
Cytokine Profiling of Amniotic Fluid from Congenital Cytomegalovirus Infection
 , ,
, ,
Abstract
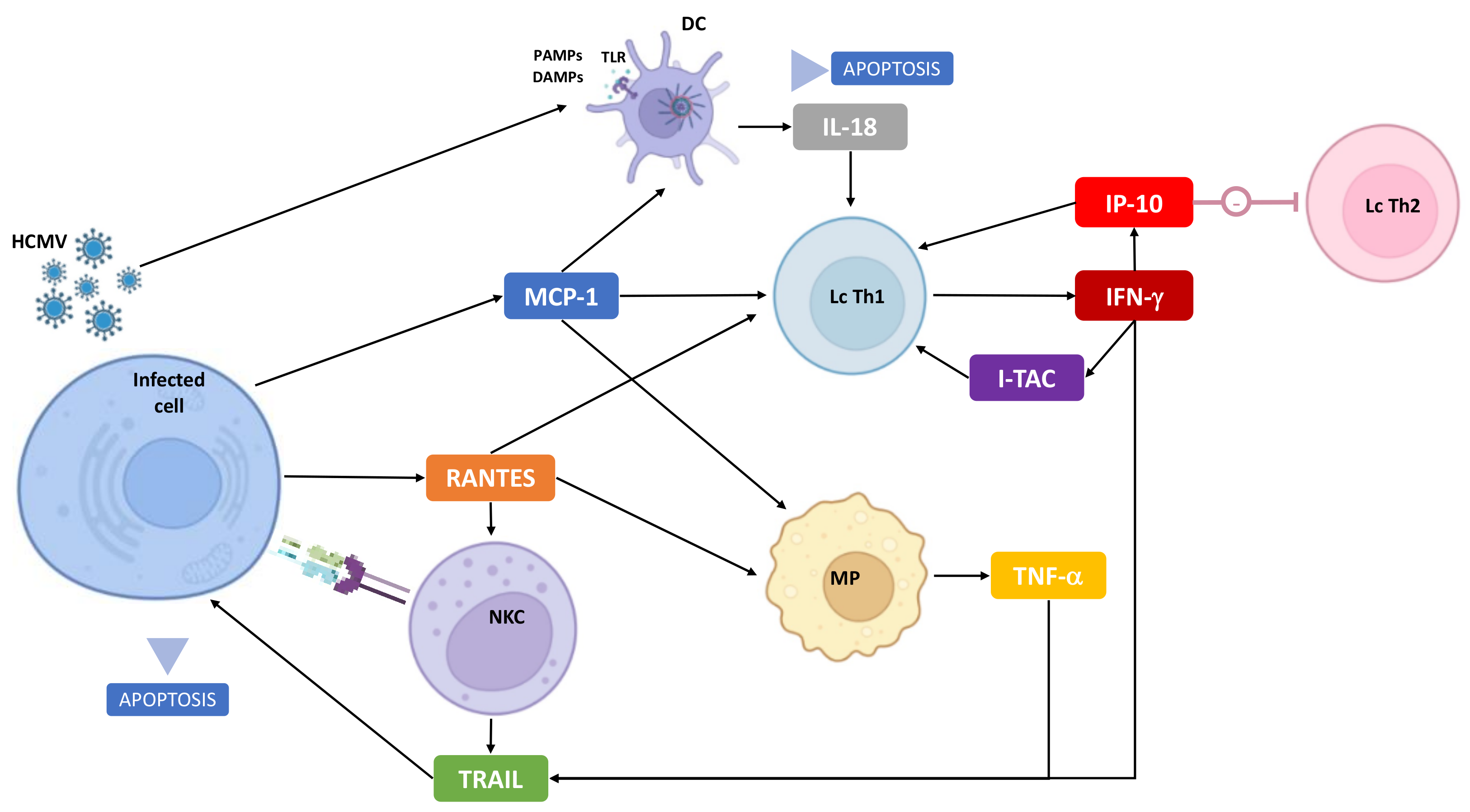
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Diagnosis of MPI, Fetal, and Neonatal Infection
2.3. Clinical Classification of Infected Newborns
2.4. Preparation of Extracellular Vesicle Fractions
2.5. Cytokines Concentrations Measurement
2.6. Statistical Analysis
3. Results
3.1. Description of the Cohort
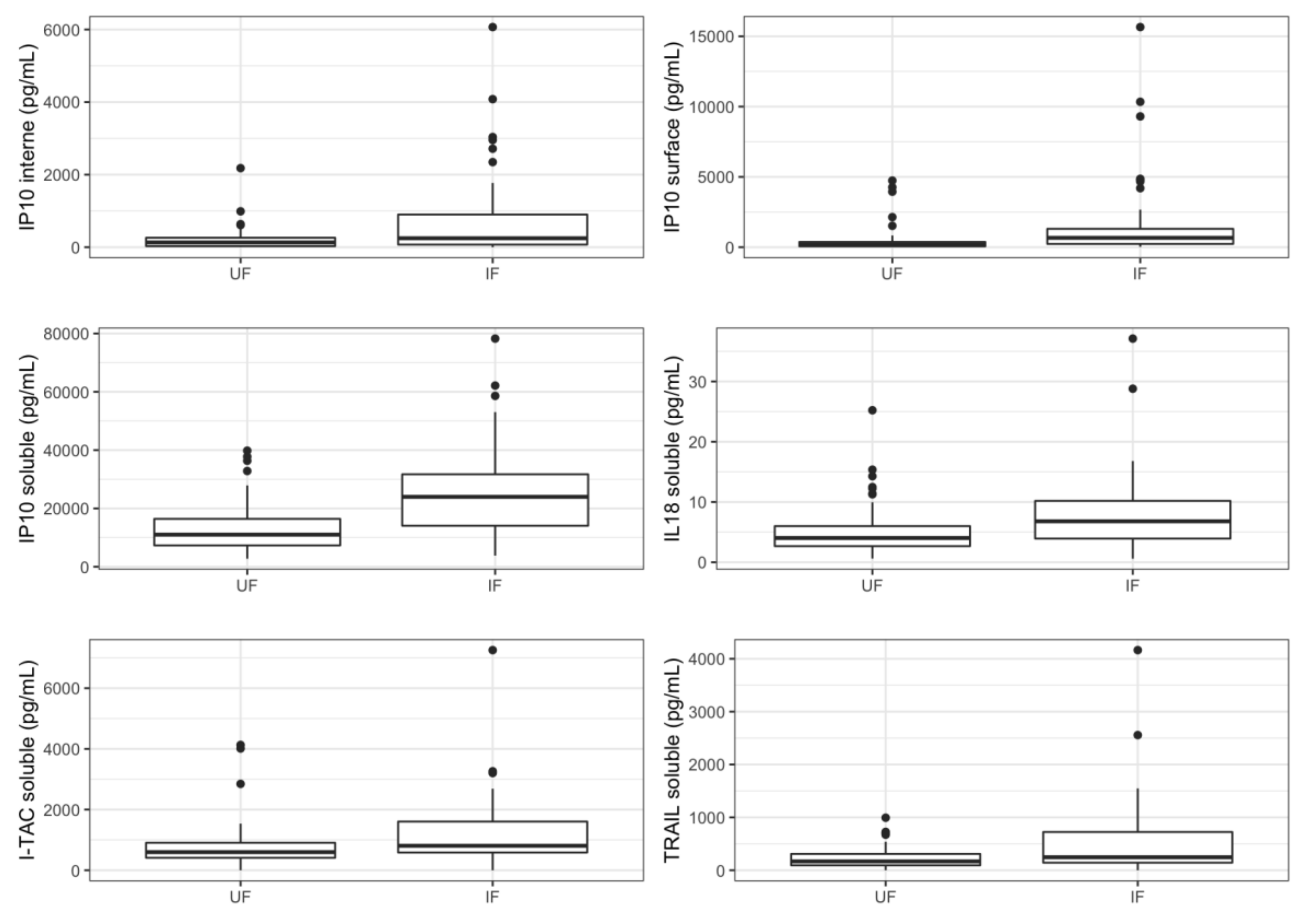
3.2. Cytokines Profiling in Amniotic Fluid of Infected Fetuses
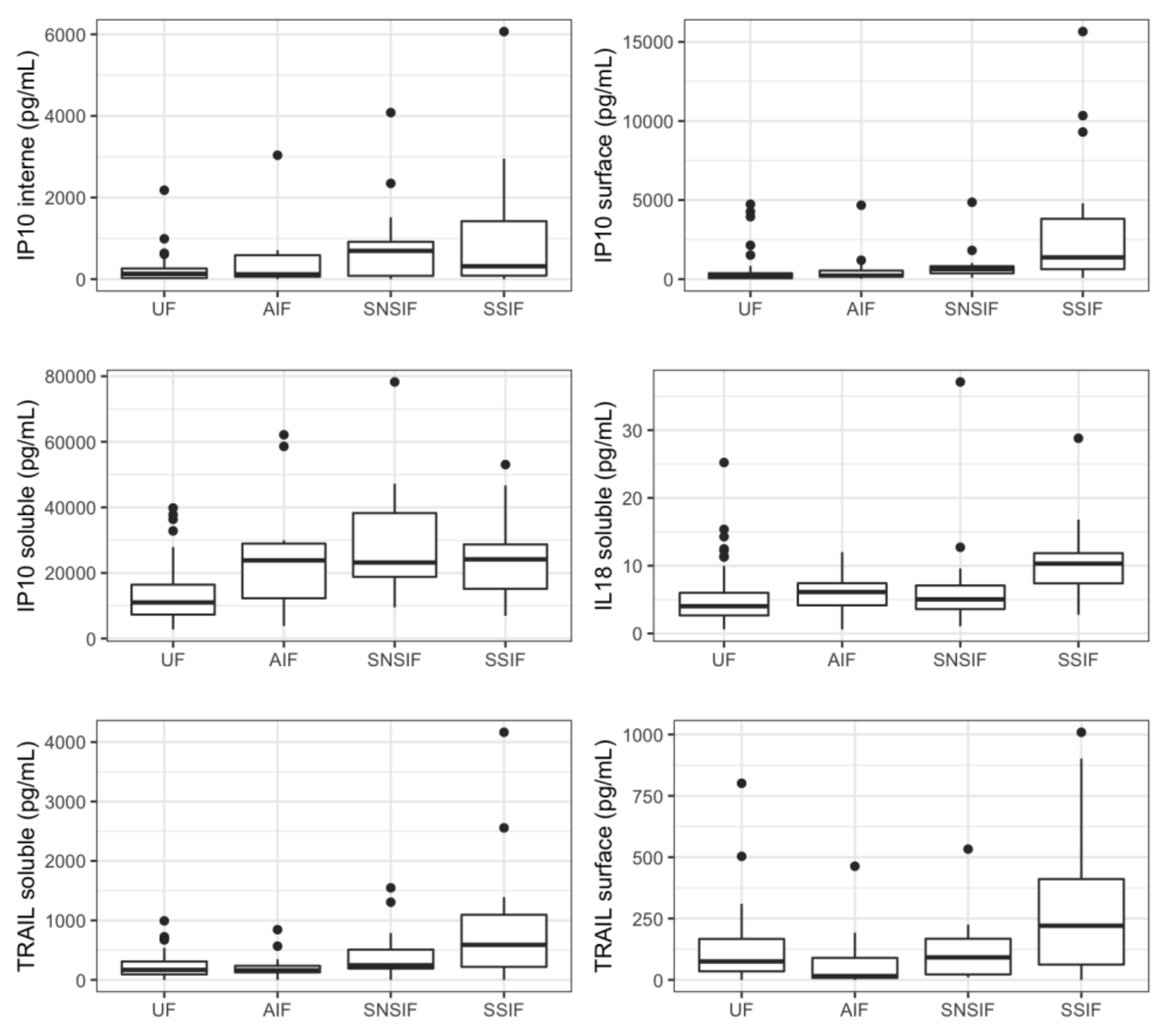
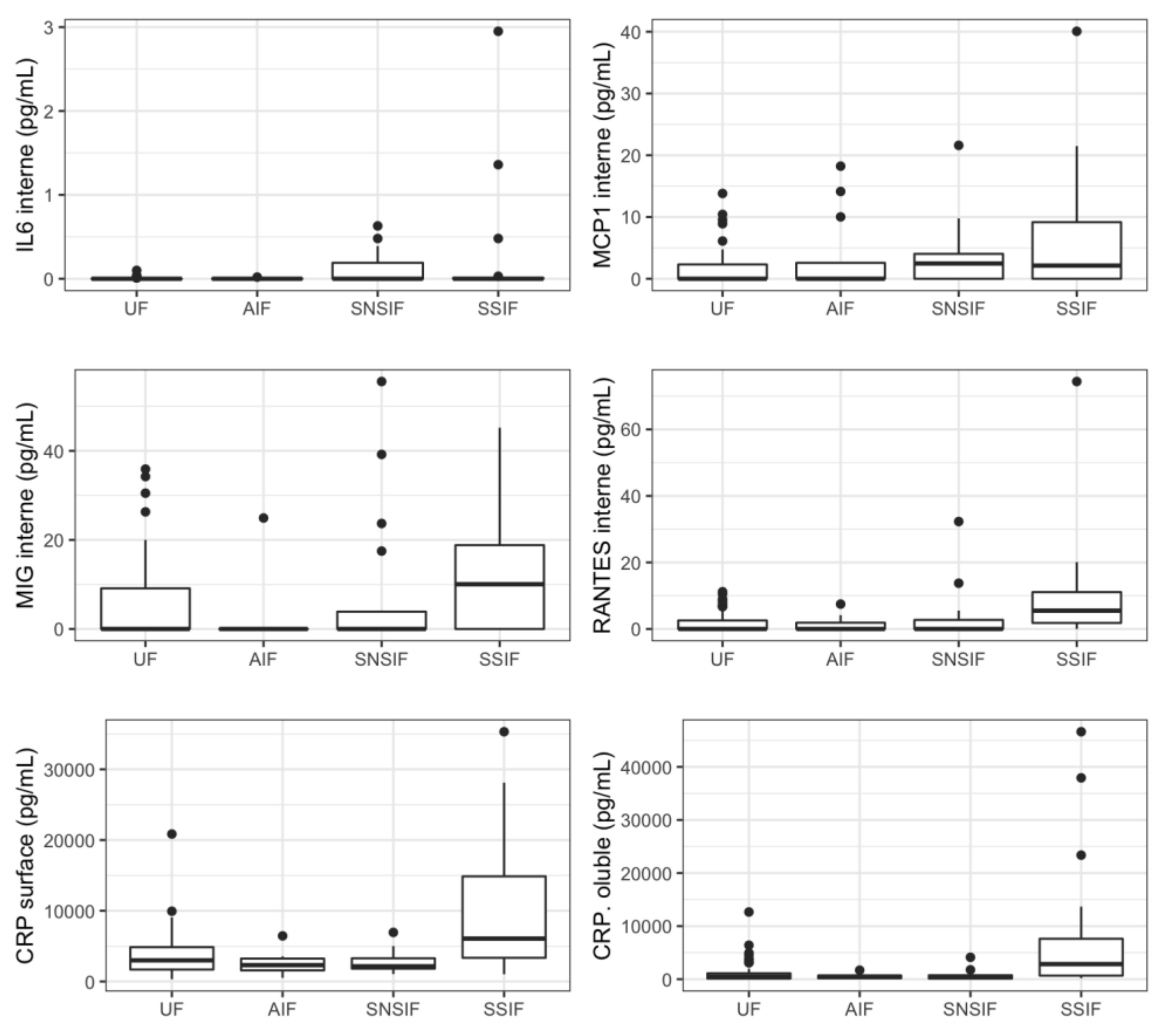
3.3. Cytokines Profiling in Amniotic Fluid of Symptomatic Newborns
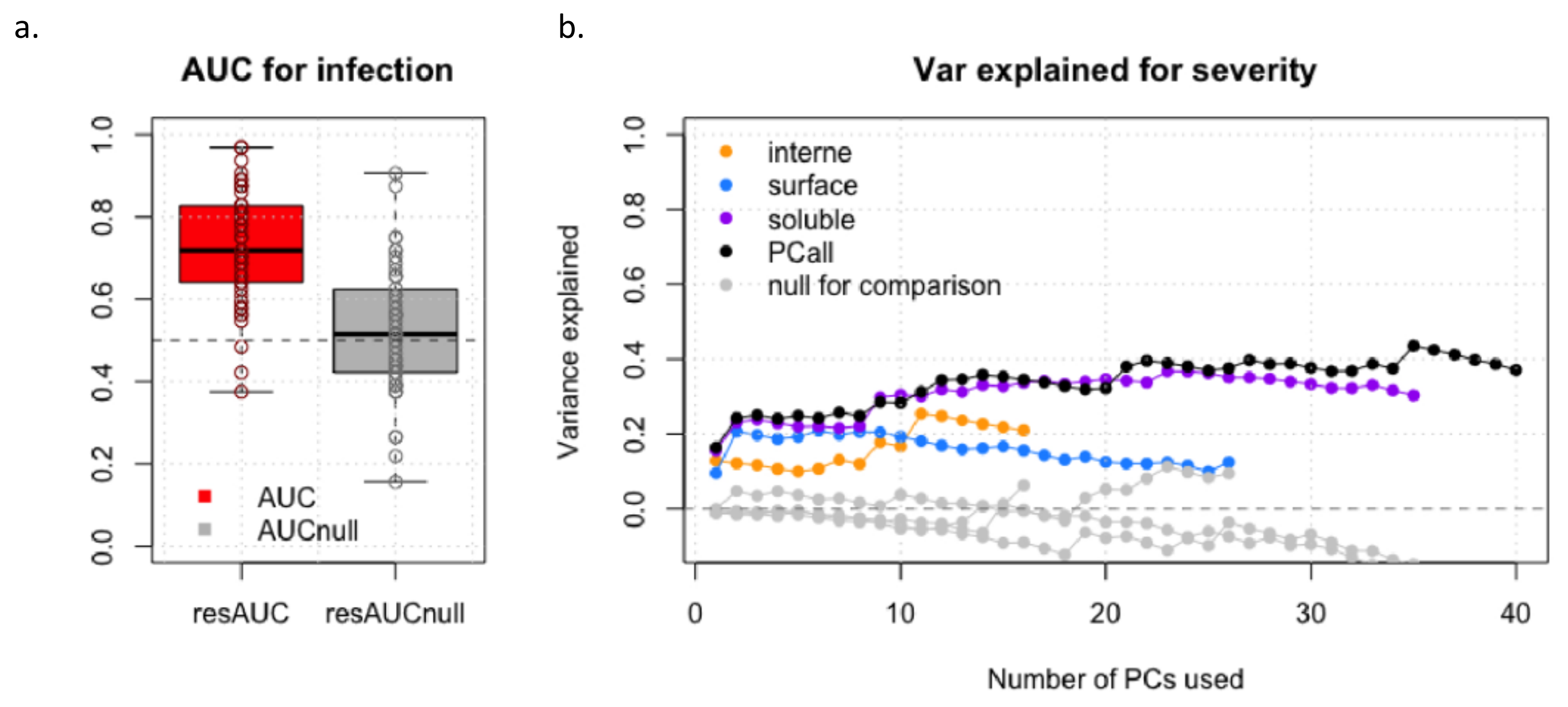
3.4. Prediction Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Cytokines Studied
Appendix B. Neonatal Assessment of Infected Fetuses
References
- Kenneson, A.; Cannon, M.J. Review and Meta-Analysis of the Epidemiology of Congenital Cytomegalovirus (CMV) Infection. Rev. Med. Virol. 2007, 17, 253–276. [Google Scholar] [CrossRef] [PubMed]
- Leruez-Ville, M.; Foulon, I.; Pass, R.; Ville, Y. Cytomegalovirus Infection during Pregnancy: State of the Science. Am. J. Obstet. Gynecol. 2020, 223, 330–349. [Google Scholar] [CrossRef] [PubMed]
- Smithers-Sheedy, H.; Raynes-Greenow, C.; Badawi, N.; Fernandez, M.A.; Kesson, A.; McIntyre, S.; Leung, K.-C.; Jones, C.A. Congenital Cytomegalovirus among Children with Cerebral Palsy. J. Pediatr. 2017, 181, 267–271.e1. [Google Scholar] [CrossRef] [PubMed]
- Korver, A.M.H.; de Vries, J.J.C.; Konings, S.; de Jong, J.W.; Dekker, F.W.; Vossen, A.C.T.M.; Frijns, J.H.M.; Oudesluys-Murphy, A.M.; DECIBEL collaborative study group. DECIBEL Study: Congenital Cytomegalovirus Infection in Young Children with Permanent Bilateral Hearing Impairment in the Netherlands. J. Clin. Virol. 2009, 46 (Suppl. 4), S27–S31. [Google Scholar] [CrossRef] [PubMed]
- Goderis, J.; De Leenheer, E.; Smets, K.; Van Hoecke, H.; Keymeulen, A.; Dhooge, I. Hearing Loss and Congenital CMV Infection: A Systematic Review. Pediatrics 2014, 134, 972–982. [Google Scholar] [CrossRef]
- Avettand-Fenoël, V.; Marlin, S.; Vauloup-Fellous, C.; Loundon, N.; François, M.; Couloigner, V.; Rouillon, I.; Drouin-Garraud, V.; Laccourreye, L.; Denoyelle, F.; et al. Congenital Cytomegalovirus Is the Second Most Frequent Cause of Bilateral Hearing Loss in Young French Children. J. Pediatr. 2013, 162, 593–599. [Google Scholar] [CrossRef]
- Nance, W.E.; Lim, B.G.; Dodson, K.M. Importance of Congenital Cytomegalovirus Infections as a Cause for Pre-Lingual Hearing Loss. J. Clin. Virol. 2006, 35, 221–225. [Google Scholar] [CrossRef]
- Faure-Bardon, V.; Millischer, A.-E.; Deloison, B.; Sonigo, P.; Grévent, D.; Salomon, L.; Stirnemann, J.; Nicloux, M.; Magny, J.-F.; Leruez-Ville, M.; et al. Refining the Prognosis of Fetuses Infected with Cytomegalovirus in the First Trimester of Pregnancy by Serial Prenatal Assessment: A Single-Centre Retrospective Study. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 355–362. [Google Scholar] [CrossRef]
- Rawlinson, W.D.; Boppana, S.B.; Fowler, K.B.; Kimberlin, D.W.; Lazzarotto, T.; Alain, S.; Daly, K.; Doutré, S.; Gibson, L.; Giles, M.L.; et al. Congenital Cytomegalovirus Infection in Pregnancy and the Neonate: Consensus Recommendations for Prevention, Diagnosis, and Therapy. Lancet Infect. Dis. 2017, 17, e177–e188. [Google Scholar] [CrossRef]
- Dollard, S.C.; Grosse, S.D.; Ross, D.S. New Estimates of the Prevalence of Neurological and Sensory Sequelae and Mortality Associated with Congenital Cytomegalovirus Infection. Rev. Med. Virol. 2007, 17, 355–363. [Google Scholar] [CrossRef]
- Goderis, J.; Keymeulen, A.; Smets, K.; Van Hoecke, H.; De Leenheer, E.; Boudewyns, A.; Desloovere, C.; Kuhweide, R.; Muylle, M.; Royackers, L.; et al. Hearing in Children with Congenital Cytomegalovirus Infection: Results of a Longitudinal Study. J. Pediatr. 2016, 172, 110–115.e2. [Google Scholar] [CrossRef] [PubMed]
- Foulon, I.; Naessens, A.; Foulon, W.; Casteels, A.; Gordts, F. A 10-Year Prospective Study of Sensorineural Hearing Loss in Children with Congenital Cytomegalovirus Infection. J. Pediatr. 2008, 153, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Foulon, I.; De Brucker, Y.; Buyl, R.; Lichtert, E.; Verbruggen, K.; Piérard, D.; Camfferman, F.A.; Gucciardo, L.; Gordts, F. Hearing Loss With Congenital Cytomegalovirus Infection. Pediatrics 2019, 144, e20183095. [Google Scholar] [CrossRef] [PubMed]
- Grosse, S.D.; Ross, D.S.; Dollard, S.C. Congenital Cytomegalovirus (CMV) Infection as a Cause of Permanent Bilateral Hearing Loss: A Quantitative Assessment. J. Clin. Virol. 2008, 41, 57–62. [Google Scholar] [CrossRef]
- Fowler, K.B.; McCollister, F.P.; Dahle, A.J.; Boppana, S.; Britt, W.J.; Pass, R.F. Progressive and Fluctuating Sensorineural Hearing Loss in Children with Asymptomatic Congenital Cytomegalovirus Infection. J. Pediatr. 1997, 130, 624–630. [Google Scholar] [CrossRef]
- Dahle, A.J.; Fowler, K.B.; Wright, J.D.; Boppana, S.B.; Britt, W.J.; Pass, R.F. Longitudinal Investigation of Hearing Disorders in Children with Congenital Cytomegalovirus. J. Am. Acad. Audiol. 2000, 11, 283–290. [Google Scholar] [CrossRef]
- Ross, S.A.; Fowler, K.B.; Ashrith, G.; Stagno, S.; Britt, W.J.; Pass, R.F.; Boppana, S.B. Hearing Loss in Children with Congenital Cytomegalovirus Infection Born to Mothers with Preexisting Immunity. J. Pediatr. 2006, 148, 332–336. [Google Scholar] [CrossRef]
- Leruez-Ville, M.; Magny, J.-F.; Couderc, S.; Pichon, C.; Parodi, M.; Bussières, L.; Guilleminot, T.; Ghout, I.; Ville, Y. Risk Factors for Congenital Cytomegalovirus Infection Following Primary and Nonprimary Maternal Infection: A Prospective Neonatal Screening Study Using Polymerase Chain Reaction in Saliva. Clin. Infect. Dis. 2017, 65, 398–404. [Google Scholar] [CrossRef]
- Mussi-Pinhata, M.M.; Yamamoto, A.Y.; Brito, R.M.M.; de Lima Isaac, M.; de Carvalhoe Oliveira, P.F.; Boppana, S.; Britt, W.J. Birth Prevalence and Natural History of Congenital Cytomegalovirus Infection in a Highly Seroimmune Population. Clin. Infect. Dis. 2009, 49, 522–528. [Google Scholar] [CrossRef]
- Benoist, G.; Salomon, L.; Jacquemard, F.; Daffos, F.; Ville, Y. The Prognostic Value of Ultrasound Abnormalities and Biological Parameters in Blood of Fetuses Infected with Cytomegalovirus. BJOG Int. J. Obstet. Gynaecol. 2008, 115, 823–829. [Google Scholar] [CrossRef]
- Cannie, M.M.; Devlieger, R.; Leyder, M.; Claus, F.; Leus, A.; De Catte, L.; Cossey, V.; Foulon, I.; Van der valk, E.; Foulon, W.; et al. Congenital Cytomegalovirus Infection: Contribution and Best Timing of Prenatal MR Imaging. Eur. Radiol. 2016, 26, 3760–3769. [Google Scholar] [CrossRef]
- Picone, O.; Simon, I.; Benachi, A.; Brunelle, F.; Sonigo, P. Comparison between Ultrasound and Magnetic Resonance Imaging in Assessment of Fetal Cytomegalovirus Infection. Prenat. Diagn. 2008, 28, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Lipitz, S.; Hoffmann, C.; Feldman, B.; Tepperberg-Dikawa, M.; Schiff, E.; Weisz, B. Value of Prenatal Ultrasound and Magnetic Resonance Imaging in Assessment of Congenital Primary Cytomegalovirus Infection. Ultrasound Obstet. Gynecol. 2010, 36, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Leruez-Ville, M.; Stirnemann, J.; Sellier, Y.; Guilleminot, T.; Dejean, A.; Magny, J.-F.; Couderc, S.; Jacquemard, F.; Ville, Y. Feasibility of Predicting the Outcome of Fetal Infection with Cytomegalovirus at the Time of Prenatal Diagnosis. Am. J. Obstet. Gynecol. 2016, 215, 342.e1–342.e9. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Simonazzi, G.; Puccetti, C.; Lanari, M.; Farina, A.; Lazzarotto, T.; Rizzo, N. Ultrasound Prediction of Symptomatic Congenital Cytomegalovirus Infection. Am. J. Obstet. Gynecol. 2008, 198, 380.e1–380.e7. [Google Scholar] [CrossRef] [PubMed]
- Leyder, M.; Vorsselmans, A.; Done, E.; Berkel, K.V.; Faron, G.; Foulon, I.; Naessens, A.; Jansen, A.; Foulon, W.; Gucciardo, L. Primary Maternal Cytomegalovirus Infections: Accuracy of Fetal Ultrasound for Predicting Sequelae in Offspring. Am. J. Obstet. Gynecol. 2016, 215, 638.e1–638.e8. [Google Scholar] [CrossRef]
- Fabbri, E.; Revello, M.G.; Furione, M.; Zavattoni, M.; Lilleri, D.; Tassis, B.; Quarenghi, A.; Rustico, M.; Nicolini, U.; Ferrazzi, E.; et al. Prognostic Markers of Symptomatic Congenital Human Cytomegalovirus Infection in Fetal Blood. BJOG Int. J. Obstet. Gynaecol. 2011, 118, 448–456. [Google Scholar] [CrossRef]
- Griffiths, P.; Reeves, M. Pathogenesis of Human Cytomegalovirus in the Immunocompromised Host. Nat. Rev. Microbiol. 2021, 19, 759–773. [Google Scholar] [CrossRef]
- Faure-Bardon, V.; Magny, J.-F.; Parodi, M.; Couderc, S.; Garcia, P.; Maillotte, A.-M.; Benard, M.; Pinquier, D.; Astruc, D.; Patural, H.; et al. Sequelae of Congenital Cytomegalovirus Following Maternal Primary Infections Are Limited to Those Acquired in the First Trimester of Pregnancy. Clin. Infect. Dis. 2019, 69, 1526–1532. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, G.; Romero, R.; Rice, G.E.; Fitzgerald, W.; Pacora, P.; Gomez-Lopez, N.; Kavdia, M.; Tarca, A.L.; Margolis, L. Compartmentalized Profiling of Amniotic Fluid Cytokines in Women with Preterm Labor. PLoS ONE 2020, 15, e0227881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, R.; Grivel, J.-C.; Tarca, A.L.; Chaemsaithong, P.; Xu, Z.; Fitzgerald, W.; Hassan, S.S.; Chaiworapongsa, T.; Margolis, L. Evidence of Perturbations of the Cytokine Network in Preterm Labor. Am. J. Obstet. Gynecol. 2015, 213, 836.e1–836.e18. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, W.; Freeman, M.L.; Lederman, M.M.; Vasilieva, E.; Romero, R.; Margolis, L. A System of Cytokines Encapsulated in ExtraCellular Vesicles. Sci. Rep. 2018, 8, 8973. [Google Scholar] [CrossRef] [PubMed]
- Schleiss, M.R. Cytomegalovirus in the Neonate: Immune Correlates of Infection and Protection. Clin. Dev. Immunol. 2013, 2013, 501801. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.M.; Chow, S.S.W.; Craig, M.E.; Pang, C.N.I.; Hall, B.; Wilkins, M.R.; Jones, C.A.; Lloyd, A.R.; Rawlinson, W.D. Cytomegalovirus Infection during Pregnancy with Maternofetal Transmission Induces a Proinflammatory Cytokine Bias in Placenta and Amniotic Fluid. J. Infect. Dis. 2012, 205, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Tetta, C.; Ghigo, E.; Silengo, L.; Deregibus, M.C.; Camussi, G. Extracellular Vesicles as an Emerging Mechanism of Cell-to-Cell Communication. Endocrine 2013, 44, 11–19. [Google Scholar] [CrossRef]
- Keller, S.; Sanderson, M.P.; Stoeck, A.; Altevogt, P. Exosomes: From Biogenesis and Secretion to Biological Function. Immunol. Lett. 2006, 107, 102–108. [Google Scholar] [CrossRef]
- Costa, A.; Quarto, R.; Bollini, S. Small Extracellular Vesicles from Human Amniotic Fluid Samples as Promising Theranostics. Int. J. Mol. Sci. 2022, 23, 590. [Google Scholar] [CrossRef]
- Sheller-Miller, S.; Menon, R. Chapter Ten—Isolation and Characterization of Human Amniotic Fluid-Derived Exosomes. In Methods in Enzymology; Extracellular Vesicles; Spada, S., Galluzzi, L., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 645, pp. 181–194. [Google Scholar]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, Y.; Han, S.; Zhang, W.; Zhou, Q.; Guan, H.; Liu, J.; Shi, J.; Su, L.; Hu, D. Exosomes Derived from Human Amniotic Epithelial Cells Accelerate Wound Healing and Inhibit Scar Formation. J. Mol. Histol. 2017, 48, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Farhadihosseinabadi, B.; Farahani, M.; Tayebi, T.; Jafari, A.; Biniazan, F.; Modaresifar, K.; Moravvej, H.; Bahrami, S.; Redl, H.; Tayebi, L.; et al. Amniotic Membrane and Its Epithelial and Mesenchymal Stem Cells as an Appropriate Source for Skin Tissue Engineering and Regenerative Medicine. Artif. Cells Nanomed. Biotechnol. 2018, 46, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Lau, S.N.; Leaw, B.; Nguyen, H.P.T.; Salamonsen, L.A.; Saad, M.I.; Chan, S.T.; Zhu, D.; Krause, M.; Kim, C.; et al. Amnion Epithelial Cell-Derived Exosomes Restrict Lung Injury and Enhance Endogenous Lung Repair. Stem Cells Transl. Med. 2018, 7, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Sheller, S.; Papaconstantinou, J.; Urrabaz-Garza, R.; Richardson, L.; Saade, G.; Salomon, C.; Menon, R. Amnion-Epithelial-Cell-Derived Exosomes Demonstrate Physiologic State of Cell under Oxidative Stress. PLoS ONE 2016, 11, e0157614. [Google Scholar] [CrossRef]
- Keller, S.; Rupp, C.; Stoeck, A.; Runz, S.; Fogel, M.; Lugert, S.; Hager, H.-D.; Abdel-Bakky, M.S.; Gutwein, P.; Altevogt, P. CD24 Is a Marker of Exosomes Secreted into Urine and Amniotic Fluid. Kidney Int. 2007, 72, 1095–1102. [Google Scholar] [CrossRef]
- Asea, A.; Jean-Pierre, C.; Kaur, P.; Rao, P.; Linhares, I.M.; Skupski, D.; Witkin, S.S. Heat Shock Protein-Containing Exosomes in Mid-Trimester Amniotic Fluids. J. Reprod. Immunol. 2008, 79, 12–17. [Google Scholar] [CrossRef]
- Mellows, B.; Mitchell, R.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.-T.; Denecke, B.; Musante, L.; Ramachandra, D.L.; Debacq-Chainiaux, F.; et al. Protein and Molecular Characterization of a Clinically Compliant Amniotic Fluid Stem Cell-Derived Extracellular Vesicle Fraction Capable of Accelerating Muscle Regeneration Through Enhancement of Angiogenesis. Stem Cells Dev. 2017, 26, 1316–1333. [Google Scholar] [CrossRef]
- Bellio, M.A.; Young, K.C.; Milberg, J.; Santos, I.; Abdullah, Z.; Stewart, D.; Arango, A.; Chen, P.; Huang, J.; Williams, K.; et al. Amniotic Fluid-Derived Extracellular Vesicles: Characterization and Therapeutic Efficacy in an Experimental Model of Bronchopulmonary Dysplasia. Cytotherapy 2021, 23, 1097–1107. [Google Scholar] [CrossRef]
- Tavanasefat, H.; Li, F.; Koyano, K.; Gourtani, B.K.; Marty, V.; Mulpuri, Y.; Lee, S.H.; Shin, K.-H.; Wong, D.T.W.; Xiao, X.; et al. Molecular Consequences of Fetal Alcohol Exposure on Amniotic Exosomal MiRNAs with Functional Implications for Stem Cell Potency and Differentiation. PLoS ONE 2020, 15, e0242276. [Google Scholar] [CrossRef]
- Dixon, C.L.; Sheller-Miller, S.; Saade, G.R.; Fortunato, S.J.; Lai, A.; Palma, C.; Guanzon, D.; Salomon, C.; Menon, R. Amniotic Fluid Exosome Proteomic Profile Exhibits Unique Pathways of Term and Preterm Labor. Endocrinology 2018, 159, 2229–2240. [Google Scholar] [CrossRef] [Green Version]
- Fabietti, I.; Nardi, T.; Favero, C.; Dioni, L.; Cantone, L.; Pergoli, L.; Hoxha, M.; Pinatel, E.; Mosca, F.; Bollati, V.; et al. Extracellular Vesicles and Their MiRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention. Cells 2021, 10, 1493. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Humphreys, I.R. Cytokine-Mediated Induction and Regulation of Tissue Damage During Cytomegalovirus Infection. Front. Immunol. 2019, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Pitti, R.M.; Marsters, S.A.; Ruppert, S.; Donahue, C.J.; Moore, A.; Ashkenazi, A. Induction of Apoptosis by Apo-2 Ligand, a New Member of the Tumor Necrosis Factor Cytokine Family. J. Biol. Chem. 1996, 271, 12687–12690. [Google Scholar] [CrossRef]
- Sheridan, J.; Marsters, S.; Pitti, R.; Gurney, A.; Skubatch, M.; Baldwin, D.; Ramakrishnan, L.; Gray, C.; Baker, K.; Wood, W.; et al. Control of TRAIL-Induced Apoptosis by a Family of Signaling and Decoy Receptors. Science 1997, 277, 818–821. [Google Scholar] [CrossRef]
- Verma, S.; Loewendorf, A.; Wang, Q.; McDonald, B.; Redwood, A.; Benedict, C.A. Inhibition of the TRAIL Death Receptor by CMV Reveals Its Importance in NK Cell-Mediated Antiviral Defense. PLoS Pathog. 2014, 10, e1004268. [Google Scholar] [CrossRef]
- Sedger, L.M.; Shows, D.M.; Blanton, R.A.; Peschon, J.J.; Goodwin, R.G.; Cosman, D.; Wiley, S.R. IFN-Gamma Mediates a Novel Antiviral Activity through Dynamic Modulation of TRAIL and TRAIL Receptor Expression. J. Immunol. Baltim. 1999, 163, 920–926. [Google Scholar]
- Wu, Z.; Sinzger, C.; Frascaroli, G.; Reichel, J.; Bayer, C.; Wang, L.; Schirmbeck, R.; Mertens, T. Human Cytomegalovirus-Induced NKG2Chi CD57hi Natural Killer Cells Are Effectors Dependent on Humoral Antiviral Immunity. J. Virol. 2013, 87, 7717–7725. [Google Scholar] [CrossRef]
- Lopez-Vergès, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a Unique CD57+NKG2Chi Natural Killer Cell Subset during Acute Human Cytomegalovirus Infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef]
- Wang, D.; Bresnahan, W.; Shenk, T. Human Cytomegalovirus Encodes a Highly Specific RANTES Decoy Receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 16642–16647. [Google Scholar] [CrossRef]
- Mansfield, A.S.; Nevala, W.K.; Dronca, R.S.; Leontovich, A.A.; Shuster, L.; Markovic, S.N. Normal Ageing Is Associated with an Increase in Th2 Cells, MCP-1 (CCL1) and RANTES (CCL5), with Differences in SCD40L and PDGF-AA between Sexes. Clin. Exp. Immunol. 2012, 170, 186–193. [Google Scholar] [CrossRef]
- Bernasconi, S.; Cinque, P.; Peri, G.; Sozzani, S.; Crociati, A.; Torri, W.; Vicenzi, E.; Vago, L.; Lazzarin, A.; Poli, G.; et al. Selective Elevation of Monocyte Chemotactic Protein-1 in the Cerebrospinal Fluid of AIDS Patients with Cytomegalovirus Encephalitis. J. Infect. Dis. 1996, 174, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Netea, M.G.; Dinarello, C.A.; Joosten, L.A.B. Inflammasome Activation and IL-1β and IL-18 Processing during Infection. Trends Immunol. 2011, 32, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P.-Y. Inflammasomes: Mechanism of Action, Role in Disease, and Therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef]
- Shi, X.; Dong, Y.; Li, Y.; Zhao, Z.; Li, H.; Qiu, S.; Li, Y.; Guo, W.; Qiao, Y. Inflammasome Activation in Mouse Inner Ear in Response to MCMV Induced Hearing Loss. J. Otol. 2015, 10, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Hokeness, K.L.; Deweerd, E.S.; Munks, M.W.; Lewis, C.A.; Gladue, R.P.; Salazar-Mather, T.P. CXCR3-Dependent Recruitment of Antigen-Specific T Lymphocytes to the Liver during Murine Cytomegalovirus Infection. J. Virol. 2007, 81, 1241–1250. [Google Scholar] [CrossRef]
- Basílio-Queirós, D.; Venturini, L.; Luther-Wolf, S.; Dammann, E.; Ganser, A.; Stadler, M.; Falk, C.S.; Weissinger, E.M. Adaptive NK Cells Undergo a Dynamic Modulation in Response to Human Cytomegalovirus and Recruit T Cells in in Vitro Migration Assays. Bone Marrow Transplant. 2022, 57, 712–720. [Google Scholar] [CrossRef]
- Van de Berg, P.J.; Heutinck, K.M.; Raabe, R.; Minnee, R.C.; Young, S.L.; van Donselaar-van der Pant, K.A.; Bemelman, F.J.; van Lier, R.A.; ten Berge, I.J. Human Cytomegalovirus Induces Systemic Immune Activation Characterized by a Type 1 Cytokine Signature. J. Infect. Dis. 2010, 202, 690–699. [Google Scholar] [CrossRef]
- Cheeran, M.C.; Hu, S.; Yager, S.L.; Gekker, G.; Peterson, P.K.; Lokensgard, J.R. Cytomegalovirus Induces Cytokine and Chemokine Production Differentially in Microglia and Astrocytes: Antiviral Implications. J. Neurovirol. 2001, 7, 135–147. [Google Scholar] [CrossRef]
- Lee, J.; Choi, J.A.; Ju, H.-H.; Kim, J.-E.; Paik, S.-Y.; Rao, P.V. Role of MCP-1 and IL-8 in Viral Anterior Uveitis, and Contractility and Fibrogenic Activity of Trabecular Meshwork Cells. Sci. Rep. 2021, 11, 14950. [Google Scholar] [CrossRef]
- Rott, D.; Zhu, J.; Zhou, Y.F.; Burnett, M.S.; Zalles-Ganley, A.; Epstein, S.E. IL-6 Is Produced by Splenocytes Derived from CMV-Infected Mice in Response to CMV Antigens, and Induces MCP-1 Production by Endothelial Cells: A New Mechanistic Paradigm for Infection-Induced Atherogenesis. Atherosclerosis 2003, 170, 223–228. [Google Scholar] [CrossRef]
- Froberg, M.K.; Dannen, D.; Adams, A.; Parker-Thornburg, J.; Kolattukudy, P. Murine Cytomegalovirus Infection Markedly Reduces Serum MCP-1 Levels in MCP-1 Transgenic Mice. Ann. Clin. Lab. Sci. 2006, 36, 179–184. [Google Scholar] [PubMed]
- Alcendor, D.J.; Charest, A.M.; Zhu, W.Q.; Vigil, H.E.; Knobel, S.M. Infection and Upregulation of Proinflammatory Cytokines in Human Brain Vascular Pericytes by Human Cytomegalovirus. J. Neuroinflamm. 2012, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Talaya, A.; Giménez, E.; Vinuesa, V.; Pérez, A.; Amat, P.; Piñana, J.L.; Albert, E.; Hernández-Boluda, J.C.; Solano, C.; Navarro, D. Kinetics of Inflammatory Biomarkers in Plasma Predict the Occurrence and Features of Cytomegalovirus DNAemia Episodes in Allogeneic Hematopoietic Stem Cell Transplant Recipients. Med. Microbiol. Immunol. 2019, 208, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Karason, K.; Jernås, M.; Hägg, D.A.; Svensson, P.-A. Evaluation of CXCL9 and CXCL10 as Circulating Biomarkers of Human Cardiac Allograft Rejection. BMC Cardiovasc. Disord. 2006, 6, 29. [Google Scholar] [CrossRef]
- Poole, E.; Neves, T.C.; Oliveira, M.T.; Sinclair, J.; da Silva, M.C.C. Human Cytomegalovirus Interleukin 10 Homologs: Facing the Immune System. Front. Cell. Infect. Microbiol. 2020, 10, 245. [Google Scholar] [CrossRef]
- Lučin, P.; Mahmutefendić, H.; Blagojević Zagorac, G.; Ilić Tomaš, M. Cytomegalovirus Immune Evasion by Perturbation of Endosomal Trafficking. Cell. Mol. Immunol. 2015, 12, 154–169. [Google Scholar] [CrossRef]
- Billstrom, M.A.; Lehman, L.A.; Scott Worthen, G. Depletion of Extracellular RANTES during Human Cytomegalovirus Infection of Endothelial Cells. Am. J. Respir. Cell Mol. Biol. 1999, 21, 163–167. [Google Scholar] [CrossRef]
- Desveaux, C.; Klein, J.; Leruez-Ville, M.; Ramirez-Torres, A.; Lacroix, C.; Breuil, B.; Froment, C.; Bascands, J.-L.; Schanstra, J.P.; Ville, Y. Identification of Symptomatic Fetuses Infected with Cytomegalovirus Using Amniotic Fluid Peptide Biomarkers. PLOS Pathog. 2016, 12, e1005395. [Google Scholar] [CrossRef]
- Vorontsov, O.; Levitt, L.; Lilleri, D.; Vainer, G.W.; Kaplan, O.; Schreiber, L.; Arossa, A.; Spinillo, A.; Furione, M.; Alfi, O.; et al. Amniotic Fluid Biomarkers Predict the Severity of Congenital Cytomegalovirus Infection. J. Clin. Investig. 2022, 132, e157415. [Google Scholar] [CrossRef]
- Jacquemard, F.; Yamamoto, M.; Costa, J.-M.; Romand, S.; Jaqz-Aigrain, E.; Dejean, A.; Daffos, F.; Ville, Y. Maternal Administration of Valaciclovir in Symptomatic Intrauterine Cytomegalovirus Infection. BJOG Int. J. Obstet. Gynaecol. 2007, 114, 1113–1121. [Google Scholar] [CrossRef]
- Shahar-Nissan, K.; Pardo, J.; Peled, O.; Krause, I.; Bilavsky, E.; Wiznitzer, A.; Hadar, E.; Amir, J. Valaciclovir to Prevent Vertical Transmission of Cytomegalovirus after Maternal Primary Infection during Pregnancy: A Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2020, 396, 779–785. [Google Scholar] [CrossRef]
- Faure-Bardon, V.; Fourgeaud, J.; Guilleminot, T.; Magny, J.-F.; Salomon, L.J.; Bernard, J.-P.; Leruez-Ville, M.; Ville, Y. First-Trimester Diagnosis of Congenital Cytomegalovirus Infection after Maternal Primary Infection in Early Pregnancy: Feasibility Study of Viral Genome Amplification by PCR on Chorionic Villi Obtained by CVS. Ultrasound Obstet. Gynecol. 2021, 57, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.M.; Arendt, L.M.; Zhou, W.; Baig, S.; Walter, S.R.; Buchsbaum, R.J.; Kuperwasser, C.; Walt, D.R. Ultra-Sensitive Protein Detection via Single Molecule Arrays towards Early Stage Cancer Monitoring. Sci. Rep. 2015, 5, 11034. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uninfected Fetuses n = 40 | Infected Fetuses n = 40 | Asymptomatic Infected Fetuses n = 9 | Symptomatic Non-Severe Infected Fetuses n = 13 | Symptomatic and Severe Infected Fetuses n = 18 | p-Value * | p-Value ** | |
|---|---|---|---|---|---|---|---|
| Maternal age (years) | 32 (30; 35) | 32 (30; 36) | 33 (32; 36) | 32 (30; 33) | 30 (30; 34) | 0.9 | 0.4 |
| BMI (kg.m−2) | 20.70 (19.25; 21.75) | 21.00 (19.70; 22.70) | 20.70 (20.00; 23.40) | 21.00 (20.62; 22.00) | 21.20 (19.40; 24.10) | 0.3 | 0.9 |
| Nulliparity | 7 (19%) | 6 (16%) | 0 (0%) | 3 (23%) | 3 (19%) | 0.7 | 0.7 |
| GA amniocentesis (WG) | 22.0 (20.0; 25.0) | 22.1 (20.6; 24.9) | 20.7 (20.3; 22.3) | 23.6 (21.9; 27.0) | 22.0 (18.8; 25.1) | 0.8 | 0.2 |
| Uninfected Fetuses n = 40 | Infected Fetuses n = 40 | Asymptomatic Infected Fetuses n = 9 | Symptomatic Infected Fetuses n = 31 | Symptomatic Non-Severe Infected Fetuses n = 13 | Symptomatic and Severe Infected Fetuses n = 18 | p-Value * | p-Value ** | |
|---|---|---|---|---|---|---|---|---|
| IP10 interne | 135.945 (54.1875; 272.7225) | 451.405 (79.335; 956.49) | 118.04 (49.58; 605.312) | 505.33 (84.27; 1342.97) | 717.53 (84.06; 1170.79) | 315.52 (84.9775; 1419.735) | 0.0037 | 0.0006 |
| IP10 surface | 216.47 (71.6525; 393.6525) | 775.625 (300.34; 1680.205) | 264.38 (181.94; 704.448) | 892.91 (587.435; 2036.44) | 740.69 (566.94; 892.91) | 1374.01 (629.18; 3813.8825) | 0.0138 | 0.0005 |
| IL18 soluble | 4.206 (3.099; 7.05) | 7.416 (4.29; 10.698) | 6.48 (4.8; 9.24) | 8.124 (4.26; 11.562) | 5.16 (4.2; 7.2) | 10.308 (7.395; 11.85) | 0.0234 | 0.0060 |
| IP10 soluble | 10,852.986 (7354.974; 15,979.596) | 24,155.274 (13,671.278; 33,171.693) | 13,415.432 (7824.636; 28,769.988) | 24,195.876 (16,469.694; 36,399.324) | 27,967.932 (18,816.576; 39,577.848) | 24,155.274 (15,166.275; 28,695.609) | 0.0000 | 0.0001 |
| I-TAC soluble | 548.298 (384.462; 831.096) | 838.956 (588.57; 1682.418) | 822.924 (546.864; 1233.608) | 1015.44 (609.666; 1809.216) | 1015.44 (628.452; 1651.2) | 967.158 (601.602; 1825.788) | 0.0418 | 0.0547 |
| TRAIL soluble | 180.4938 (98.5061; 346.6263) | 299.2086 (141.0222; 802.9308) | 174.888 (141.1776; 224.6172) | 415.3464 (155.9106; 888.8652) | 245.2572 (140.556; 509.3052) | 589.3542 (219.0777; 1095.1194) | 0.0123 | 0.0004 |
| Uninfected Fetuses n = 40 | Infected Fetuses n = 40 | Asymptomatic Infected Fetuses n = 9 | Symptomatic Infected Fetuses n = 31 | Symptomatic Non-Severe Infected Fetuses n = 13 | Symptomatic and Severe infected Fetuses n = 18 | p-Value * | p-Value ** | |
|---|---|---|---|---|---|---|---|---|
| IP10 internal | 135.945 (54.1875; 272.7225) | 451.405 (79.335; 956.49) | 118.04 (49.58; 605.312) | 505.33 (84.27; 1342.97) | 717.53 (84.06; 1170.79) | 315.52 (84.9775; 1419.735) | 0.0037 | 0.0006 |
| MCP1 internal | 0 (0; 2.39) | 0.375 (0; 5.415) | 0 (0; 2.57) | 0.75 (0; 5.52) | 0.75 (0; 3.66) | 2.13 (0; 9.1625) | 0.0636 | 0.0156 |
| MIG internal | 0 (0; 9.7825) | 0.975 (0; 17.695) | 0 (0; 0) | 3.7 (0; 18.655) | 1.05 (0; 17.49) | 10.05 (0; 18.8275) | 0.1480 | 0.0309 |
| RANTES internal | 0 (0; 1.8625) | 1.65 (0; 5.65) | 0 (0; 2.89) | 2.08 (0; 6.29) | 0 (0; 1.58) | 5.505 (1.81; 11.0875) | 0.0585 | 0.0046 |
| IP10 surface | 216.47 (71.6525; 393.6525) | 775.625 (300.34; 1680.205) | 264.38 (181.94; 704.448) | 892.91 (587.435; 2036.44) | 740.69 (566.94; 892.91) | 1374.01 (629.18; 3813.8825) | 0.0138 | 0.0005 |
| CRP surface | 2993.57 (1885.88; 5824.6725) | 3253.165 (2054.605; 6279.77) | 2893.28 (1733.97; 3241.92) | 3319.65 (2095.535; 7397.78) | 2121.79 (1816.45; 3264.41) | 6068.58 (3347.9625; 14,867.88) | 0.1388 | 0.0035 |
| TRAIL surface | 77.8392 (36.0268; 216.2042) | 90.6635 (21.82125; 218.6225) | 25.688 (7.6784; 101.64) | 125.625 (27.66375; 229.949) | 89.695 (21.015; 133.802) | 220.6135 (62.4187; 410.7662) | 0.3280 | 0.0302 |
| IL18 soluble | 4.206 (3.099; 7.05) | 7.416 (4.29; 10.698) | 6.48 (4.8; 9.24) | 8.124 (4.26; 11.562) | 5.16 (4.2; 7.2) | 10.308 (7.395; 11.85) | 0.0234 | 0.0060 |
| IP10 soluble | 10,852.986 (7354.974; 15,979.596) | 24,155.274 (13,671.278; 33,171.693) | 13,415.432 (7824.636; 28,769.988) | 24,195.876 (16,469.694; 36,399.324) | 27,967.932 (18,816.576; 39,577.848) | 24,155.274 (15,166.275; 28,695.609) | 0.0000 | 0.0001 |
| CRP soluble | 725.232 (196.254; 1356.666) | 757.836 (352.305; 2831.577) | 646.2 (356.676; 772.752) | 898.992 (374.388; 2997.594) | 699.528 (168.756; 898.992) | 2841.414 (675.408; 7620.282) | 0.0987 | 0.0044 |
| TRAIL soluble | 180.4938 (98.5061; 346.6263) | 299.2086 (141.0222; 802.9308) | 174.888 (141.1776; 224.6172) | 415.3464 (155.9106; 888.8652) | 245.2572 (140.556; 509.3052) | 589.3542 (219.0777; 1095.1194) | 0.0123 | 0.0004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourgon, N.; Fitzgerald, W.; Aschard, H.; Magny, J.-F.; Guilleminot, T.; Stirnemann, J.; Romero, R.; Ville, Y.; Margolis, L.; Leruez-Ville, M. Cytokine Profiling of Amniotic Fluid from Congenital Cytomegalovirus Infection. Viruses 2022, 14, 2145. https://doi.org/10.3390/v14102145
Bourgon N, Fitzgerald W, Aschard H, Magny J-F, Guilleminot T, Stirnemann J, Romero R, Ville Y, Margolis L, Leruez-Ville M. Cytokine Profiling of Amniotic Fluid from Congenital Cytomegalovirus Infection. Viruses. 2022; 14(10):2145. https://doi.org/10.3390/v14102145
Chicago/Turabian StyleBourgon, Nicolas, Wendy Fitzgerald, Hugues Aschard, Jean-François Magny, Tiffany Guilleminot, Julien Stirnemann, Roberto Romero, Yves Ville, Leonid Margolis, and Marianne Leruez-Ville. 2022. "Cytokine Profiling of Amniotic Fluid from Congenital Cytomegalovirus Infection" Viruses 14, no. 10: 2145. https://doi.org/10.3390/v14102145