
3-Indoleacetonitrile Is Highly Effective in Treating Influenza A Virus Infection In Vitro and In Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compounds
2.2. Cells and Viruses
2.3. Cell Viability Assay
2.4. Influenza Virus Manipulation
2.5. Flow Cytometry Analysis
2.6. Real-Time Quantitative PCR (RT-qPCR) Analysis
2.7. Western Blotting Analysis
2.8. Mouse Experiments
2.9. Statistics
3. Results
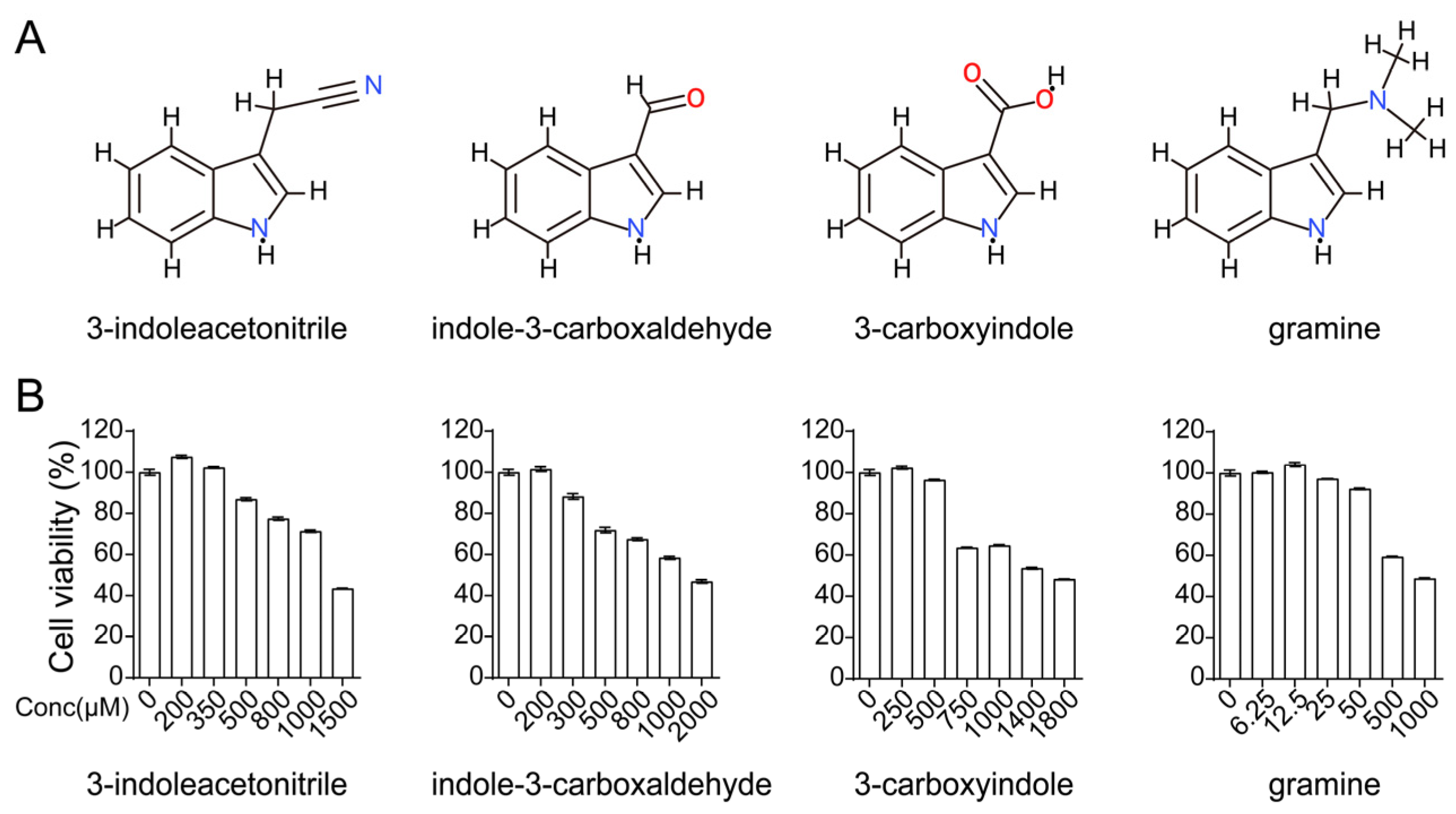
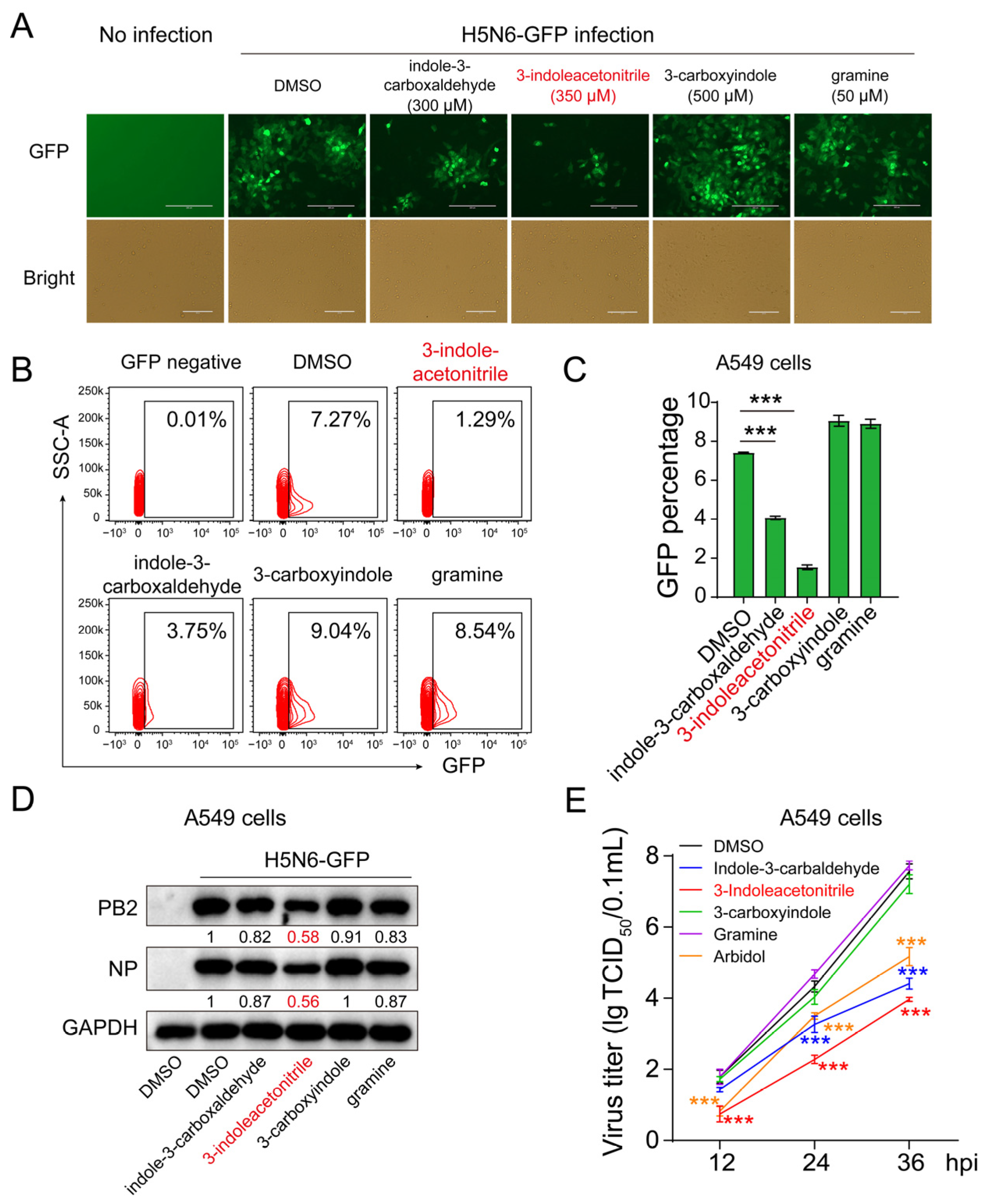
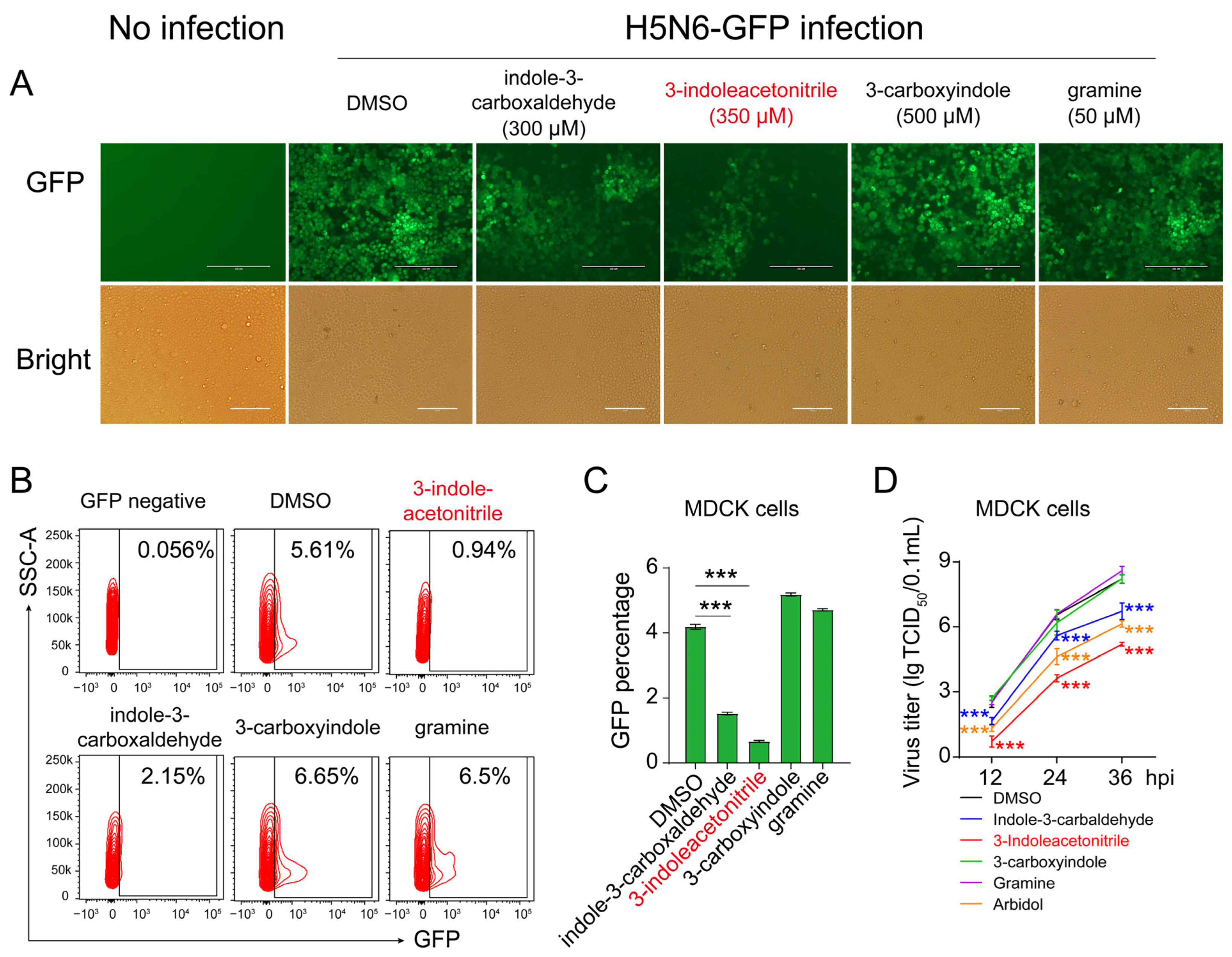
3.1. Screening 3-Indoleacetonitrile as an Antiviral Agent against IAV Infection
3.2. 3-Indoleacetonitrile Exerts Broad-Spectrum Activity against the Influenza A Virus
3.3. 3-Indoleacetonitrile Reduces Viral Replication in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krammer, F.; Smith, G.J.D.; Fouchier, R.; Peiris, J.S.M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Trilla, A.; Trilla, G.; Daer, C. The 1918 “Spanish Flu” in Spain. Clin. Infect. Dis. 2008, 47, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Jester, B.J.; Uyeki, T.M.; Jernigan, D.B. Fifty Years of Influenza A(H3N2) Following the Pandemic of 1968. Am. J. Public Health 2020, 110, 669–676. [Google Scholar] [CrossRef]
- LaRussa, P. Pandemic Novel 2009 H1N1 Influenza: What Have We Learned? Semin. Respir. Crit. Care Med. 2011, 32, 393–399. [Google Scholar] [CrossRef]
- Buchy, P.; Badur, S. Who and When to Vaccinate Against Influenza. Int. J. Infect. Dis. 2020, 93, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.D.; Chung, J.R.; Kim, S.S.; Gaglani, M.; Raiyani, C.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; et al. Interim Estimates of 2018–19 Seasonal Influenza Vaccine Effectiveness—United States, February 2019. MMWR. Morb. Mortal. Wkly. Rep. 2019, 68, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Yamayoshi, S.; Kawaoka, Y. Current and Future Influenza Vaccines. Nat. Med. 2019, 25, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted Influenza Vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.; Andrews, N.; Stellitano, L.; Stowe, J.; Winstone, A.M.; Shneerson, J.; Verity, C. Risk of Narcolepsy in Children and Young People Receiving AS03 Adjuvanted Pandemic A/H1N1 2009 Influenza Vaccine: Retrospective Analysis. BMJ 2013, 346, f794. [Google Scholar] [CrossRef] [Green Version]
- Nohynek, H.; Jokinen, J.J.; Partinen, M.; Vaarala, O.; Kirjavainen, T.; Sundman, J.; Himanen, S.-L.; Hublin, C.; Julkunen, I.; Olsén, P.; et al. AS03 Adjuvanted AH1N1 Vaccine Associated with an Abrupt Increase in the Incidence of Childhood Narcolepsy in Finland. PLoS ONE 2012, 7, e33536. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.; Whitley, R.J.; Pocock, S.; Monto, A.S. Oseltamivir Treatment for Influenza in Adults: A Meta-Analysis of randomised Controlled Trials. Lancet 2015, 385, 1729–1737. [Google Scholar] [CrossRef]
- Ishiguro, N.; Koseki, N.; Kaiho, M.; Ariga, T.; Kikuta, H.; Oba, K.; Togashi, T.; Morita, K.; Inagawa, A.; Okamura, A.; et al. Clinical Effectiveness of Four Neuraminidase Inhibitors (Oseltamivir, Zanamivir, Laninamivir, and Peramivir) for Children with Influenza A and B in the 2014–2015 to 2016–2017 Influenza Seasons in Japan. J. Infect. Chemother. 2018, 24, 449–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.; Ma, C.; Ohigashi, Y.; Oliveira, F.A.; Jardetzky, T.S.; Pinto, L.H.; Lamb, R.A. Functional Studies Indicate Amantadine Binds to the Pore of the Influenza A Virus M2 Proton-Selective Ion Channel. Proc. Natl. Acad. Sci. USA 2008, 105, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Beigel, J.H.; Hayden, F.G. Influenza Therapeutics in Clinical Practice—Challenges and Recent Advances. Cold Spring Harb. Perspect. Med. 2020, 11, a038463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ison, M.G. Antivirals and Resistance: Influenza Virus. Curr. Opin. Virol. 2011, 1, 563–573. [Google Scholar] [CrossRef]
- Hurt, A.C. Antiviral Therapy for the Next Influenza Pandemic. Trop. Med. Infect. Dis. 2019, 4, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Xie, Q.; Bian, G.; Zhang, B.; Wang, M.; Wang, Y.; Chen, Z.; Li, Y. Anti-H1N1 Viral Activity of Three Main Active Ingredients From Zedoary Oil. Fitoterapia 2020, 142, 104489. [Google Scholar] [CrossRef]
- Sithisarn, P.; Michaelis, M.; Schubert-Zsilavecz, M.; Cinatl, J. Differential Antiviral and Anti-Inflammatory Mechanisms of the Flavonoids Biochanin A and Baicalein in H5N1 Influenza A Virus-Infected Cells. Antivir. Res. 2013, 97, 41–48. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, Z.; Huo, C.; Guo, X.; Yang, G.; Wang, M.; Tian, H.; Hu, Y.; Dong, H. Antiviral Effects of Shuanghuanglian Injection Powder Against Influenza A Virus H5N1 In Vitro and In Vivo. Microb. Pathog. 2018, 121, 318–324. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.; Lu, J.; Chen, Q.; Ti, H.; Huang, W.; Li, J.; Yang, Z.; Jiang, Z.; Wang, X. Inhibition of Influenza Virus Via a Sesquiterpene Fraction Isolated from Laggera Pterodonta by targeTing the NF-κB and p38 Pathways. BMC Complement. Altern. Med. 2017, 17, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Chen, S.; Li, Y.; Zhang, A.; Zhou, H.; Chen, H.; Jin, M. 14-Deoxy-11,12-Didehydroandrographolide Attenuates Excessive Inflammatory Responses and Protects Mice Lethally Challenged with Highly Pathogenic A(H5N1) Influenza Viruses. Antivir. Res. 2016, 133, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Li, Y.; Chen, S.; Wang, M.; Zhang, A.; Zhou, H.; Chen, H.; Jin, M. 14-Deoxy-11,12-dehydroandrographolide exerts anti-influenza A virus activity and inhibits replication of H5N1 virus by restraining nuclear export of viral ribonucleoprotein complexes. Antivir. Res. 2015, 118, 82–92. [Google Scholar] [CrossRef]
- Cai, W.; Wen, H.; Zhou, Q.; Wu, L.; Chen, Y.; Zhou, H.; Jin, M. 14-Deoxy-11,12-Didehydroandrographolide Inhibits Apoptosis in Influenza A(H5N1) Virus-Infected Human Lung Epithelial Cells Via the Caspase-9-Dependent Intrinsic Apoptotic Pathway which Contributes to Its Antiviral Activity. Antivir. Res. 2020, 181, 104885. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Cho, H.S.; Kim, Y.; Kim, J.-A.; Banskota, S.; Cho, M.H.; Lee, J. Indole and 7-Benzyloxyindole Attenuate the Virulence of Staphylococcus Aureus. Appl. Microbiol. Biotechnol. 2013, 97, 4543–4552. [Google Scholar] [CrossRef]
- Dadashpour, S.; Emami, S. Indole in the Target-Based Design of Anticancer Agents: A Versatile Scaffold with Diverse Mechanisms. Eur. J. Med. Chem. 2018, 150, 9–29. [Google Scholar] [CrossRef]
- Mehndiratta, S.; Hsieh, Y.-L.; Liu, Y.-M.; Wang, A.W.; Lee, H.-Y.; Liang, L.-Y.; Kumar, S.; Teng, C.-M.; Yang, C.-R.; Liou, J.-P. Indole-3-Ethylsulfamoylphenylacrylamides: Potent Histone Deacetylase Inhibitors with Anti-Inflammatory Activity. Eur. J. Med. Chem. 2014, 85, 468–479. [Google Scholar] [CrossRef]
- Blaising, J.; Polyak, S.J.; Pécheur, E.-I. Arbidol as a Broad-Spectrum Antiviral: An Update. Antivir. Res. 2014, 107, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Nasser, Z.H.; Swaminathan, K.; Müller, P.; Downard, K.M. Inhibition of Influenza Hemagglutinin with the Antiviral Inhibitor Arbidol Using a Proteomics Based Approach and Mass Spectrometry. Antivir. Res. 2013, 100, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Kadam, R.U.; Wilson, I.A. Structural Basis of Influenza Virus Fusion Inhibition by the Antiviral Drug Arbidol. Proc. Natl. Acad. Sci. USA 2017, 114, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Perez, J.T.; García-Sastre, A.; Manicassamy, B. Insertion of a GFP Reporter Gene in Influenza Virus. Curr. Protoc. Microbiol. 2013, 29, 15G.4.1–15G.4.16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Lin, X.; Qin, C.; Huang, K.; Sun, X.; Zhao, L.; Jin, M. Avian Chaperonin Containing TCP1 Subunit 5 Supports Influenza A Virus Replication by Interacting With Viral Nucleoprotein, PB1, and PB2 Proteins. Front. Microbiol. 2020, 11, 538355. [Google Scholar] [CrossRef]
- Kutáček, M.; Procházka, Ž.; Grünberger, D. Biogenesis of Ascorbigen, 3-Indolylacetonitrile and Indole-3-carboxylic Acid from D,L-Tryptophan-3-14C in Brassica oleracea L. Nature 1960, 187, 61–62. [Google Scholar] [CrossRef]
- Wang, X.; Jia, W.; Zhao, A.; Wang, X. Anti-Influenza Agents from Plants and Traditional Chinese Medicine. Phytother. Res. 2006, 20, 335–341. [Google Scholar] [CrossRef]
- Böttcher, C.; Chapman, A.; Fellermeier, F.; Choudhary, M.; Scheel, D.; Glawischnig, E. The Biosynthetic Pathway of Indole-3-Carbaldehyde and Indole-3-Carboxylic Acid Derivatives in Arabidopsis. Plant Physiol. 2014, 165, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Patiño, R.; Rashel, R.; Rubio, A.; Longing, S. Growth-Suppressing and Algicidal Properties of an Extract from Arundo Donax, an Invasive Riparian Plant, Against Prymnesium Parvum, an Invasive Harmful Alga. Harmful Algae 2018, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hautala, E.; Holopainen, J. Gramine and Free Amino Acids as Indicators of Fluoride-Induced Stress in Barley and Its Consequences to Insect Herbivory. Ecotoxicol. Environ. Saf. 1995, 31, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Goelz, M.F.; Rothenbacher, H.; Wiggins, J.; Kendall, W.; Hershberger, T. Some Hematological and Histopathological Effects of the Alkaloids Gramine and Hordenine on Meadow Voles (Microtus Pennsylvanicus). Toxicology 1980, 18, 125–131. [Google Scholar] [CrossRef]
- Wei, Y.; Shi, L.; Wang, K.; Liu, M.; Yang, Q.; Yang, Z.; Ke, S. Discovery of Gramine Derivatives That Inhibit the Early Stage of EV71 Replication in Vitro. Molecules 2014, 19, 8949–8964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Cho, M.H.; Lee, J. 3-Indolylacetonitrile Decreases Escherichia coliO157:H7 Biofilm Formation and Pseudomonas Aeruginosa Virulence. Environ. Microbiol. 2010, 13, 62–73. [Google Scholar] [CrossRef]
- Lee, S.; Jin, G.; Kim, D.; Son, S.; Lee, K.; Lee, C. Indole Derivatives Inhibit Hepatitis C Virus Replication Through Induction of Pro-inflammatory Cytokines. Acta Virol. 2015, 59, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Brancato, V.; Peduto, A.; Wharton, S.; Martin, S.; More, V.; Di Mola, A.; Massa, A.; Perfetto, B.; Donnarumma, G.; Schiraldi, C.; et al. Design of Inhibitors of Influenza Virus Membrane Fusion: Synthesis, Structure–Activity Relationship and In Vitro Antiviral Activity of a Novel Indole Series. Antivir. Res. 2013, 99, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Bardiot, D.; Koukni, M.; Smets, W.; Carlens, G.; McNaughton, M.; Kaptein, S.; Dallmeier, K.; Chaltin, P.; Neyts, J.; Marchand, A. Discovery of Indole Derivatives as Novel and Potent Dengue Virus Inhibitors. J. Med. Chem. 2018, 61, 8390–8401. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Zhao, Y.; Zhao, C.; Gong, P. Synthesis and In Vitro Anti-Hepatitis B Virus Activities of Some Ethyl 6-Bromo-5-Hydroxy-1H-Indole-3-Carboxylates. Bioorg. Med. Chem. 2006, 14, 911–917. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhao, L.; Zhao, Y.; Huang, K.; Gong, W.; Yang, Y.; Zhao, L.; Xia, X.; Li, Z.; Sheng, F.; et al. 3-Indoleacetonitrile Is Highly Effective in Treating Influenza A Virus Infection In Vitro and In Vivo. Viruses 2021, 13, 1433. https://doi.org/10.3390/v13081433
Zhao X, Zhao L, Zhao Y, Huang K, Gong W, Yang Y, Zhao L, Xia X, Li Z, Sheng F, et al. 3-Indoleacetonitrile Is Highly Effective in Treating Influenza A Virus Infection In Vitro and In Vivo. Viruses. 2021; 13(8):1433. https://doi.org/10.3390/v13081433
Chicago/Turabian StyleZhao, Xuejin, Lianzhong Zhao, Ya Zhao, Kun Huang, Wenxiao Gong, Ying Yang, Li Zhao, Xiaohan Xia, Zaiyun Li, Feng Sheng, and et al. 2021. "3-Indoleacetonitrile Is Highly Effective in Treating Influenza A Virus Infection In Vitro and In Vivo" Viruses 13, no. 8: 1433. https://doi.org/10.3390/v13081433