
Associations between NK Cells in Different Immune Organs and Cellular SIV DNA and RNA in Regional HLADR− CD4+ T Cells in Chronically SIVmac239-Infected, Treatment-Naïve Rhesus Macaques

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Viral Challenge
2.2. Necropsy and Sample Collection
2.3. Plasma and Tissue Viral Load Analyses
2.4. NK Cell Stimulation and Functional Assays
2.5. Flow Cytometry and Antibodies
2.6. Statistical Analyses
3. Results
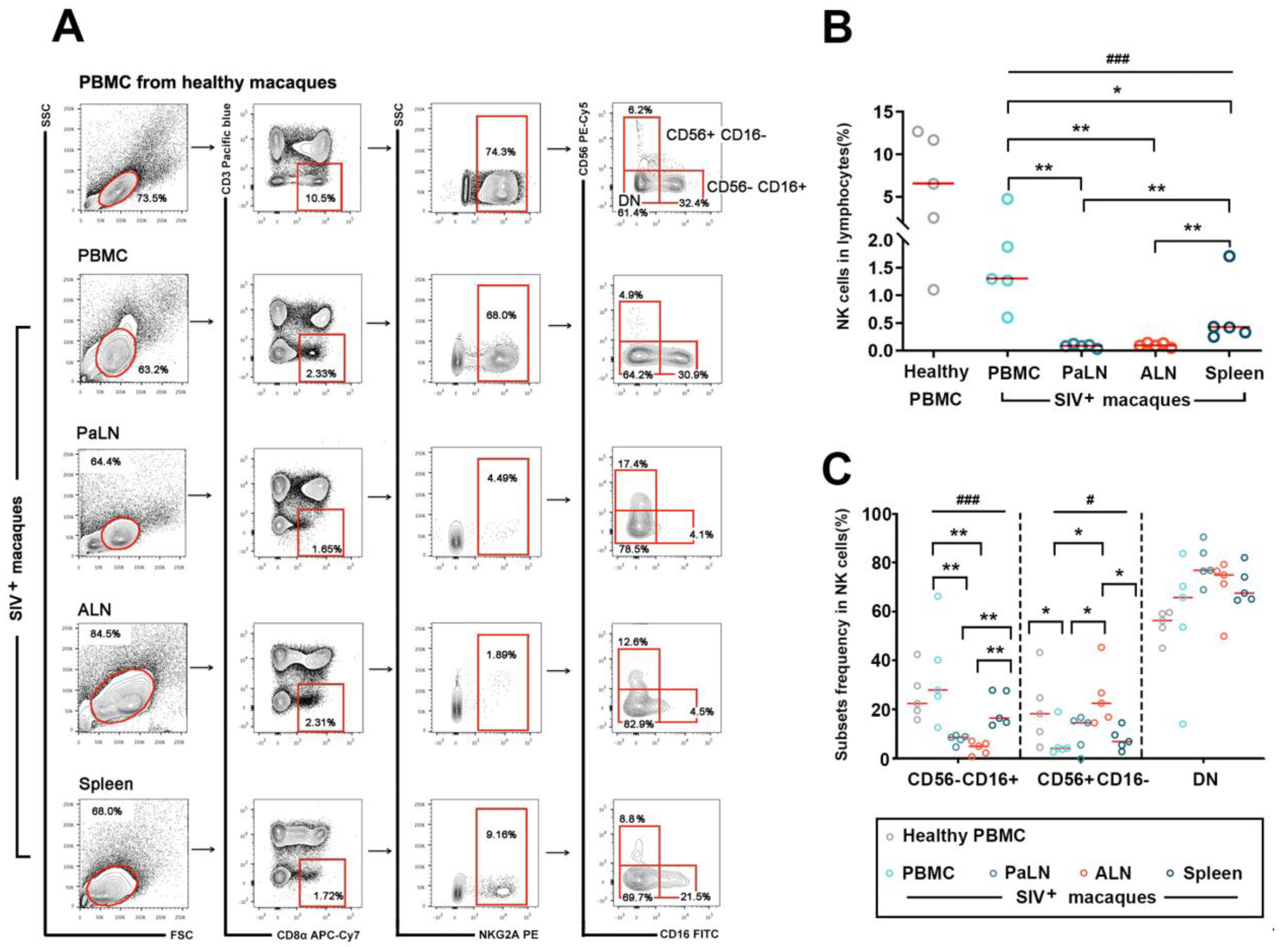
3.1. Distributional Characteristics of NK Cells in the Peripheral Blood and Secondary Lymphoid Organs in Chronic SIVmac239-Infected Rhesus Macaques
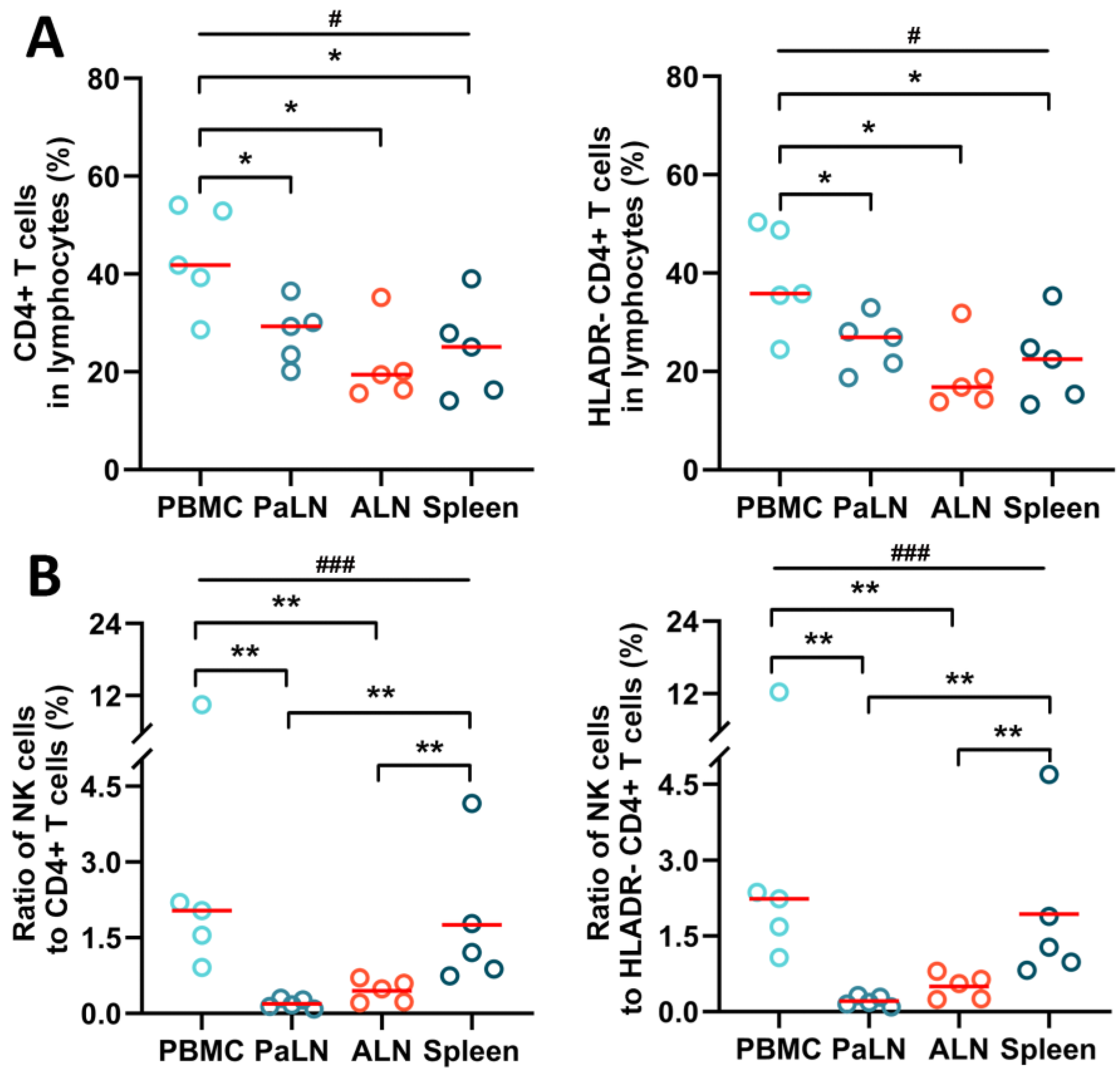
3.2. NK Cells and CD4+ T Cells in Peripheral Blood and Secondary Lymphoid Organs of SIV Chronically Infected Macaques
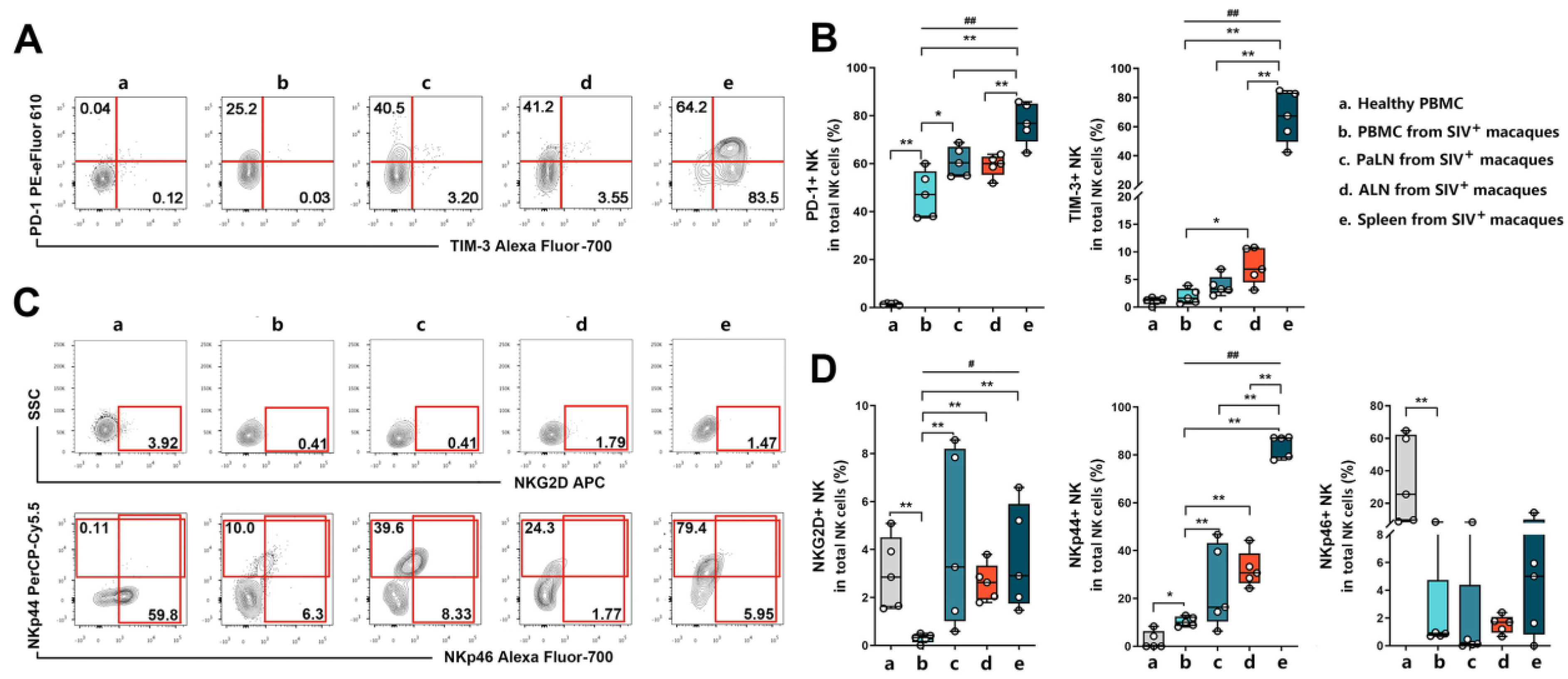
3.3. Expression of NK Cell Surface Phenotypes in Peripheral Blood and Secondary Lymphoid Organs of SIV Chronically Infected Macaques
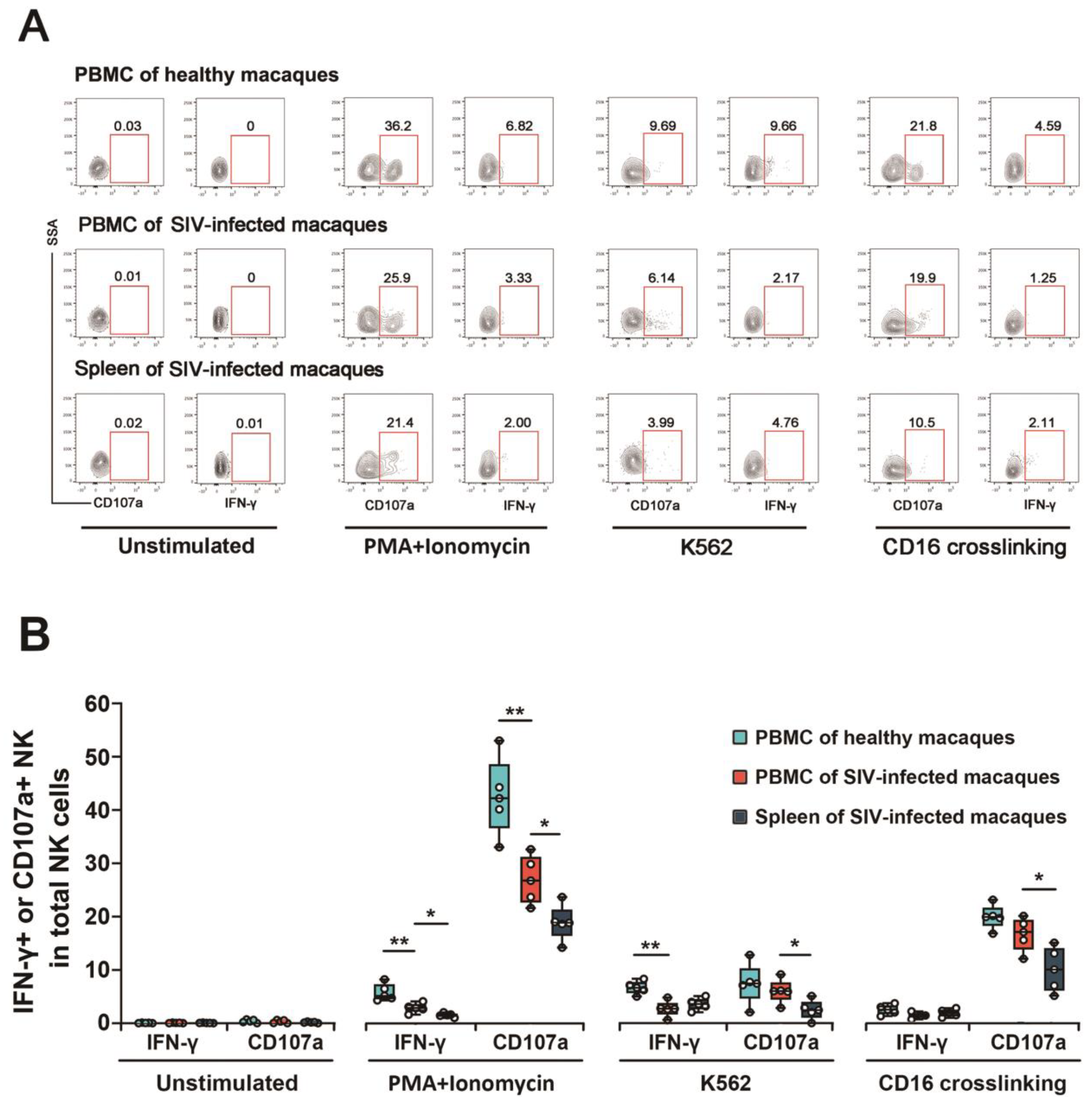
3.4. NK Cytotoxic Function Was Impaired in SIV Chronically Infected Macaques
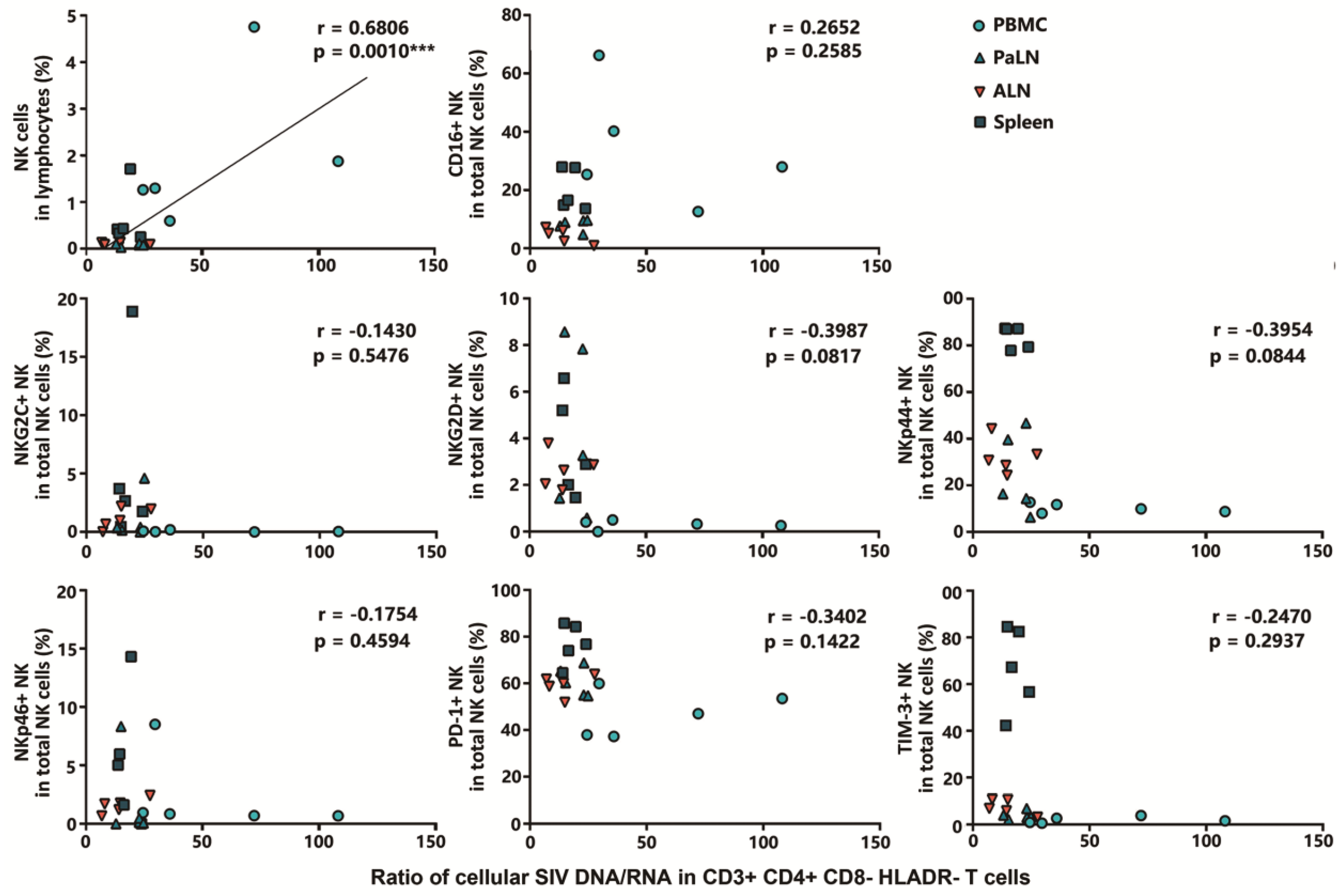
3.5. Positive Correlation between NK Cell Frequency in Lymphocytes and Cellular SIV DNA/RNA Ratio in HLADR− CD4+ T Cells
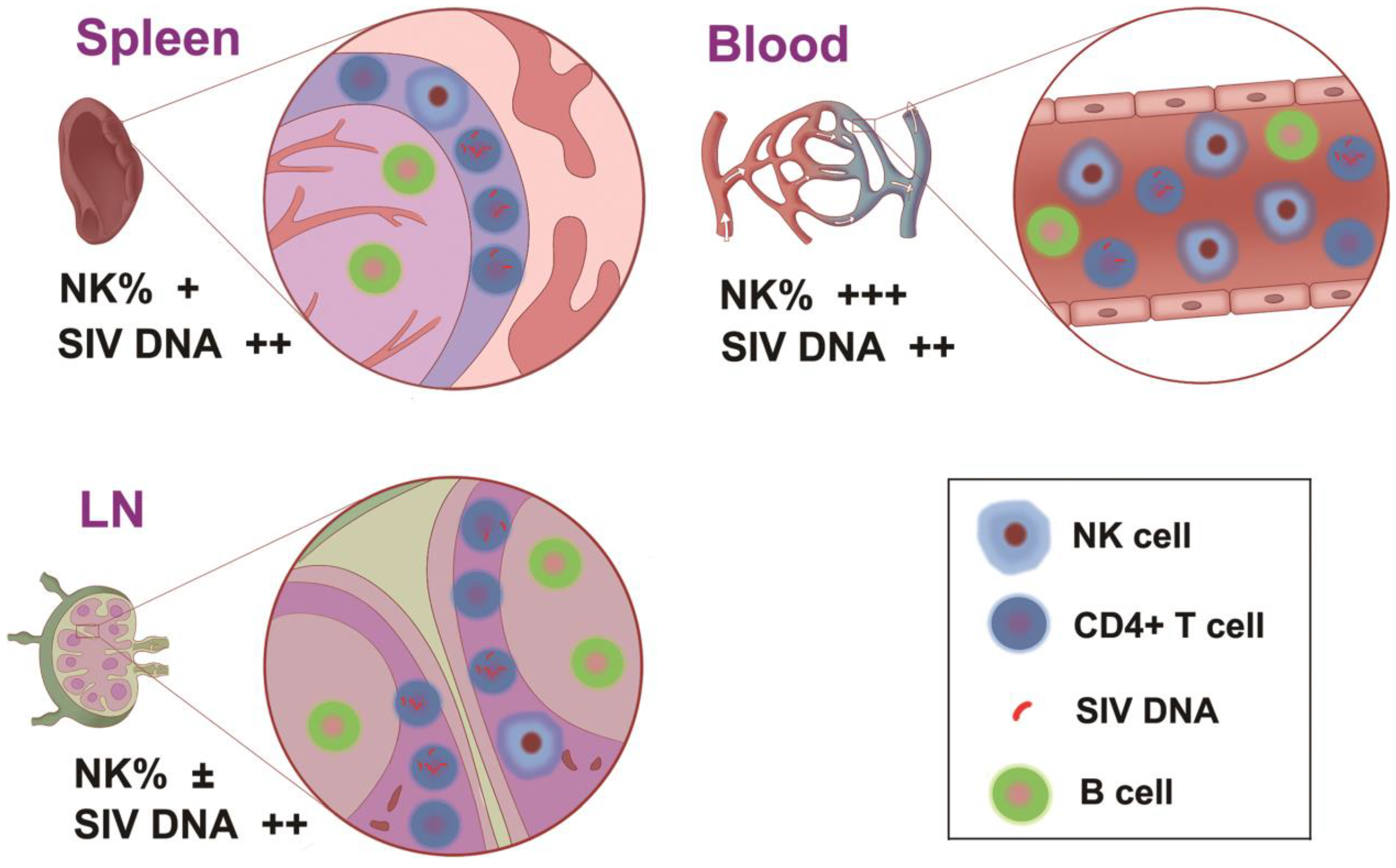
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleinman, A.J.; Pandrea, I.; Apetrei, C. So Pathogenic or So What?-A Brief Overview of SIV Pathogenesis with an Emphasis on Cure Research. Viruses 2022, 14, 135. [Google Scholar] [CrossRef] [PubMed]
- Castro-Gonzalez, S.; Colomer-Lluch, M.; Serra-Moreno, R. Barriers for HIV Cure: The Latent Reservoir. AIDS Res. Hum. Retrovir. 2018, 34, 739–759. [Google Scholar] [CrossRef]
- Bailon, L.; Mothe, B.; Berman, L.; Brander, C. Novel Approaches towards a Functional Cure of HIV/AIDS. Drugs 2020, 80, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Huot, N.; Bosinger, S.E.; Paiardini, M.; Reeves, R.K.; Müller-Trutwin, M. Lymph Node Cellular and Viral Dynamics in Natural Hosts and Impact for HIV Cure Strategies. Front. Immunol. 2018, 9, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huot, N.; Rascle, P.; Garcia-Tellez, T.; Jacquelin, B.; Müller-Trutwin, M. Innate immune cell responses in non pathogenic versus pathogenic SIV infections. Curr. Opin. Virol. 2016, 19, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, Y.; Lum, R.; Okoye, A.A.; Park, H.; Matsuda, K.; Bae, J.Y.; Hagen, S.I.; Shoemaker, R.; Deleage, C.; Lucero, C.; et al. B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers. Nat. Med. 2015, 21, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Cantero-Pérez, J.; Grau-Expósito, J.; Serra-Peinado, C.; Rosero, D.A.; Luque-Ballesteros, L.; Astorga-Gamaza, A.; Castellví, J.; Sanhueza, T.; Tapia, G.; Lloveras, B.; et al. Resident memory T cells are a cellular reservoir for HIV in the cervical mucosa. Nat. Commun. 2019, 10, 4739. [Google Scholar] [CrossRef] [Green Version]
- Whitney, J.B.; Hill, A.L.; Sanisetty, S.; Penaloza-MacMaster, P.; Liu, J.; Shetty, M.; Parenteau, L.; Cabral, C.; Shields, J.; Blackmore, S.; et al. Rapid seeding of the viral reservoir prior to SIV viraemia in rhesus monkeys. Nature 2014, 512, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Whitney, J.B.; Lim, S.Y.; Osuna, C.E.; Kublin, J.L.; Chen, E.; Yoon, G.; Liu, P.T.; Abbink, P.; Borducci, E.N.; Hill, A.; et al. Prevention of SIVmac251 reservoir seeding in rhesus monkeys by early antiretroviral therapy. Nat. Commun. 2018, 9, 5429. [Google Scholar] [CrossRef] [Green Version]
- Eisele, E.; Siliciano, R.F. Redefining the viral reservoirs that prevent HIV-1 eradication. Immunity 2012, 37, 377–388. [Google Scholar] [CrossRef]
- Churchill, M.J.; Deeks, S.G.; Margolis, D.M.; Siliciano, R.F.; Swanstrom, R. HIV reservoirs: What, where and how to target them. Nat. Rev. Microbiol. 2016, 14, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Manickam, C.; Shah, S.V.; Nohara, J.; Ferrari, G.; Reeves, R.K. Monkeying Around: Using Non-human Primate Models to Study NK Cell Biology in HIV Infections. Front. Immunol. 2019, 10, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flórez-Álvarez, L.; Hernandez, J.C.; Zapata, W. NK Cells in HIV-1 Infection: From Basic Science to Vaccine Strategies. Front. Immunol. 2018, 9, 2290. [Google Scholar] [CrossRef] [Green Version]
- Forthal, D.N.; Finzi, A. Antibody-dependent cellular cytotoxicity in HIV infection. AIDS (Lond. Engl.) 2018, 32, 2439–2451. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.Y.; Pegu, A.; Rudicell, R.S.; Yang, Z.Y.; Joyce, M.G.; Chen, X.; Wang, K.; Bao, S.; Kraemer, T.D.; Rath, T.; et al. Enhanced neonatal Fc receptor function improves protection against primate SHIV infection. Nature 2014, 514, 642–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, J.; Balkow, S.; Anel, A.; Simon, M.M. Granzymes are essential for natural killer cell-mediated and perf-facilitated tumor control. Eur. J. Immunol. 2002, 32, 2881–2887. [Google Scholar] [CrossRef]
- Street, S.E.; Hayakawa, Y.; Zhan, Y.; Lew, A.M.; MacGregor, D.; Jamieson, A.M.; Diefenbach, A.; Yagita, H.; Godfrey, D.I.; Smyth, M.J. Innate immune surveillance of spontaneous B cell lymphomas by natural killer cells and gammadelta T cells. J. Exp. Med. 2004, 199, 879–884. [Google Scholar] [CrossRef]
- Robertson, M.J. Role of chemokines in the biology of natural killer cells. J. Leukoc. Biol. 2002, 71, 173–183. [Google Scholar] [CrossRef]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef] [Green Version]
- Huot, N.; Jacquelin, B.; Garcia-Tellez, T.; Rascle, P.; Ploquin, M.J.; Madec, Y.; Reeves, R.K.; Derreudre-Bosquet, N.; Müller-Trutwin, M. Natural killer cells migrate into and control simian immunodeficiency virus replication in lymph node follicles in African green monkeys. Nat. Med. 2017, 23, 1277–1286. [Google Scholar] [CrossRef]
- Jacquelin, B.; Petitjean, G.; Kunkel, D.; Liovat, A.S.; Jochems, S.P.; Rogers, K.A.; Ploquin, M.J.; Madec, Y.; Barré-Sinoussi, F.; Dereuddre-Bosquet, N.; et al. Innate immune responses and rapid control of inflammation in African green monkeys treated or not with interferon-alpha during primary SIVagm infection. PLoS Pathog. 2014, 10, e1004241. [Google Scholar] [CrossRef] [PubMed]
- Terrade, G.; Huot, N.; Petitdemange, C.; Lazzerini, M.; Orta Resendiz, A.; Jacquelin, B.; Müller-Trutwin, M. Interests of the Non-Human Primate Models for HIV Cure Research. Vaccines 2021, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Tuyishime, M.; Garrido, C.; Jha, S.; Moeser, M.; Mielke, D.; LaBranche, C.; Montefiori, D.; Haynes, B.F.; Joseph, S.; Margolis, D.M.; et al. Improved killing of HIV-infected cells using three neutralizing and non-neutralizing antibodies. J. Clin. Investig. 2020, 130, 5157–5170. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Zhang, W.; Hwang, J.; An, E.-K.; Choi, Y.K.; Moon, E.; Loznik, M.; Huh, Y.H.; Herrmann, A.; Kwak, M.; et al. Carrier-free micellar CpG interacting with cell membrane for enhanced immunological treatment of HIV-1. Biomaterials 2021, 277, 121081. [Google Scholar] [CrossRef]
- Rajashekar, J.K.; Richard, J.; Beloor, J.; Prévost, J.; Anand, S.P.; Beaudoin-Bussières, G.; Shan, L.; Herndler-Brandstetter, D.; Gendron-Lepage, G.; Medjahed, H.; et al. Modulating HIV-1 envelope glycoprotein conformation to decrease the HIV-1 reservoir. Cell Host Microbe 2021, 29. [Google Scholar] [CrossRef]
- Wang, W.; Cong, Z.; Jiang, H.; Chen, T.; Jin, G.; Xiong, J.; Qin, C.; Wei, Q. Comparison of viral burden and disease progression in Chinese-origin rhesus macaques infected with common experimentally applied chimeric virus: SHIV-1157ipd3N4, SHIV-162P3, or SHIV-KB9. J. Med. Primatol. 2014, 43, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Whitney, J.B.; Luedemann, C.; Hraber, P.; Rao, S.S.; Mascola, J.R.; Nabel, G.J.; Letvin, N.L. T-cell vaccination reduces simian immunodeficiency virus levels in semen. J. Virol. 2009, 83, 10840–10843. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Fan, X.; Xie, Z.; Shen, T.; Shao, Y. Establishment of a real-time fluorescent quantitative PCR for the rapid detection of viral reservoir in the peripheral blood of SIV-infected rhesus monkeys. Chin. J. Microbiol. Immunol. (Chin.) 2016, 36, 304–309. [Google Scholar]
- Reeves, R.K.; Gillis, J.; Wong, F.E.; Yu, Y.; Connole, M.; Johnson, R.P. CD16- natural killer cells: Enrichment in mucosal and secondary lymphoid tissues and altered function during chronic SIV infection. Blood 2010, 115, 4439–4446. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhang, Z.; Wang, X.; Fu, J.L.; Yao, J.; Jiao, Y.; Chen, L.; Zhang, H.; Wei, J.; Jin, L.; et al. PD-1 up-regulation is correlated with HIV-specific memory CD8+ T-cell exhaustion in typical progressors but not in long-term nonprogressors. Blood 2007, 109, 4671–4678. [Google Scholar] [CrossRef]
- Jones, R.B.; Ndhlovu, L.C.; Barbour, J.D.; Sheth, P.M.; Jha, A.R.; Long, B.R.; Wong, J.C.; Satkunarajah, M.; Schweneker, M.; Chapman, J.M.; et al. Tim-3 expression defines a novel population of dysfunctional T cells with highly elevated frequencies in progressive HIV-1 infection. J. Exp. Med. 2008, 205, 2763–2779. [Google Scholar] [CrossRef]
- Fenwick, C.; Joo, V.; Jacquier, P.; Noto, A.; Banga, R.; Perreau, M.; Pantaleo, G. T-cell exhaustion in HIV infection. Immunol. Rev. 2019, 292, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Redondo, R.; Fryer, H.R.; Bedford, T.; Kim, E.Y.; Archer, J.; Pond, S.L.K.; Chung, Y.S.; Penugonda, S.; Chipman, J.; Fletcher, C.V.; et al. Persistent HIV-1 replication maintains the tissue reservoir during therapy. Nature 2016, 530, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaBonte, M.L.; McKay, P.F.; Letvin, N.L. Evidence of NK cell dysfunction in SIV-infected rhesus monkeys: Impairment of cytokine secretion and NKG2C/C2 expression. Eur. J. Immunol. 2006, 36, 2424–2433. [Google Scholar] [CrossRef]
- Masopust, D.; Soerens, A.G. Tissue-Resident T Cells and Other Resident Leukocytes. Annu Rev Immunol 2019, 37, 521–546. [Google Scholar] [CrossRef]
- Dogra, P.; Rancan, C.; Ma, W.; Toth, M.; Senda, T.; Carpenter, D.J.; Kubota, M.; Matsumoto, R.; Thapa, P.; Szabo, P.A.; et al. Tissue Determinants of Human NK Cell Development, Function, and Residence. Cell 2020, 180, 749–763.e713. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, E.; Malarkannan, S. Tissue-Resident NK Cells: Development, Maturation, and Clinical Relevance. Cancers 2020, 12, 1553. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Richard, J.; Lichtfuss, M.; Smith, A.B., 3rd; Park, J.; Courter, J.R.; Melillo, B.N.; Sodroski, J.G.; Kaufmann, D.E.; Finzi, A.; et al. Antibody-Dependent Cellular Cytotoxicity against Reactivated HIV-1-Infected Cells. J. Virol. 2016, 90, 2021–2030. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Wang, Y.; Ji, P.; Li, W.; Ying, T.; Huang, J.; Wang, C.; Wu, Y.; Wang, Y.; Chen, W.; et al. A defucosylated bispecific multivalent molecule exhibits broad HIV-1-neutralizing activity and enhanced antibody-dependent cellular cytotoxicity against reactivated HIV-1 latently infected cells. AIDS (Lond. Engl.) 2018, 32, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Abad-Fernandez, M.; Tuyishime, M.; Pollara, J.J.; Ferrari, G.; Soriano-Sarabia, N.; Margolis, D.M. Interleukin-15-Stimulated Natural Killer Cells Clear HIV-1-Infected Cells following Latency Reversal Ex Vivo. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.; Huot, N.; Micci, L.; Tharp, G.; King, C.; Rascle, P.; Shenvi, N.; Wang, H.; Galardi, C.; Upadhyay, A.A.; et al. IL-21 and IFNα therapy rescues terminally differentiated NK cells and limits SIV reservoir in ART-treated macaques. Nat. Commun. 2021, 12, 2866. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, G.D.; Ferrari, G.; Shen, X.; Alam, S.M.; Liao, H.X.; Pollara, J.; Bonsignori, M.; Moody, M.A.; Fong, Y.; Chen, X.; et al. Vaccine-induced plasma IgA specific for the C1 region of the HIV-1 envelope blocks binding and effector function of IgG. Proc. Natl. Acad. Sci. USA 2013, 110, 9019–9024. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal No. | PBMC | 2 PaLNs | 3 ALNs | Spleen | p Value | |
|---|---|---|---|---|---|---|
| CD4+T cells (CD3+CD4+CD8−) | ||||||
| DNA | G0101R | 440 | 441 | 1150 | 850 | 0.5206 |
| G0102R | 1554 | 827 | 13,666 | 4908 | ||
| G0104R | 3040 | 1295 | 861 | 1201 | ||
| G0105R | 2473 | 4873 | 7824 | 1214 | ||
| G0106R | 588 | 2536 | 1404 | 851 | ||
| RNA | G0101R | 310 | 287 | 455 | 660 | 0.1066 |
| G0102R | 73 | 505 | 2150 | 145 | ||
| G0104R | 100 | 242 | 124 | 81 | ||
| G0105R | 860 | 632 | 3237 | 1049 | ||
| G0106R | 81 | 463 | 507 | 313 | ||
| DNA/RNA | G0101R | 1.42 | 1.54 | 2.53 | 1.29 | 0.2096 |
| G0102R | 21.29 | 1.64 | 6.36 | 3.84 | ||
| G0104R | 30.4 | 5.35 | 6.94 | 14.83 | ||
| G0105R | 2.87 | 7.71 | 2.41 | 1.18 | ||
| G0106R | 7.26 | 5.48 | 2.75 | 2.72 | ||
| HLADR− CD4+ T cells (CD3+CD4+CD8−HLADR−) | ||||||
| DNA | G0101R | 3971 | 1100 | 3693 | 4550 | 0.5206 |
| G0102R | 33,338 | 2828 | 14,670 | 10,199 | ||
| G0104R | 6391 | 4474 | 927 | 1795 | ||
| G0105R | 2863 | 12791 | 9944 | 1689 | ||
| G0106R | 2594 | 6322 | 2185 | 1335 | ||
| RNA | G0101R | 163 | 72 | 257 | 323 | 0.7709 |
| G0102R | 463 | 216 | 2089 | 424 | ||
| G0104R | 178 | 194 | 112 | 122 | ||
| G0105R | 97 | 553 | 359 | 102 | ||
| G0106R | 24 | 254 | 146 | 68 | ||
| DNA/RNA | G0101R | 24.36 | 15.28 | 14.37 | 14.08 | 0.0055 ** |
| G0102R | 72 | 13.09 | 7.02 | 24.05 | ||
| G0104R | 35.9 | 23.06 | 8.27 | 14.71 | ||
| G0105R | 29.51 | 23.13 | 27.7 | 16.56 | ||
| G0106R | 108.08 | 24.89 | 14.96 | 19.63 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhu, L.; Yin, Y.; Fan, X.; Lv, L.; Zhang, Y.; Pan, Y.; Yan, Y.; Liang, H.; Xue, J.; et al. Associations between NK Cells in Different Immune Organs and Cellular SIV DNA and RNA in Regional HLADR− CD4+ T Cells in Chronically SIVmac239-Infected, Treatment-Naïve Rhesus Macaques. Viruses 2022, 14, 2513. https://doi.org/10.3390/v14112513
Li X, Zhu L, Yin Y, Fan X, Lv L, Zhang Y, Pan Y, Yan Y, Liang H, Xue J, et al. Associations between NK Cells in Different Immune Organs and Cellular SIV DNA and RNA in Regional HLADR− CD4+ T Cells in Chronically SIVmac239-Infected, Treatment-Naïve Rhesus Macaques. Viruses. 2022; 14(11):2513. https://doi.org/10.3390/v14112513
Chicago/Turabian StyleLi, Xinjie, Liyan Zhu, Yue Yin, Xueying Fan, Linting Lv, Yuqi Zhang, Yijin Pan, Yangxuanyu Yan, Hua Liang, Jing Xue, and et al. 2022. "Associations between NK Cells in Different Immune Organs and Cellular SIV DNA and RNA in Regional HLADR− CD4+ T Cells in Chronically SIVmac239-Infected, Treatment-Naïve Rhesus Macaques" Viruses 14, no. 11: 2513. https://doi.org/10.3390/v14112513