W.; Chassagnon, R.; Ayouni, S.; Daval-Frerot, P.; Martin, L.; Bouyer, F.; et al. Epidemiological Impact of GII.17 Human Noroviruses Associated With Attachment to Enterocytes. Front. Microbiol. 2022, 13, 1270. [Google Scholar] [CrossRef]
- Bull, R.A.; White, P.A. Mechanisms of GII.4 norovirus evolution. Tren. Microbil. 2011, 19, 233–240. [Google Scholar] [CrossRef]
- Eden, J.-S.; Tanaka, M.M.; Boni, M.F.; Rawlinson, W.D.; White, P.A. Recombination within the Pandemic Norovirus GII.4 Lineage. J. Virol. 2013, 87, 6270. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Wang, W.B.; Zhang, J.; Hou, J.; Tang, F.; Zhang, X.F.; Du, L.F.; Su, J.G.; Li, Q.M. Evolution of the interactions between GII.4 noroviruses and histo-blood group antigens: Insights from experimental and computational studies. PLoS Pathog. 2021, 17, e1009745. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Eden, J.S.; Rawlinson, W.D.; White, P.A. Rapid Evolution of Pandemic Noroviruses of the GII.4 Lineage. PLoS Pathog. 2010, 6, e1000831. [Google Scholar] [CrossRef]
- Yu, J.M.; Liang, Z.Y.; Guo, K.; Sun, X.M.; Zhang, Q.; Dong, Y.J.; Duan, Z.J. Intra-Host Evolution of Norovirus GII.4 in a Chronic Infected Patient With Hematopoietic Stem Cell Transplantation. Front. Microbiol. 2020, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doshi, M.; Woodwell, S.; Kelleher, K.; Mangan, K.; Axelrod, P. An outbreak of norovirus infection in a bone marrow transplant unit. Am. J. Infect. Dis. 2013, 41, 820–823. [Google Scholar] [CrossRef]
- Schwartz, S.; Vergoulidou, M.; Schreier, E.; Loddenkemper, C.; Reinwald, M.; Schmidt-Hieber, M.; Flegel, W.A.; Thiel, E.; Schneider, T. Norovirus gastroenteritis causes severe and lethal complications after chemotherapy and hematopoietic stem cell transplantation. Blood 2011, 117, 5850–5856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, J.S.; Fankhauser, R.L.; Ando, T.; Monroe, S.S.; Glass, R.I. Identification of a distinct common strain of “Norwalk-like viruses” having a global distribution. J. Infect. Dis. 1999, 179, 1334–1344. [Google Scholar] [CrossRef] [Green Version]
- Pringle, K.; Lopman, B.; Vega, E.; Vinje, J.; Parashar, U.D.; Hall, A.J. Noroviruses: Epidemiology, immunity and prospects for prevention. Future Microbiol. 2015, 10, 53–67. [Google Scholar] [CrossRef]
- Lopman, B.; Vennema, H.; Kohli, E.; Pothier, P.; Sanchez, A.; Negredo, A.; Buesa, J.; Schreier, E.; Reacher, M.; Brown, D.; et al. Increase in viral gastroenteritis outbreaks in Europe and epidemic spread of new norovirus variant. Lancet 2004, 363, 682–688. [Google Scholar] [CrossRef]
- Bull, R.A.; Tu, E.T.; McIver, C.J.; Rawlinson, W.D.; White, P.A. Emergence of a new norovirus genotype II.4 variant associated with global outbreaks of gastroenteritis. J. Clin. Microbiol. 2006, 44, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Holland, D.; Thomson, L.; Mahmoudzadeh, N.; Khaled, A. Estimating deaths from foodborne disease in the UK for 11 key pathogens. BMJ Open Gastro 2020, 7, 377. [Google Scholar] [CrossRef]
- Cyrus, S.; Ho, C.Y. Mental Health Strategies to Combat the Psychological Impact of Coronavirus Disease 2019 (COVID-19) Beyond Paranoia and Panic. Ann. Acad. Med. Singap. 2020, 49, 155–160. [Google Scholar]
- Douglas, A.; Sandmann, F.G.; Allen, D.J.; Celma, C.C.; Beard, S.; Larkin, L. Impact of COVID-19 on national surveillance of norovirus in England and potential risk of increased disease activity in 2021. J. Hosp. Infect. 2021, 112, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G. Efficacy of ethanol against viruses in hand disinfection. J. Hosp. Infect. 2018, 98, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M. Pathogenesis of Noroviruses, Emerging RNA Viruses. Viruses 2010, 2, 748. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Kambhampati, A.; Koopmans, M.; Lopman, B.A. Burden of norovirus in healthcare facilities and strategies for outbreak control. J. Hosp. Infect. 2015, 89, 296–301. [Google Scholar] [CrossRef] [Green Version]
- Frange, P.; Touzot, F.; Debré, M.; Héritier, S.; Leruez-Ville, M.; Cros, G.; Rouzioux, C.; Blanche, S.; Fischer, A.; Avettand-Fenoël, V. Prevalence and clinical impact of norovirus fecal shedding in children with inherited immune deficiencies. J. Infect. Dis. 2012, 206, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, S.S.; Green, K.Y.; Korba, B.E. Treatment of norovirus infections: Moving antivirals from the bench to the bedside. Antivir. Res. 2014, 105, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Munir, N.; Liu, P.; Gastañaduy, P.; Montes, J.; Shane, A.; Moe, C. Norovirus infection in immunocompromised children and children with hospital-acquired acute gastroenteritis. J. Med. Virol. 2014, 86, 1203–1209. [Google Scholar] [CrossRef]
- Harris, J.P.; Lopman, B.A.; Cooper, B.S.; O’Brien, S.J. Does spatial proximity drive norovirus transmission during outbreaks in hospitals? BMJ Open 2013, 3, e003060. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.P.; Qiu, H.; Ticehurst, J.R.; Dickson, C.; Rosenbaum, P.; Lawson, P.; Stokes, A.B.; Lowenstein, C.; Kaminsky, M.; Cosgrove, S.E.; et al. Outbreak management and implications of a nosocomial norovirus outbreak. Clin. Infect. Dis. 2007, 45, 534–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhrie, F.H.; Teunis, P.; Vennema, H.; Copra, C.; Thijs Beersma, M.F.; Bogerman, J.; Koopmans, M. Nosocomial transmission of norovirus is mainly caused by symptomatic cases. Clin. Infect. Dis. 2012, 54, 931–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartsch, S.M.; Lopman, B.A.; Hall, A.J.; Parashar, U.D.; Lee, B.Y. The potential economic value of a human norovirus vaccine for the United States. Vaccine. 2012, 30, 7097–7104. [Google Scholar] [CrossRef] [PubMed]
- Health Protection Agency. Guidelines for the Management of Norovirus Outbreaks in Acute and Community Health and Social Care Settings. 2012. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/322943/Guidance_for_managing_norovirus_outbreaks_in_healthcare_settings.pdf (accessed on 24 October 2022).
- Maccannell, T.; Umscheid, C.A.; Agarwal, R.K.; Agarwal, K.; Lee, I.; Kuntz, G.; Stevenson, K. Guideline for the prevention and control of norovirus gastroenteritis outbreaks in healthcare settings. Infect. Control. Hosp. Epidemiol. 2015, 32, 939–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feliciano, L.; Li, J.; Lee, J.; Pascall, M.A. Efficacies of Sodium Hypochlorite and Quaternary Ammonium Sanitizers for Reduction of Norovirus and Selected Bacteria during Ware-Washing Operations. PLoS ONE 2012, 7, e50273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Yuen, Y.; Hsiao, H.M.; Jaykus, L.A.; Moe, C. Effectiveness of Liquid Soap and Hand Sanitizer against Norwalk Virus on Contaminated Hands. Appl. Environ. Microbiol. 2010, 76, 394. [Google Scholar] [CrossRef] [Green Version]
- Predmore, A.; Li, J. Enhanced Removal of a Human Norovirus Surrogate from Fresh Vegetables and Fruits by a Combination of Surfactants and Sanitizers. Appl. Environ. Microbiol. 2011, 77, 4829. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; Ferris, M.T.; Mullan, C.W.; Ferreira, J.; Debbink, K.; Swanstrom, J.; Richardson, C.; Goodwin, R.R.; Baehner, F.; Mendelman, P.M.; et al. Broad blockade antibody responses in human volunteers after immunization with a multivalent norovirus VLP candidate vaccine: Immunological analyses from a phase I clinical trial. PLoS Med. 2015, 12, e1001807. [Google Scholar] [CrossRef] [Green Version]
- Czakó, R.; Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Graham, D.Y.; Estes, M.K. Experimental human infection with norwalk virus elicits a surrogate neutralizing antibody response with cross-genogroup activity. Clin. Vaccine Immunol. 2015, 22, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Koelle, K.; Cobey, S.; Grenfell, B.; Pascual, M. Epochal evolution shapes the phylodynamics of interpandemic influenza a (H3N2) in humans. Science 2006, 314, 1898–1903. [Google Scholar] [CrossRef]
- Atmar, R.L.; Bernstein, D.I.; Harro, C.D.; Al-Ibrahim, M.S.; Chen, W.H.; Ferreira, J.; Estes, M.; Graham, D.; Opekun, A.; Richardson, C.; et al. Norovirus Vaccine against Experimental Human Norwalk Virus Illness. N. Engl. J. Med. 2011, 365, 2178–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M. Norovirus Vaccines: Current Clinical Development and Challenges. Pathogens 2021, 10, 1641. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wang, Z.; Bian, X.; Du, X.; Wei, C. Folate-modified doxorubicin-loaded nanoparticles for tumor-targeted therapy. Pharm. Biol. 2014, 14, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, J.; Mendelman, P.M.; Lloyd, E.; Liu, M.; Boslego, J.; Borkowski, A.; Jackson, A.; Faix, D.; on behalf of the US Navy Study Team. Efficacy of an intramuscular bivalent norovirus GI.1/GII.4 virus-like particle vaccine candidate in healthy US adults. Vaccine. 2020, 38, 6442–6449. [Google Scholar] [CrossRef]
- Weerasekara, S.; Prior, A.M.; Hua, D.H. Current Tools for Norovirus Drug Discovery. Expert Opin. Drug Descov. 2016, 11, 529. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Jiang, X. Library screen for inhibitors targeting norovirus binding to histo-blood group antigen receptors. Antimicrob. Agents Chemother. 2007, 51, 324–331. [Google Scholar] [CrossRef]
- Takeda. Long-Term Immunogenicity of the Norovirus GI.1/GII.4 Bivalent Virus-like Particle (VLP) Vaccine (NoV Vaccine) in Adults. Clinicaltrials. 2017. Available online: https://clinicaltrials.gov/ct2/show/NCT03039790h (accessed on 2 November 2022).
- Vaxart. Ph 1b: Safety & Immunogenicity of Ad5 Based Oral Norovirus Vaccine (VXA-NVV-104). Clinicaltrials. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT04854746h (accessed on 2 November 2022).
- National Vaccine Serum Institute, C. Clinical Trial to Evaluate the Recombinant Norovirus Bivalent (GI. 1/GII. 4) Vaccine (Hansenula Polymorpha). Clinicaltrials. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT04941261h (accessed on 1 November 2022).
- Takeda. Safety and Immunogenicity of Norovirus GI.1/GII.4 Bivalent Virus-Like Particle Vaccine in an Elderly Population. Clinicaltrials. 2016. Available online: https://www.clinicaltrials.gov/ct2/show/NCT02661490 (accessed on 31 October 2022).









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winder, N.; Gohar, S.; Muthana, M. Norovirus: An Overview of Virology and Preventative Measures. Viruses 2022, 14, 2811. https://doi.org/10.3390/v14122811
Winder N, Gohar S, Muthana M. Norovirus: An Overview of Virology and Preventative Measures. Viruses. 2022; 14(12):2811. https://doi.org/10.3390/v14122811
Chicago/Turabian StyleWinder, Natalie, Sara Gohar, and Munitta Muthana. 2022. "Norovirus: An Overview of Virology and Preventative Measures" Viruses 14, no. 12: 2811. https://doi.org/10.3390/v14122811