
Common Laboratory Mice Are Susceptible to Infection with the SARS-CoV-2 Beta Variant

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virus Isolation and Sequencing
2.3. Infection Experiments
2.4. Virus Titration
2.5. RNA Isolation and RT-qPCR
2.6. Histology and Immunohistochemistry
3. Results
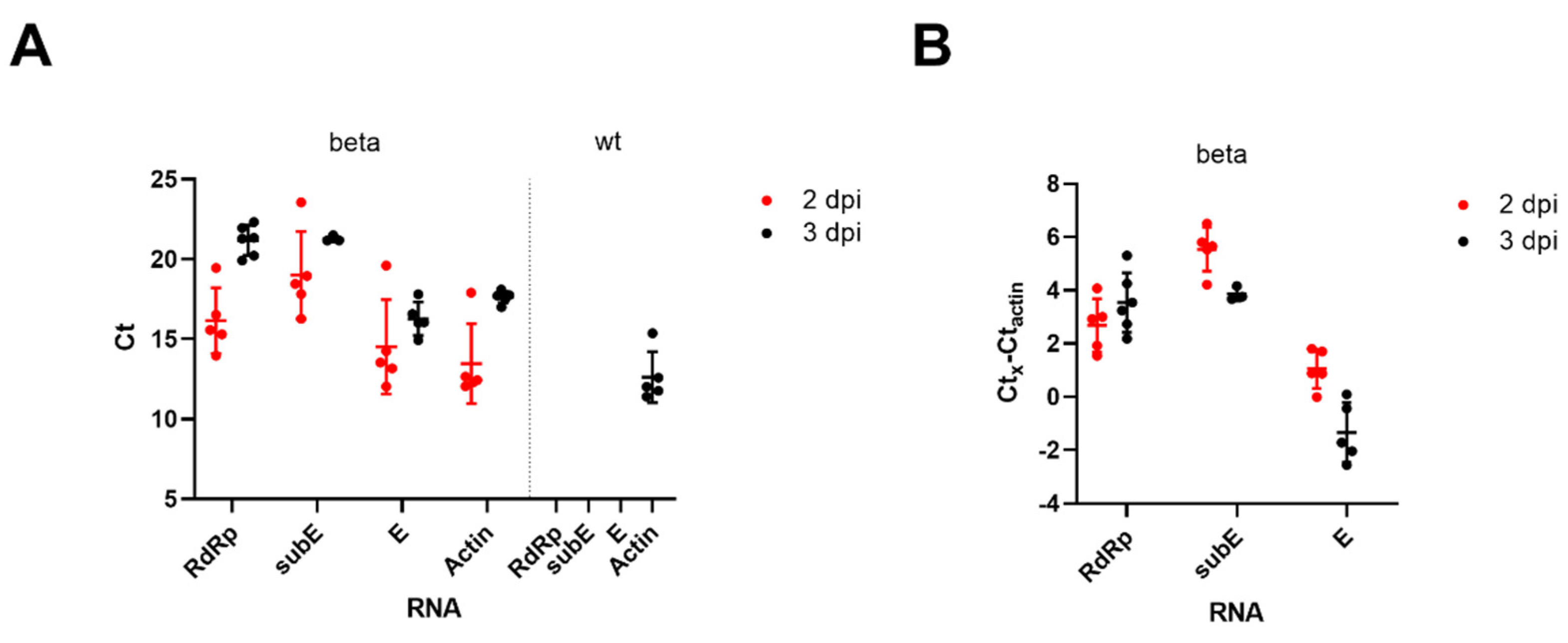
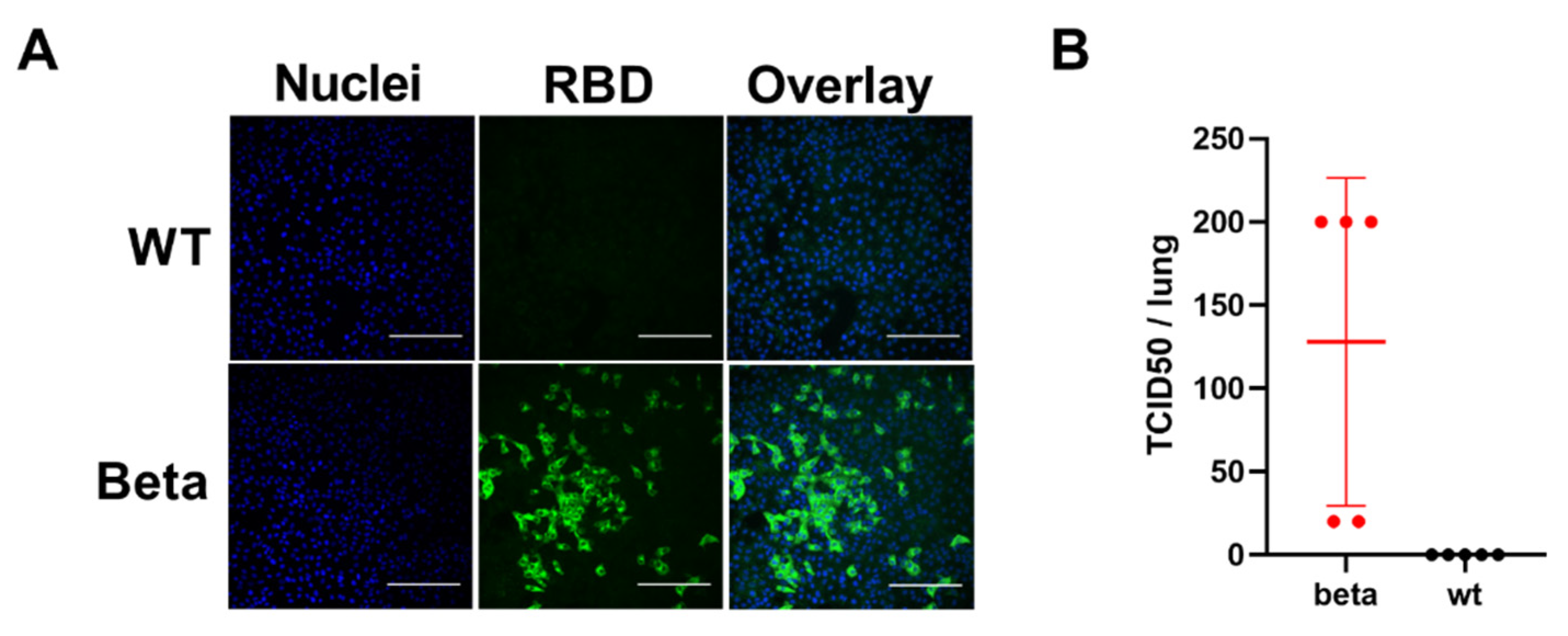
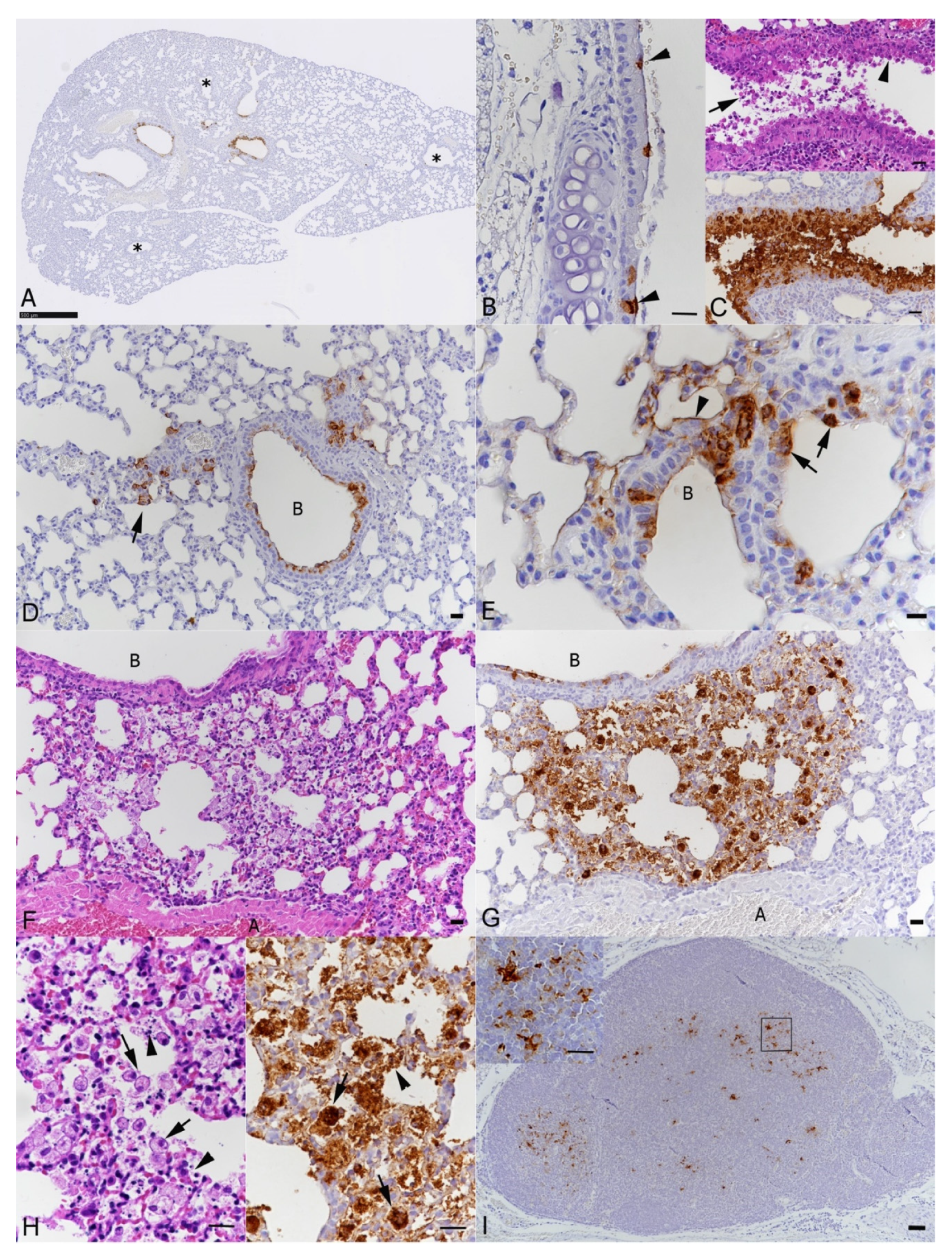
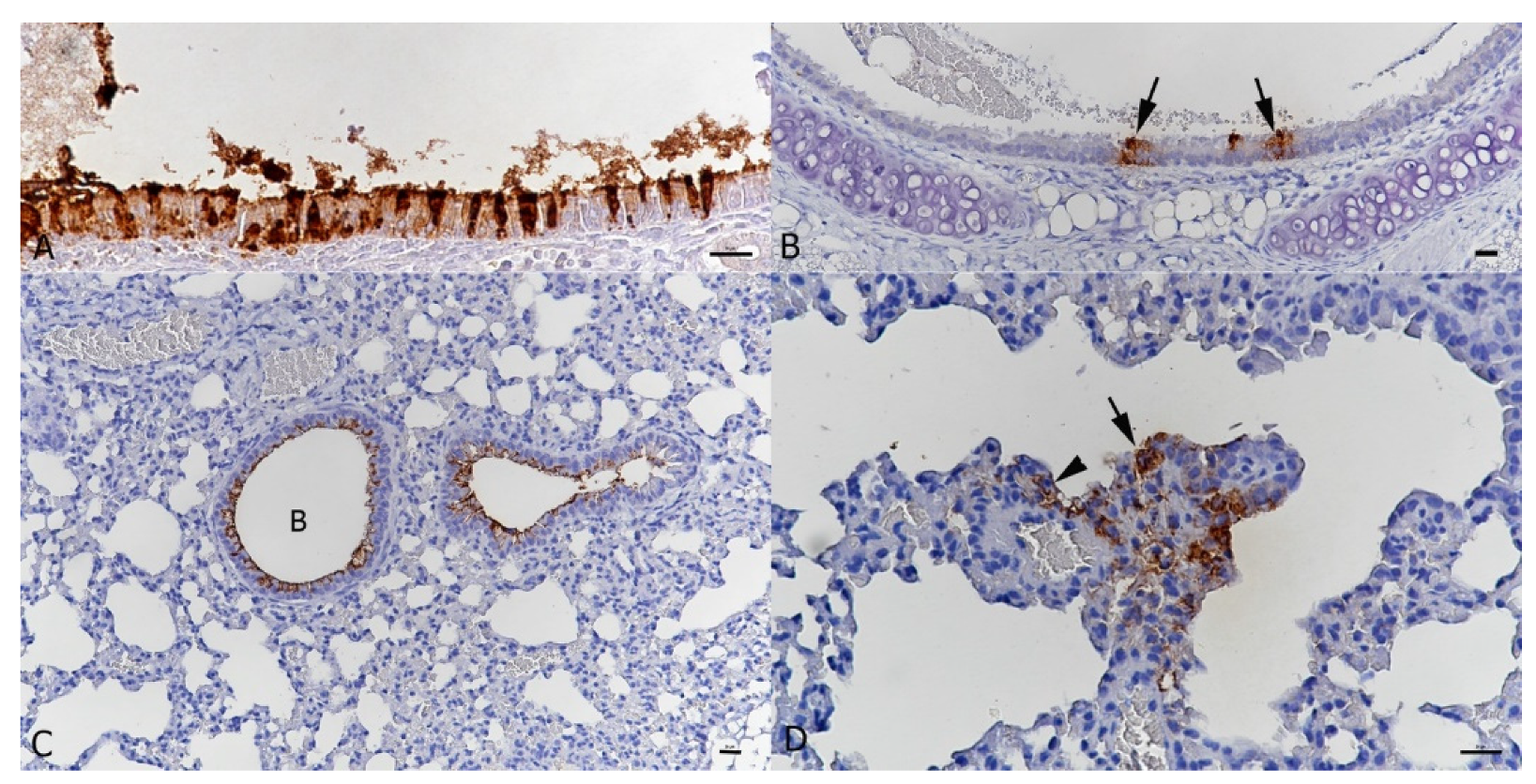
3.1. BALB/c Mice Develop Pulmonary Infection, Alveolar Damage and a Mild Inflammatory Response within 3 Days of Intranasal Challenge with the Beta Variant of SARS-CoV-2
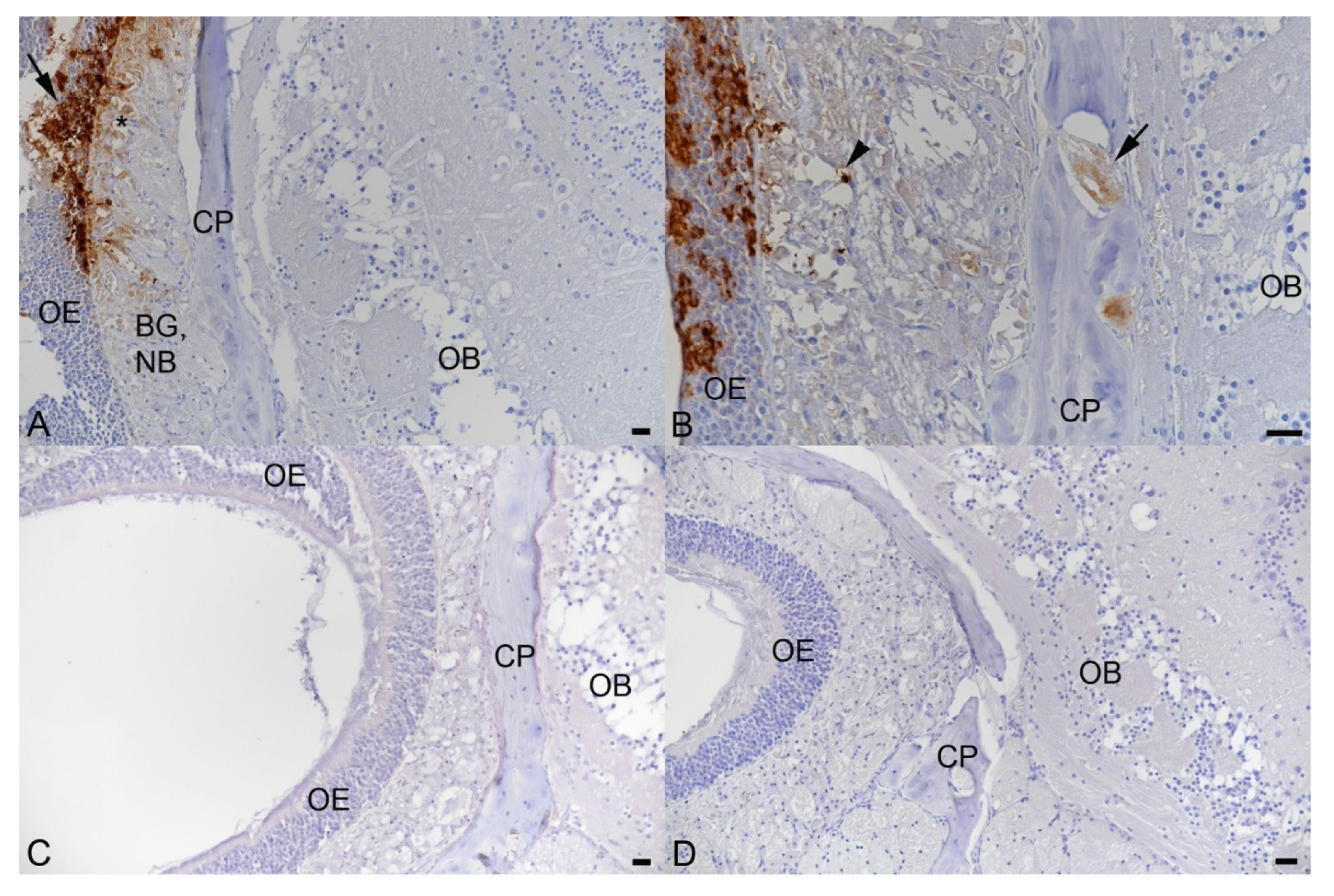
3.2. Intranasal Challenge of BALB/c Mice with a Low Dose of the SARS-CoV-2 Beta Variant Leads to Nasal Infection but Inconsistent Pulmonary Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virtanen, J.; Uusitalo, R.; Korhonen, E.; Aaltonen, K.; Smura, T.; Kuivanen, S.; Pakkanen, S.; Mero, S.; Patjas, A.; Riekkinen, M.; et al. Kinetics of Neutralizing Antibodies of COVID-19 Patients Tested Using Clinical D614G, B.1.1.7, and B 1.351 Isolates in Microneutralization Assays. Viruses 2021, 13, 996. [Google Scholar] [CrossRef]
- Davies, N.G.; Jarvis, C.I.; Edmunds, W.J.; Jewell, N.P.; Diaz-Ordaz, K.; Keogh, R.H. Increased mortality in community-tested cases of SARS-CoV-2 lineage B.1.1.7. Nature 2021, 2021 593, 270–274. [Google Scholar] [CrossRef]
- Konings, F.; Perkins, M.D.; Kuhn, J.H.; Pallen, M.J.; Alm, E.J.; Archer, B.N.; Barakat, A.; Bedford, T.; Bhiman, J.N.; Caly, L.; et al. SARS-CoV-2 Variants of Interest and Concern naming scheme conducive for global discourse. Nat. Microbiol. 2021, 6, 821–823. [Google Scholar] [CrossRef]
- Renn, M.; Bartok, E.; Zillinger, T.; Hartmann, G.; Behrendt, R. Animal models of SARS-CoV-2 and COVID-19 for the development of prophylactic and therapeutic interventions. Pharmacol. Ther. 2021, 228, 107931. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.-S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef]
- Wong, L.Y.R.; Zheng, J.; Wilhelmsen, K.; Li, K.; Ortiz, M.E.; Schnicker, N.J.; Pezzulo, A.A.; Szachowicz, P.J.; Klumpp, K.; Aswad, F.; et al. Eicosanoid signaling as a therapeutic target in middle-aged mice with severe COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Rusanen, J.; Kareinen, L.; Szirovicza, L.; Uğurlu, H.; Levanov, L.; Jääskeläinen, A.; Ahava, M.; Kurkela, S.; Saksela, K.; Hedman, K.; et al. A Generic, Scalable, and Rapid Time-Resolved Förster Resonance Energy Transfer-Based Assay for Antigen Detection-SARS-CoV-2 as a Proof of Concept. Mbio 2021, 12, e00902-21. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed]
- Dagotto, G.; Mercado, N.B.; Martinez, D.R.; Hou, Y.J.; Nkolola, J.P.; Carnahan, R.H.; Crowe, J.E.; Baric, R.S.; Barouch, D.H. Comparison of Subgenomic and Total RNA in SARS-CoV-2 Challenged Rhesus Macaques. J. Virol. 2021, 95, e02370-20. [Google Scholar] [CrossRef]
- Zivcec, M.; Safronetz, D.; Haddock, E.; Feldmann, H.; Ebihara, H. Validation of assays to monitor immune responses in the Syrian golden hamster (Mesocricetus auratus). J. Immunol. Methods 2011, 368, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.J.; Penrice-Randal, R.; Sharma, P.; Kipar, A.; Dong, X.; Pennington, S.H.; Marriott, A.E.; Colombo, S.; Davidson, A.; Williamson, M.K.; et al. Sequential infection with influenza A virus followed by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) leads to more severe disease and encephalitis in a mouse model of COVID-19. bioRxiv 2021. bioRxiv:2020.10.13.334532. [Google Scholar]
- Harvey, W.T.; Carabelli, A.; Jackson, B.; Gupta, R.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Carossino, M.; Montanaro, P.; O’Connell, A.; Kenney, D.; Gertje, H.; Grosz, K.; Kurnick, S.; Bosmann, M.; Saeed, M.; Balasuriya, U.; et al. Fatal neuroinvasion of SARS-CoV-2 in K18-hACE2 mice is partially dependent on hACE2 expression. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yao, W.; Wang, Y.; Ma, D.; Tang, X.; Wang, H.; Li, C.; Lin, H.; Li, Y.; Zhong, G. Effect of SARS-CoV-2 spike mutations on animal ACE2 usage and in vitro neutralization sensitivity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Kuiper, M.J.; Wilson, L.O.W.; Mangalaganesh, S.; Lee, C.; Reti, D.; Vasan, S.S. But Mouse, you are not alone: On some severe acute respiratory syndrome coronavirus 2 variants infecting mice. bioRxiv 2021. [Google Scholar] [CrossRef]
- Montagutelli, X.; Prot, M.; Levillayer, L.; Salazar, E.B.; Jouvion, G.; Conquet, L.; Donati, F.; Albert, M.; Gambaro, F.; Behillil, S.; et al. The B1.351 and P.1 variants extend SARS-CoV-2 host range to mice. bioRxiv 2021. [Google Scholar] [CrossRef]
- Seehusen, F.; Clark, J.J.; Sharma, P.; Subramaniam, K.; Giuliani, S.W.; Hughes, G.; Patterson, E.I.; Michael, B.D.; Owen, A.; Hiscox, J.A.; et al. Viral neuroinvasion and neurotropism without neuronal damage in the hACE2 mouse model of COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence | Ref. | |
|---|---|---|---|
| RdRp | Forward | gtgaratggtcatgtgtggcgg | [11] |
| Probe | caggtggaacctcatcaggagatgc | [11] | |
| Reverse | caratgttaaasacactattagcata | [11] | |
| Subgenomic E | Forward | cgatctcttgtagatctgttctc | [12] |
| Probe | acactagccatccttactgcgcttcg | [12] | |
| Reverse | atattgcagcagtacgcacaca | [12] | |
| Genomic E | Forward | acaggtacgttaatagttaatagcgt | [12] |
| Probe | acac-tagccatccttactgcgcttcg | [12] | |
| Revere | atattgcagcagtacgcacaca | [12] | |
| Beta-actin | Forward | actgccgcatcctcttcct | [13] |
| Probe | cctggagaagagctatgagctgcctgatg | [13] | |
| Reverse | tcgttgccaatggtgatgac | [13] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kant, R.; Kareinen, L.; Smura, T.; Freitag, T.L.; Jha, S.K.; Alitalo, K.; Meri, S.; Sironen, T.; Saksela, K.; Strandin, T.; et al. Common Laboratory Mice Are Susceptible to Infection with the SARS-CoV-2 Beta Variant. Viruses 2021, 13, 2263. https://doi.org/10.3390/v13112263
Kant R, Kareinen L, Smura T, Freitag TL, Jha SK, Alitalo K, Meri S, Sironen T, Saksela K, Strandin T, et al. Common Laboratory Mice Are Susceptible to Infection with the SARS-CoV-2 Beta Variant. Viruses. 2021; 13(11):2263. https://doi.org/10.3390/v13112263
Chicago/Turabian StyleKant, Ravi, Lauri Kareinen, Teemu Smura, Tobias L. Freitag, Sawan Kumar Jha, Kari Alitalo, Seppo Meri, Tarja Sironen, Kalle Saksela, Tomas Strandin, and et al. 2021. "Common Laboratory Mice Are Susceptible to Infection with the SARS-CoV-2 Beta Variant" Viruses 13, no. 11: 2263. https://doi.org/10.3390/v13112263