
Quercetin Inhibits Hsp70 Blocking of Bovine Viral Diarrhea Virus Infection and Replication in the Early Stage of Virus Infection
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Reagents
2.2. Toxicity of Quercetin on MDBK Cells
2.3. Measurement of Quercetin’s Antiviral Activity
2.4. Determining the Effects of Quercetin on Hsp70 Expression and BVDV Replication
2.5. Determining the Effects of Hsp70 Overexpression on BVDV Replication
2.6. Determining the Effects of Hsp70 SiRNA on BVDV Replication
2.7. Determining the Effects of Quercetin and Hsp70 SiRNA on the Expression of Antioxidant Genes in MDBK Cells
2.8. Determining the Effects of Quercetin and Hsp70 SiRNA on Intracellular ROS in MDBK Cells
2.9. Determining the Effects of Quercetin and Hsp70 SiRNA on Antioxidant Enzymes and MDA Indexes in MDBK Cells
2.10. Determining the Effects of Quercetin and Hsp70 SiRNA on the ERK Pathway
2.11. BVDV Animal Infection Model
2.12. In Vivo Virus Replication Assay
2.13. Analysis of the IL-2 and IFN-γ Concentration In Vivo
2.14. Analysis of Antioxidant Gene Expression In Vivo
2.15. Analysis of Oxidative Stress and Antioxidant Enzymes In Vivo
2.16. Histological Analysis
2.17. Statistical Analysis
3. Results
3.1. Quercetin Inhibits Hsp70 Expression and BVDV Replication
3.2. Hsp70 Positively Regulates the Production of BVDV
3.3. Quercetin and Hsp70 Reduce BVDV-Induced Oxidative Stress
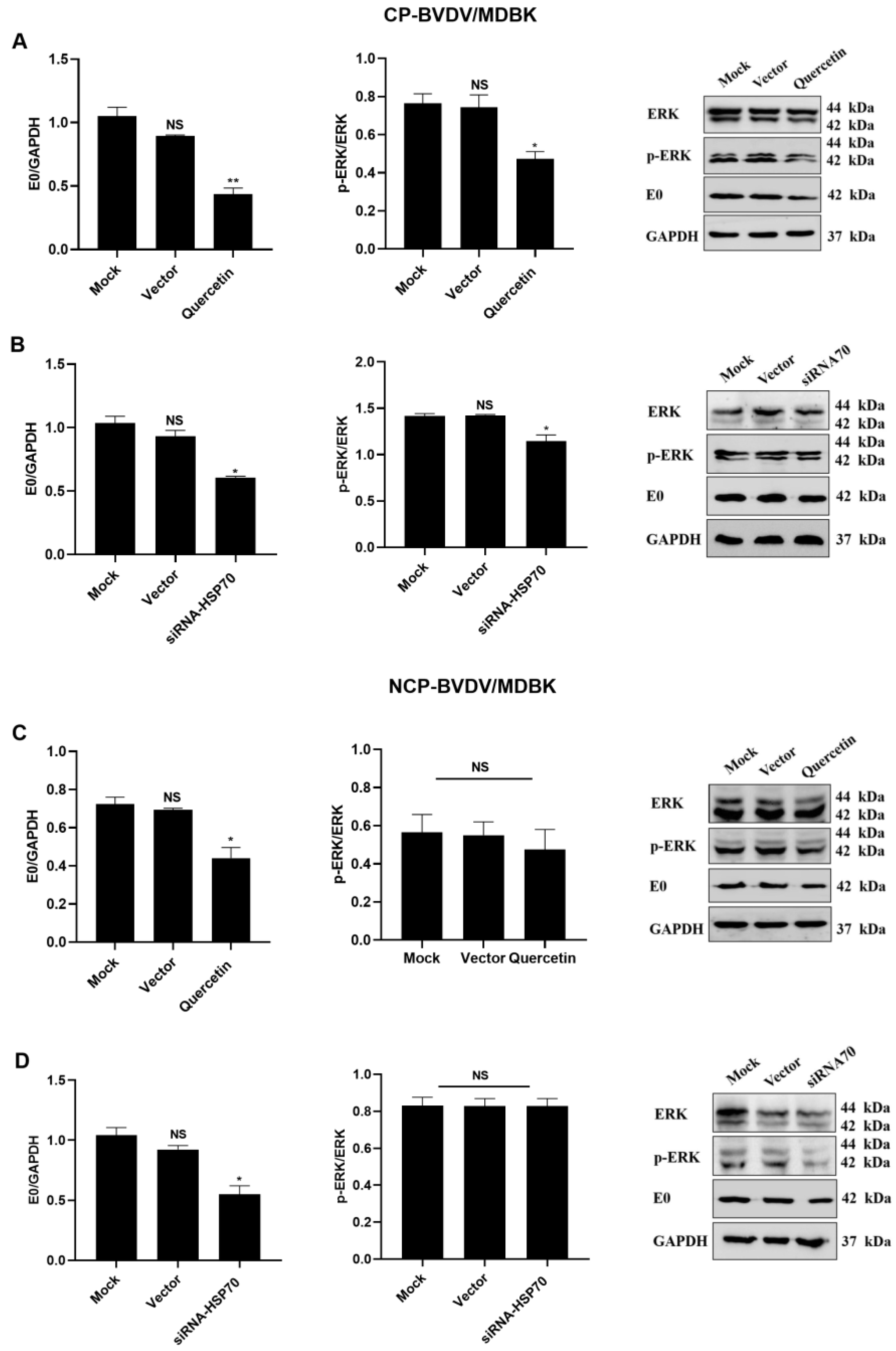
3.4. Quercetin and Hsp70 Affects ERK/P–ERK Pathway
3.5. Quercetin Inhibits Virus Replication in Mice
3.6. Quercetin Alleviates the Reduction of IFN-γ and IL-2 in the Serum of BVDV-Infected Mice
3.7. Quercetin Inhibits Oxidative Stress and Increases the Expression of Antioxidant Enzymes In Vivo
3.8. Quercetin Ameliorated BVDV-Induced Histopathological Changes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability statement
Conflicts of Interest
References
- Liu, Y.; Wu, C.; Chen, N.; Li, Y.; Fan, C.; Zhao, S.; Bai, T.; Zhao, Z.; Chen, J.; Su, S.; et al. PD-1 Blockade Restores the Proliferation of Peripheral Blood Lymphocyte and Inhibits Lymphocyte Apoptosis in a BALB/c Mouse Model of cp BVDV Acute Infection. Front. Immunol. 2021, 12, 727254. [Google Scholar] [CrossRef]
- Rajput, M.K.; Darweesh, M.F.; Park, K.; Braun, L.J.; Mwangi, W.; Young, A.J.; Chase, C.C. The effect of bovine viral diarrhea virus (BVDV) strains on bovine monocyte-derived dendritic cells (Mo-DC) phenotype and capacity to produce BVDV. J. Virol. 2014, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuca, T.; Passler, T.; Newcomer, B.W.; Neill, J.D.; Galik, P.K.; Riddell, K.P.; Zhang, Y.; Bayles, D.O.; Walz, P.H. Changes Introduced in the Open Reading Frame of Bovine Viral Diarrhea Virus During Serial Infection of Pregnant Swine. Front. Microbiol. 2020, 11, 1138. [Google Scholar] [CrossRef]
- Buckwold, V.E.; Beer, B.E.; Donis, R.O. Bovine viral diarrhea virus as a surrogate model of hepatitis C virus for the evaluation of antiviral agents. Antiviral. Res. 2003, 60, 1–15. [Google Scholar] [CrossRef]
- Newcomer, B.W.; Walz, P.H.; Givens, M.D. Potential applications for antiviral therapy and prophylaxis in bovine medicine. Anim. Health Res. Rev. 2014, 15, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Fernández, G.A.; Castro, E.F.; Rosas, R.A.; Fidalgo, D.M.; Adler, N.S.; Battini, L.; España de Marco, M.J.; Fabiani, M.; Bruno, A.M.; Bollini, M.; et al. Design and Optimization of Quinazoline Derivatives: New Non-nucleoside Inhibitors of Bovine Viral Diarrhea Virus. Front. Chem. 2020, 10, 590235. [Google Scholar] [CrossRef]
- Di, P.A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and its derivates as antiviral potentials: A comprehensive review. Phytother. Res. 2022, 36, 266–278. [Google Scholar] [CrossRef]
- Yang, J.X.; Maria, T.C.; Zhou, B.; Xiao, F.L.; Wang, M.; Mao, Y.J.; Li, Y. Quercetin improves immune function in Arbor Acre broilers through activation of NF-κB signaling pathway. Poult. Sci. 2020, 99, 906–913. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral Hepat. 2012, 19, e81–e88. [Google Scholar] [CrossRef]
- Gangwar, V.; Garg, A.; Lomore, K.; Korla, K.; Bhat, S.S.; Rao, R.P.; Rafiq, M.; Kumawath, R.; Uddagiri, B.V.; Kareenhalli, V.V. Immunomodulatory Effects of a Concoction of Natural Bioactive Compounds-Mechanistic Insights. Biomedicines 2021, 9, 1522. [Google Scholar] [CrossRef]
- Fanunza, E.; Iampietro, M.; Distinto, S.; Corona, A.; Quartu, M.; Maccioni, E.; Horvat, B.; Tramontano, E. Quercetin Blocks Ebola Virus Infection by Counteracting the VP24 Interferon-Inhibitory Function. Antimicrob. Agents Chemother. 2020, 64, e00530-20. [Google Scholar] [CrossRef] [PubMed]
- Bachar, S.C.; Mazumder, K.; Bachar, R.; Aktar, A.; Al Mahtab, M. A Review of Medicinal Plants with Antiviral Activity Available in Bangladesh and Mechanistic Insight Into Their Bioactive Metabolites on SARS-CoV-2, HIV and HBV. Front. Pharmacol. 2021, 12, 732891. [Google Scholar] [CrossRef] [PubMed]
- Rojas, Á.; Del Campo, J.A.; Clement, S.; Lemasson, M.; García-Valdecasas, M.; Gil-Gómez, A.; Ranchal, I.; Bartosch, B.; Bautista, J.D.; Rosenberg, A.R.; et al. Effect of Quercetin on Hepatitis C Virus Life Cycle: From Viral to Host Targets. Sci. Rep. 2016, 6, 31777. [Google Scholar] [CrossRef] [PubMed]
- Storniolo, A.; Raciti, M.; Cucina, A.; Bizzarri, M.; Di Renzo, L. Quercetin affects Hsp70/IRE1α mediated protection from death induced by endoplasmic reticulum stress. Oxid. Med. Cell. Longev. 2015, 2015, 645157. [Google Scholar] [CrossRef] [Green Version]
- Takashima, K.; Oshiumi, H.; Matsumoto, M.; Seya, T. DNAJB1/HSP40 Suppresses Melanoma Differentiation-Associated Gene 5-Mitochondrial Antiviral Signaling Protein Function in Conjunction with Hsp70. J. Innate Immun. 2018, 10, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Kregel, K.C. Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2022, 92, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Kang, K.; Ning, P.; Peng, Y.; Lin, Z.; Cui, H.; Cao, Z.; Wang, J.; Zhang, Y. Heat shock protein 70 is associated with CSFV NS5A protein and enhances viral RNA replication. Virology 2015, 482, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Taguwa, S.; Maringer, K.; Li, X.; Bernal-Rubio, D.; Rauch, J.N.; Gestwicki, J.E.; Andino, R.; Fernandez-Sesma, A.; Frydman, J. Defining Hsp70 Subnetworks in Dengue Virus Replication Reveals Key Vulnerability in Flavivirus Infection. Cell 2015, 163, 1108–1123. [Google Scholar] [CrossRef] [Green Version]
- Pujhari, S.; Brustolin, M.; Macias, V.M.; Nissly, R.H.; Nomura, M.; Kuchipudi, S.V.; Rasgon, J.L. Heat shock protein 70 (Hsp70) mediates Zika virus entry, replication, and egress from host cells. Emerg. Microbes Infect. 2019, 8, 8–16. [Google Scholar] [CrossRef]
- Schweizer, M.; Peterhans, E. Oxidative stress in cells infected with bovine viral diarrhoea virus: A crucial step in the induction of apoptosis. J. Gen. Virol. 1999, 80, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Yamane, D.; Zahoor, M.A.; Mohamed, Y.M.; Azab, W.; Kato, K.; Tohya, Y.; Akashi, H. Activation of extracellular signal-regulated kinase in MDBK cells infected with bovine viral diarrhea virus. Arch. Virol. 2009, 154, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kim, D.G.; Kim, S. ERK-dependent phosphorylation of the linker and substrate-binding domain of Hsp70 increases folding activity and cell proliferation. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wharton, W.; Moseley, P.; Shi, H. Heat shock protein 70 regulates cellular redox status by modulating glutathione-related enzyme activities. Cell Stress Chaperones 2007, 12, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xiao, S.; Liu, X.; Wang, L.; Ji, Q.; Mo, D.; Chen, Y. Inhibition of Hsp70 reduces porcine reproductive and respiratory syndrome virus replication in vitro. BMC Microbiol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.W.; Chen, Y.; Liu, W.; Chen, Y.J.; Fan, Z.M.; Liu, M.; Li, L.J. Inhibition of JAK2/STAT3 Signaling Pathway Suppresses Proliferation of Burkitt’s Lymphoma Raji Cells via Cell Cycle Progression, Apoptosis, and Oxidative Stress by Modulating Hsp70. Med. Sci. Monit. 2018, 24, 6255–6263. [Google Scholar] [CrossRef]
- Senthilvel, P.; Lavanya, P.; Kumar, K.M.; Swetha, R.; Anitha, P.; Bag, S.; Sarveswari, S.; Vijayakumar, V.; Ramaiah, S.; Anbarasu, A. Flavonoid from Carica papaya inhibits NS2B-NS3 protease and prevents Dengue 2 viral assembly. Bioinformation 2013, 9, 889–895. [Google Scholar] [CrossRef]
- De Sousa, L.R.; Wu, H.; Nebo, L.; Fernandes, J.B.; Da Silva, M.F.; Kiefer, W.; Kanitz, M.; Bodem, J.; Diederich, W.E.; Schirmeister, T.; et al. Flavonoids as noncompetitive inhibitors of Dengue virus NS2B-NS3 protease: Inhibition kinetics and docking studies. Bioorg. Med. Chem. 2015, 23, 466–470. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral activity of baicalein and quercetin against the Japanese encephalitis virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, B.; Boucomont-Chapeaublanc, E.; Peyrefitte, C.N.; Belghazi, M.; Fusaï, T.; Rogier, C.; Tolou, H.J.; Almeras, L. Identification of cellular proteome modifications in response to West Nile virus infection. Mol. Cell. Proteom. 2009, 8, 1623–1637. [Google Scholar] [CrossRef] [Green Version]
- Bolhassani, A.; Agi, E. Heat shock proteins in infection. Clin. Chim. Acta 2019, 498, 90–100. [Google Scholar] [CrossRef] [PubMed]
- La Polla, R.; Testard, M.C.; Goumaidi, A.; Chapot, E.; Legras-Lachuer, C.; De Saint-Vis, B. Identification of differentially expressed gene pathways between cytopathogenic and non-cytopathogenic BVDV-1 strains by analysis of the transcriptome of infected primary bovine cells. Virology 2021, 567, 34–46. [Google Scholar] [CrossRef]
- Shi, Z.; Sun, J.; Guo, H.; Tu, C. Genomic expression profiling of peripheral blood leukocytes of pigs infected with highly virulent classical swine fever virus strain Shimen. J. Gen. Virol. 2009, 90, 1670–1680. [Google Scholar] [CrossRef]
- Shaul, Y.D.; Seger, R. The MEK/ERK cascade: From signaling specificity to diverse functions. Biochim. Biophys. Acta 2016, 1773, 1213–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo, M.; Olivier, M. Hydrogen peroxide induces murine macrophage chemokine gene transcription via extracellular signal-regulated kinase-and cyclic adenosine 5′-monophosphate (cAMP)-dependent pathways: Involvement of NF-kappa B, activator protein 1, and cAMP response element binding protein. J. Immunol. 2002, 169, 7026–7038. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Qi, S.; Gui, L.; Shen, L.; Feng, Z. Daphnetin protects oxidative stress-induced neuronal apoptosis via regulation of MAPK signaling and Hsp70 expression. Oncol. Lett. 2016, 12, 1959–1964. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′–3′) | Product Length (bp) |
|---|---|---|
| Nrf2 | F: ACCCAGTCCAACCTTTGTCGT | 143 |
| R: GCGGCTTGAATGTTTGTCTTT | ||
| NQO1 | F: CGGCTCCATGTACTCTCTGC | 183 |
| R: TCCAGGCGTTTCTTCCATCC | ||
| HO-1 | F: CAAGCGCTATGTTCAGCGAC | 198 |
| R: TTGGTGGCACTGGCGATATT | ||
| β-actin | F: CACCGCAAATGCTTCTAGGC | 186 |
| R: TGTCACCTTCACCGTTCCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, N.; Liu, Y.; Bai, T.; Chen, J.; Zhao, Z.; Li, J.; Shao, B.; Zhang, Z.; Zhou, Y.; Wang, X.; et al. Quercetin Inhibits Hsp70 Blocking of Bovine Viral Diarrhea Virus Infection and Replication in the Early Stage of Virus Infection. Viruses 2022, 14, 2365. https://doi.org/10.3390/v14112365
Chen N, Liu Y, Bai T, Chen J, Zhao Z, Li J, Shao B, Zhang Z, Zhou Y, Wang X, et al. Quercetin Inhibits Hsp70 Blocking of Bovine Viral Diarrhea Virus Infection and Replication in the Early Stage of Virus Infection. Viruses. 2022; 14(11):2365. https://doi.org/10.3390/v14112365
Chicago/Turabian StyleChen, Nannan, Yu Liu, Tongtong Bai, Jinwei Chen, Zhibo Zhao, Jing Li, Baihui Shao, Zecai Zhang, Yulong Zhou, Xue Wang, and et al. 2022. "Quercetin Inhibits Hsp70 Blocking of Bovine Viral Diarrhea Virus Infection and Replication in the Early Stage of Virus Infection" Viruses 14, no. 11: 2365. https://doi.org/10.3390/v14112365