
West Nile Virus Vaccination Protects against Usutu Virus Disease in Mice

 , and
, and
Abstract
:1. Introduction
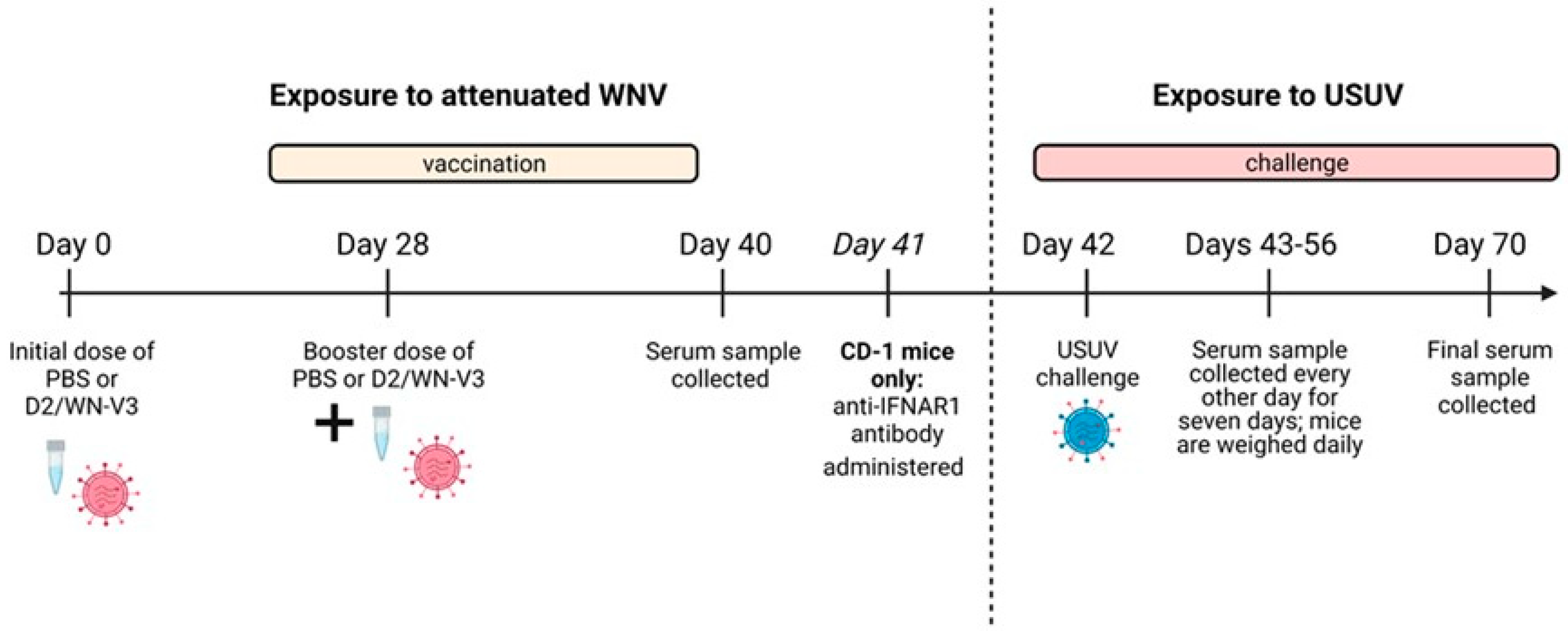
2. Materials and Methods
2.1. Viruses and Cells
2.2. Inoculation of Mice
2.2.1. Ifnar1−/− Mice
2.2.2. CD-1 Mice
2.3. Plaque Reduction Neutralization Test (PRNT)
2.4. Statistics
2.5. Additional Software
3. Results
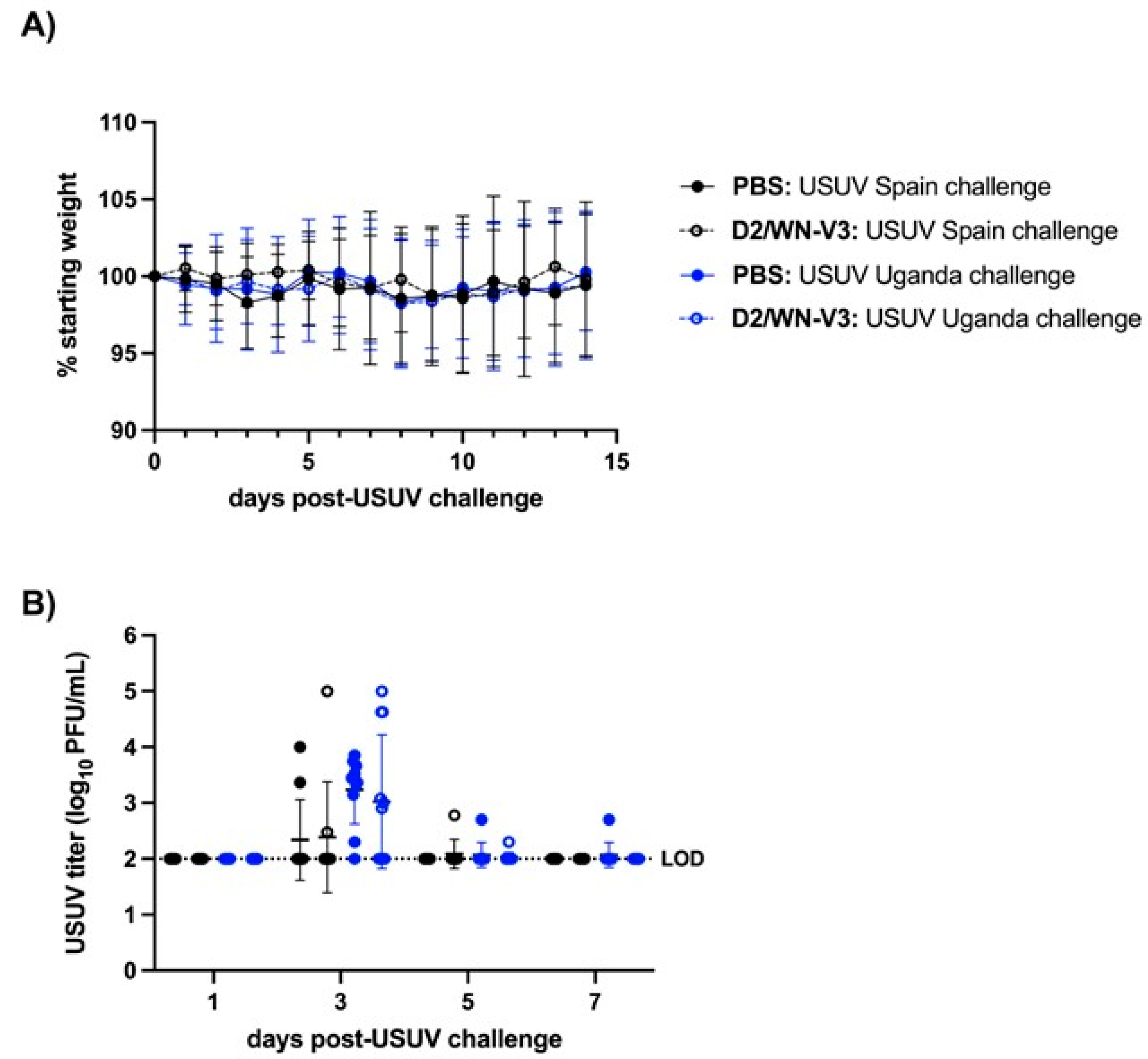
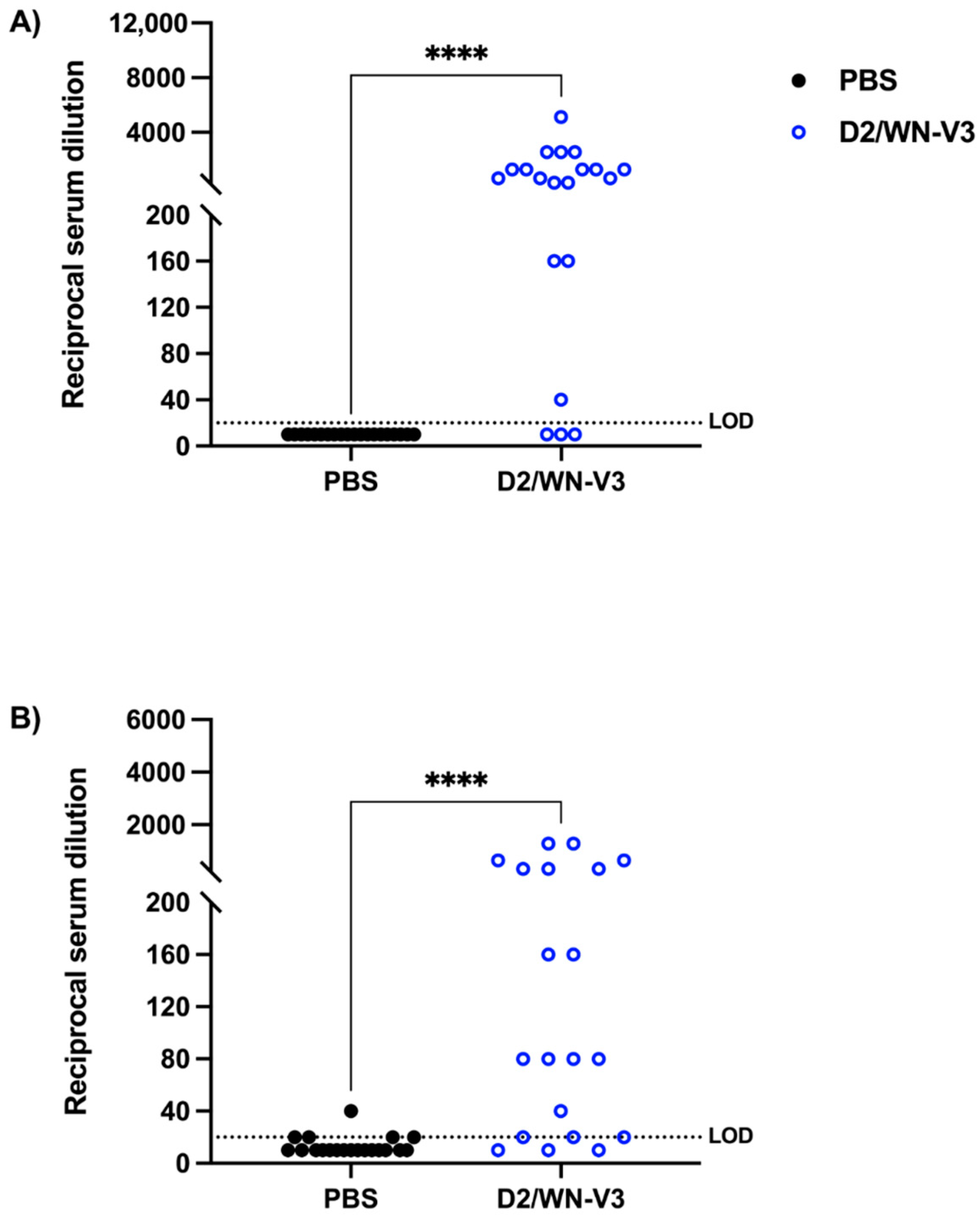
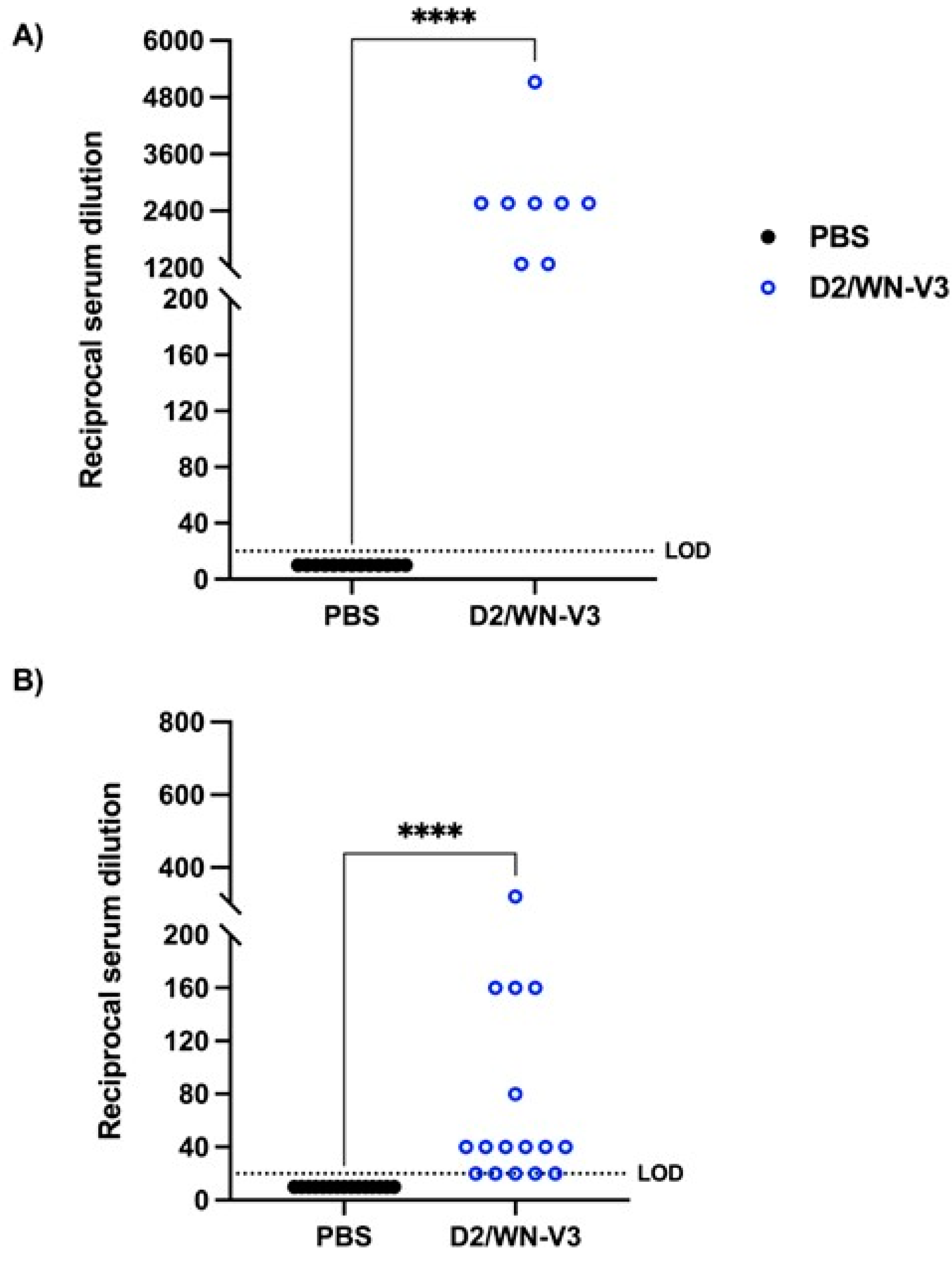
3.1. Vaccinated CD-1 Mice Produce a Neutralizing Response against WNV and USUV Prior to USUV Challenge
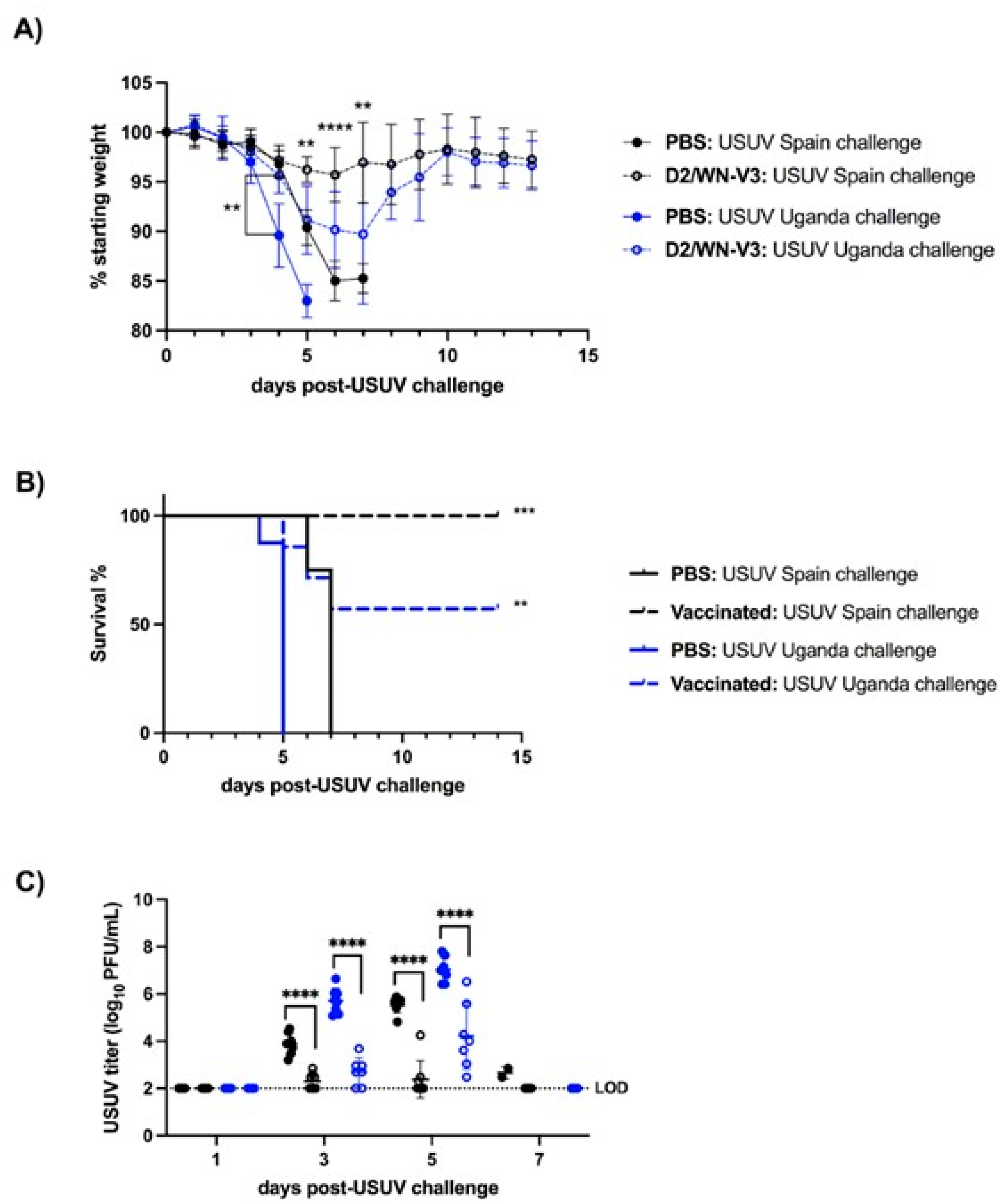
3.2. WNV Vaccination Protects Ifnar1−/− Mice from USUV Disease and Viremia
3.3. WNV Vaccination Induces a Neutralizing Response against WNV and USUV in Ifnar1−/− Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu Virus: An Arbovirus on the Rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Smithburn, K.H.T.; Burke, A. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Melnick, J.L.; Paul, J.R.; Riordan, J.T.; Barnett, V.H.; Goldblum, N.; Zabin, E. Isolation from human sera in Egypt of a virus apparently identical to West Nile virus. Proc. Soc. Exp. Biol. Med. 1951, 77, 661–665. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, B.M.; Jupp, P.G.; Dos Santos, I.; Meenehan, G.M. Epidemics of West Nile and Sindbis viruses in South Africa with Culex (Culex) univittatus Theobald as vector. S. Afr. J. Sci. 1976, 72, 295–300. [Google Scholar]
- Hurlbut, H.S.; Rizk, F.; Taylor, R.M.; Work, T.H. A study of the ecology of West Nile virus in Egypt. Am. J. Trop. Med. Hyg. 1956, 5, 579–620. [Google Scholar] [CrossRef]
- Chowdhury, P.; Khan, S.A.; Dutta, P.; Topno, R.; Mahanta, J. Characterization of West Nile virus (WNV) isolates from Assam, India: Insights into the circulating WNV in northeastern India. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 39–47. [Google Scholar] [CrossRef]
- Myint, K.S.; Kosasih, H.; Artika, I.M.; Perkasa, A.; Puspita, M.; Ma’roef, C.N.; Antonjaya, U.; Ledermann, J.P.; Powers, A.M.; Alisjahbana, B. West Nile virus documented in Indonesia from acute febrile illness specimens. Am. J. Trop. Med. Hyg. 2014, 90, 260–262. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Fu, S.H.; Liu, W.B.; Wang, H.Y.; Lu, Z.; Tong, S.X.; Li, Z.X.; Nasci, R.S.; Kosoy, O.; Cui, Y.; et al. West nile virus infection in Xinjiang, China. Vector Borne Zoonotic Dis. 2013, 13, 131–133. [Google Scholar] [CrossRef]
- Russell, R.C.; Dwyer, D.E. Arboviruses associated with human disease in Australia. Microbes Infect. 2000, 2, 1693–1704. [Google Scholar] [CrossRef]
- Frost, M.J.; Zhang, J.; Edmonds, J.H.; Prow, N.A.; Gu, X.; Davis, R.; Hornitzky, C.; Arzey, K.E.; Finlaison, D.; Hick, P.; et al. Characterization of virulent West Nile virus Kunjin strain, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 792–800. [Google Scholar] [CrossRef]
- Davis, C.T.; Ebel, G.D.; Lanciotti, R.S.; Brault, A.C.; Guzman, H.; Siirin, M.; Lambert, A.; Parsons, R.E.; Beasley, D.W.; Novak, R.J.; et al. Phylogenetic analysis of North American West Nile virus isolates, 2001–2004: Evidence for the emergence of a dominant genotype. Virology 2005, 342, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Ebel, G.D.; Carricaburu, J.; Young, D.; Bernard, K.A.; Kramer, L.D. Genetic and phenotypic variation of West Nile virus in New York, 2000–2003. Am. J. Trop. Med. Hyg. 2004, 71, 493–500. [Google Scholar] [CrossRef]
- Morales, M.A.; Barrandeguy, M.; Fabbri, C.; Garcia, J.B.; Vissani, A.; Trono, K.; Gutierrez, G.; Pigretti, S.; Menchaca, H.; Garrido, N.; et al. West Nile virus isolation from equines in Argentina, 2006. Emerg. Infect. Dis. 2006, 12, 1559–1561. [Google Scholar] [CrossRef]
- Bernkopf, H.; Levine, S.; Nerson, R. Isolation of West Nile virus in Israel. J. Infect. Dis. 1953, 93, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidemiology of the West Nile virus: Study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse. Ann. Inst. Pasteur. 1970, 118, 239–247. [Google Scholar] [PubMed]
- Hannoun, C.; Panthier, R.; Mouchet, J.; Eouzan, J.P. Isolation in France of the West Nile Virus from Patients and from the Vector Culex Modestus Ficalbi. Comptes Rendus Hebd. Seances Acad. Sci. 1964, 259, 4170–4172. [Google Scholar]
- Williams, M.C.; Simpson, D.I.; Haddow, A.J.; Knight, E.M. The Isolation of West Nile Virus from Man and of Usutu Virus from the Bird-Biting Mosquito Mansonia Aurites (Theobald) in the Entebbe Area of Uganda. Ann. Trop. Med. Parasitol. 1964, 58, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jost, H.; Wink, M.; Borstler, J.; Bosch, S.; Garigliany, M.M.; Jost, A.; Czajka, C.; Luhken, R.; Ziegler, U.; et al. Reconstruction of the Evolutionary History and Dispersal of Usutu Virus, a Neglected Emerging Arbovirus in Europe and Africa. mBio 2016, 7, e01938-15. [Google Scholar] [CrossRef] [Green Version]
- Luhken, R.; Jost, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu Virus in Germany and Its Effect on Breeding Bird Populations. Emerg. Infect. Dis. 2017, 23, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Weissenbock, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Pierro, A.; Gaibani, P.; Spadafora, C.; Ruggeri, D.; Randi, V.; Parenti, S.; Finarelli, A.C.; Rossini, G.; Landini, M.P.; Sambri, V. Detection of specific antibodies against West Nile and Usutu viruses in healthy blood donors in northern Italy, 2010–2011. Clin. Microbiol. Infect. 2013, 19, E451–E453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faggioni, G.; De Santis, R.; Pomponi, A.; Grottola, A.; Serpini, G.F.; Meacci, M.; Gennari, W.; Tagliazucchi, S.; Pecorari, M.; Monaco, F.; et al. Prevalence of Usutu and West Nile virus antibodies in human sera, Modena, Italy, 2012. J. Med. Virol. 2018, 90, 1666–1668. [Google Scholar] [CrossRef] [PubMed]
- Sinigaglia, A.; Pacenti, M.; Martello, T.; Pagni, S.; Franchin, E.; Barzon, L. West Nile virus infection in individuals with pre-existing Usutu virus immunity, northern Italy, 2018. Eurosurveillance 2019, 24, 1900261. [Google Scholar] [CrossRef] [Green Version]
- Aberle, S.W.; Kolodziejek, J.; Jungbauer, C.; Stiasny, K.; Aberle, J.H.; Zoufaly, A.; Hourfar, M.K.; Weidner, L.; Nowotny, N. Increase in human West Nile and Usutu virus infections, Austria, 2018. Eurosurveillance 2018, 23, 1800545. [Google Scholar] [CrossRef] [Green Version]
- Blazquez, A.B.; Escribano-Romero, E.; Martin-Acebes, M.A.; Petrovic, T.; Saiz, J.C. Limited susceptibility of mice to Usutu virus (USUV) infection and induction of flavivirus cross-protective immunity. Virology 2015, 482, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Silengo, S.J.; Whiteman, M.C.; Kinney, R.M. Chimeric dengue 2 PDK-53/West Nile NY99 viruses retain the phenotypic attenuation markers of the candidate PDK-53 vaccine virus and protect mice against lethal challenge with West Nile virus. J. Virol. 2005, 79, 7300–7310. [Google Scholar] [CrossRef] [Green Version]
- Butrapet, S.; Huang, C.Y.; Pierro, D.J.; Bhamarapravati, N.; Gubler, D.J.; Kinney, R.M. Attenuation markers of a candidate dengue type 2 vaccine virus, strain 16681 (PDK-53), are defined by mutations in the 5′ noncoding region and nonstructural proteins 1 and 3. J. Virol. 2000, 74, 3011–3019. [Google Scholar] [CrossRef] [Green Version]
- Yoksan, S.; Bhamarapravati, N.; Halstead, S.B. Dengue virus vaccine development: Study on biological markers of uncloned dengue 1–4 viruses serially passaged in primary kidney cells, Arbovirus Research in Australia. In Proceedings of the 4th Symposium CSIRO/QIMR, Brisbane, Australia, 15–17 February 1982; St. George, T.D., Kay, B.H., Block, J., Eds.; CSIRO/QIMR: Brisbane, Australia, 1986; pp. 35–38. [Google Scholar]
- Huang, C.Y.; Butrapet, S.; Tsuchiya, K.R.; Bhamarapravati, N.; Gubler, D.J.; Kinney, R.M. Dengue 2 PDK-53 virus as a chimeric carrier for tetravalent dengue vaccine development. J. Virol. 2003, 77, 11436–11447. [Google Scholar] [CrossRef] [Green Version]
- Calvert, A.E.; Huang, C.Y.; Kinney, R.M.; Roehrig, J.T. Non-structural proteins of dengue 2 virus offer limited protection to interferon-deficient mice after dengue 2 virus challenge. J. Gen. Virol. 2006, 87, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.A.; Chuong, C.; Hawks, S.A.; Rai, P.; Duggal, N.K.; Weger-Lucarelli, J. Development and characterization of infectious clones of two strains of Usutu virus. Virology 2021, 554, 28–36. [Google Scholar] [CrossRef]
- Kuchinsky, S.C.; Hawks, S.A.; Mossel, E.C.; Coutermarsh-Ott, S.; Duggal, N.K. Differential pathogenesis of Usutu virus isolates in mice. PLoS Negl. Trop. Dis. 2020, 14, e0008765. [Google Scholar] [CrossRef]
- Kinney, R.M.; Butrapet, S.; Chang, G.J.; Tsuchiya, K.R.; Roehrig, J.T.; Bhamarapravati, N.; Gubler, D.J. Construction of infectious cDNA clones for dengue 2 virus: Strain 16681 and its attenuated vaccine derivative, strain PDK-53. Virology 1997, 230, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, A.; Ruiz, S.; Herrero, L.; Moreno, J.; Molero, F.; Magallanes, A.; Sánchez-Seco, P.M.; Figuerola, J.; Tenorio, A. West Nile and Usutu viruses in mosquitoes in Spain, 2008–2009. Am. J. Trop. Med. Hyg. 2011, 85, 178. [Google Scholar] [CrossRef]
- Mossel, E.C.; Crabtree, M.B.; Mutebi, J.P.; Lutwama, J.J.; Borland, E.M.; Powers, A.M.; Miller, B.R. Arboviruses Isolated From Mosquitoes Collected in Uganda, 2008–2012. J. Med. Entomol. 2017, 54, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shresta, S.; Kyle, J.L.; Snider, H.M.; Basavapatna, M.; Beatty, P.R.; Harris, E. Interferon-dependent immunity is essential for resistance to primary dengue virus infection in mice, whereas T- and B-cell-dependent immunity are less critical. J. Virol. 2004, 78, 2701–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.J.; Roehrig, J.T. New mouse model for dengue virus vaccine testing. J. Virol. 1999, 73, 783–786. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Guidelines for Plaque Reduction Neutralization Testing of Human Antibodies to Dengue Viruses; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Oliphant, T.; Nybakken, G.E.; Austin, S.K.; Xu, Q.; Bramson, J.; Loeb, M.; Throsby, M.; Fremont, D.H.; Pierson, T.C.; Diamond, M.S. Induction of epitope-specific neutralizing antibodies against West Nile virus. J. Virol. 2007, 81, 11828–11839. [Google Scholar] [CrossRef] [Green Version]
- Nybakken, G.E.; Oliphant, T.; Johnson, S.; Burke, S.; Diamond, M.S.; Fremont, D.H. Structural basis of West Nile virus neutralization by a therapeutic antibody. Nature 2005, 437, 764–769. [Google Scholar] [CrossRef]
- Oliphant, T.; Engle, M.; Nybakken, G.E.; Doane, C.; Johnson, S.; Huang, L.; Gorlatov, S.; Mehlhop, E.; Marri, A.; Chung, K.M.; et al. Development of a humanized monoclonal antibody with therapeutic potential against West Nile virus. Nat. Med. 2005, 11, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Martin-Acebes, M.A.; Blazquez, A.B.; Canas-Arranz, R.; Vazquez-Calvo, A.; Merino-Ramos, T.; Escribano-Romero, E.; Sobrino, F.; Saiz, J.C. A recombinant DNA vaccine protects mice deficient in the alpha/beta interferon receptor against lethal challenge with Usutu virus. Vaccine 2016, 34, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Diamond, M.S. Role of CD8+ T cells in control of West Nile virus infection. J. Virol. 2004, 78, 8312–8321. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.; Samuel, M.A.; Diamond, M.S. CD8+ T cells require perforin to clear West Nile virus from infected neurons. J. Virol. 2006, 80, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitati, E.M.; Diamond, M.S. CD4+ T-cell responses are required for clearance of West Nile virus from the central nervous system. J. Virol. 2006, 80, 12060–12069. [Google Scholar] [CrossRef] [Green Version]
- Hassert, M.; Brien, J.D.; Pinto, A.K. Mouse Models of Heterologous Flavivirus Immunity: A Role for Cross-Reactive T Cells. Front. Immunol. 2019, 10, 1045. [Google Scholar] [CrossRef] [Green Version]
- Katzelnick, L.C.; Bos, S.; Harris, E. Protective and enhancing interactions among dengue viruses 1-4 and Zika virus. Curr. Opin. Virol. 2020, 43, 59–70. [Google Scholar] [CrossRef]
- Escribano-Romero, E.; Jimenez de Oya, N.; Camacho, M.C.; Blazquez, A.B.; Martin-Acebes, M.A.; Risalde, M.A.; Muriel, L.; Saiz, J.C.; Hofle, U. Previous Usutu Virus Exposure Partially Protects Magpies (Pica pica) against West Nile Virus Disease But Does Not Prevent Horizontal Transmission. Viruses 2021, 13, 1409. [Google Scholar] [CrossRef]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; et al. Evidence of simultaneous circulation of West Nile and Usutu viruses in mosquitoes sampled in Emilia-Romagna region (Italy) in 2009. PLoS ONE 2010, 5, e14324. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Abbo, S.R.; Visser, T.M.; Westenberg, M.; Geertsema, C.; Fros, J.J.; Koenraadt, C.J.M.; Pijlman, G.P. Competition between Usutu virus and West Nile virus during simultaneous and sequential infection of Culex pipiens mosquitoes. Emerg. Microbes. Infect. 2020, 9, 2642–2652. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WNV PRNT50 | USUV PRNT 50 | |||||
|---|---|---|---|---|---|---|
| Treatment | GMT | % seroconverted | # mice | GMT | % seroconverted | # mice |
| PBS (mock-vaccinated) | 10 | 0% | 0/20 | 12.31 | 25% | 5/20 |
| D2/WN-V3 (vaccinated) | 394 **** | 85% | 17/20 | 98.49 **** | 85% | 17/20 |
| WPN PRNT50 | USUV PRNT50 | |||||
|---|---|---|---|---|---|---|
| Treatment | GMT | % seroconverted | #mice | GMT | % seroconverted | #mice |
| PBS (mock-vaccinated) | 10 | 0% | 0/13 | 10 | 0% | 0/15 |
| D2/WN-V3 (vaccinated) | 2348 **** | 100% | 8/8 | 49.67 **** | 100% | 16/16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgado, R.; Hawks, S.A.; Frere, F.; Vázquez, A.; Huang, C.Y.-H.; Duggal, N.K. West Nile Virus Vaccination Protects against Usutu Virus Disease in Mice. Viruses 2021, 13, 2352. https://doi.org/10.3390/v13122352
Salgado R, Hawks SA, Frere F, Vázquez A, Huang CY-H, Duggal NK. West Nile Virus Vaccination Protects against Usutu Virus Disease in Mice. Viruses. 2021; 13(12):2352. https://doi.org/10.3390/v13122352
Chicago/Turabian StyleSalgado, Rebecca, Seth A. Hawks, Francesca Frere, Ana Vázquez, Claire Y.-H. Huang, and Nisha K. Duggal. 2021. "West Nile Virus Vaccination Protects against Usutu Virus Disease in Mice" Viruses 13, no. 12: 2352. https://doi.org/10.3390/v13122352