
Severe Drought Still Affects Reproductive Traits Two Years Later in a Common Garden Experiment of Frangula alnus
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Garden
2.2. Measurements
2.3. Statistical Analysis
3. Results
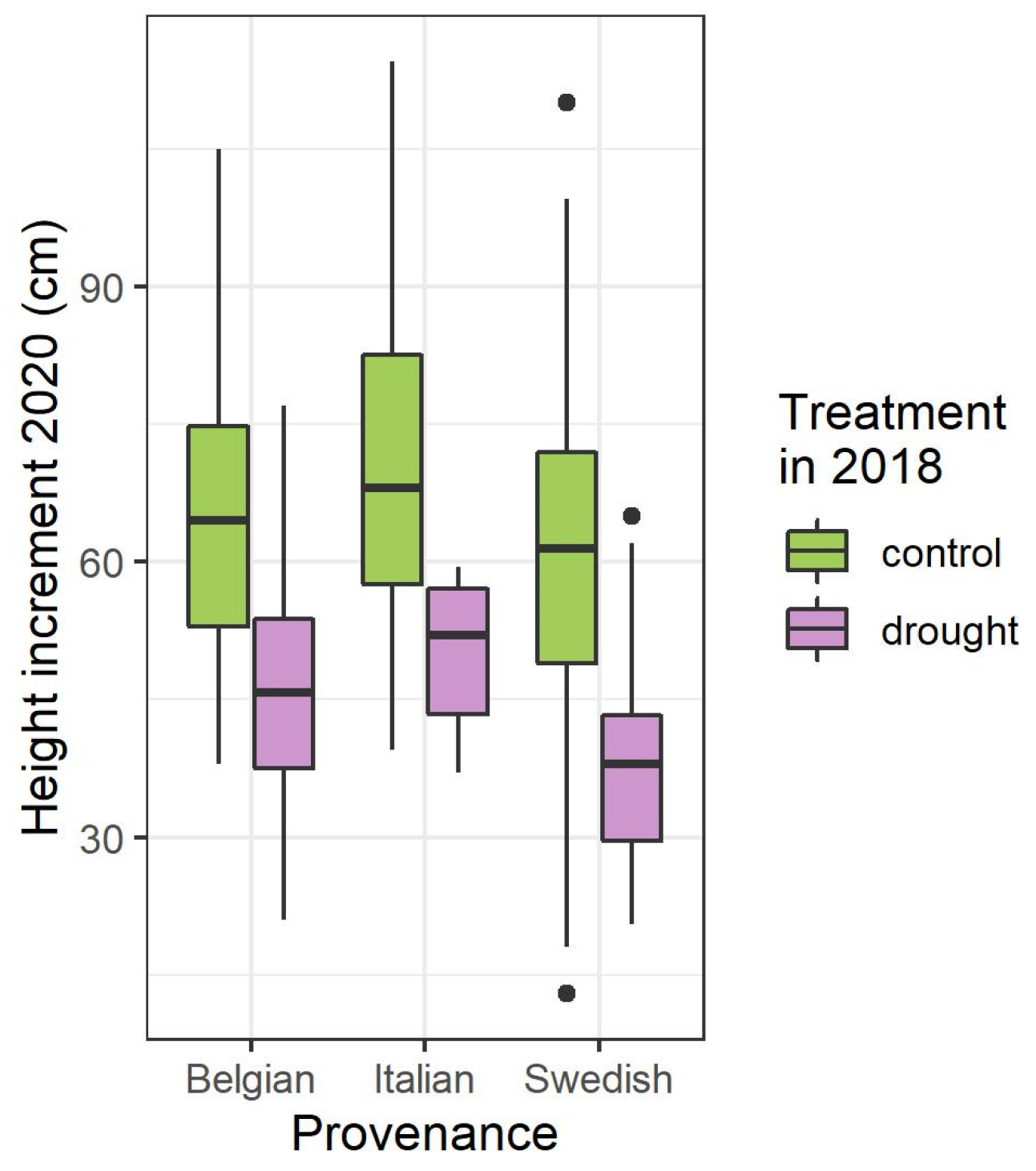
3.1. Height Increment, Two Years after the Summer Drought Treatment
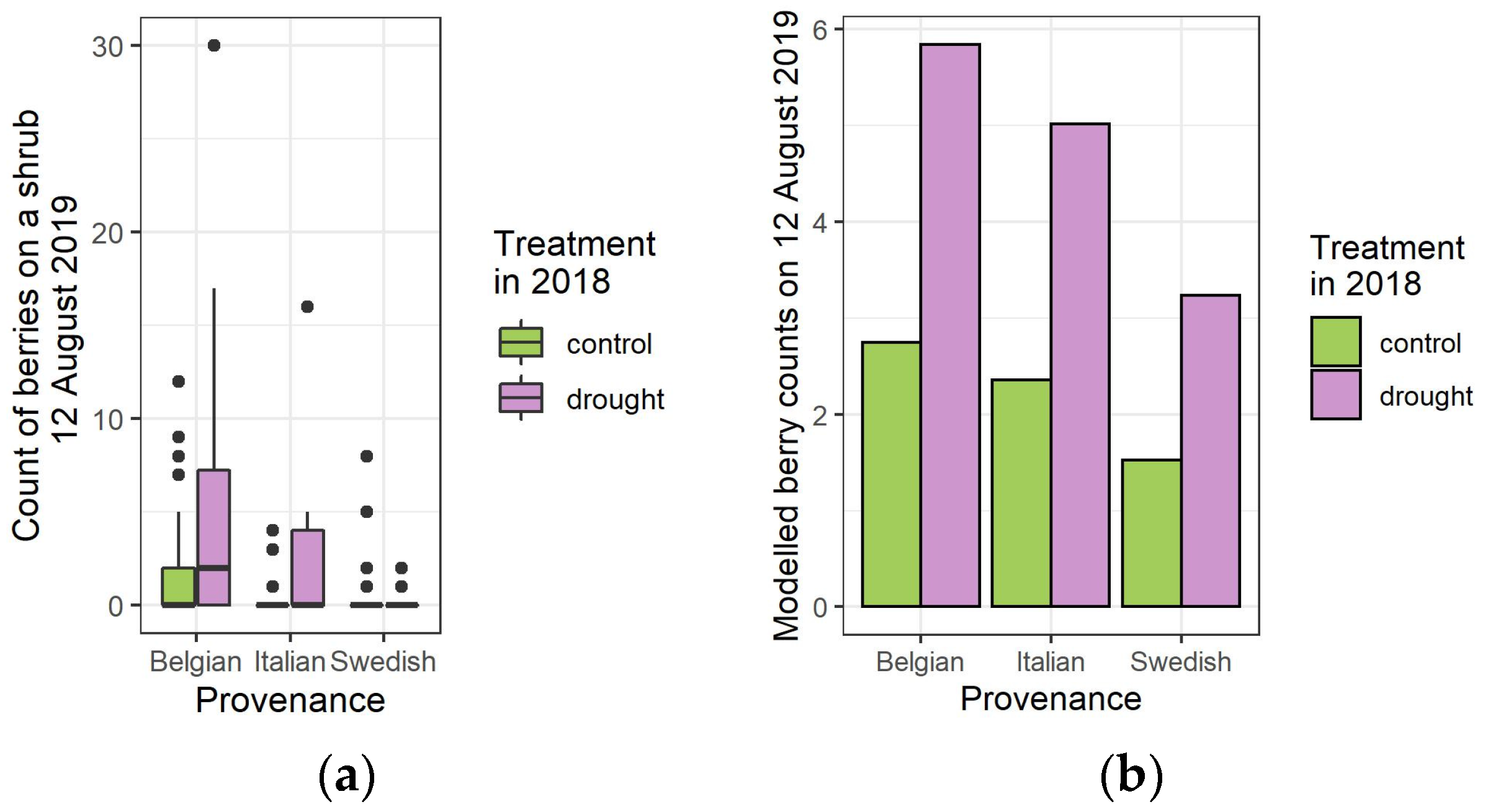
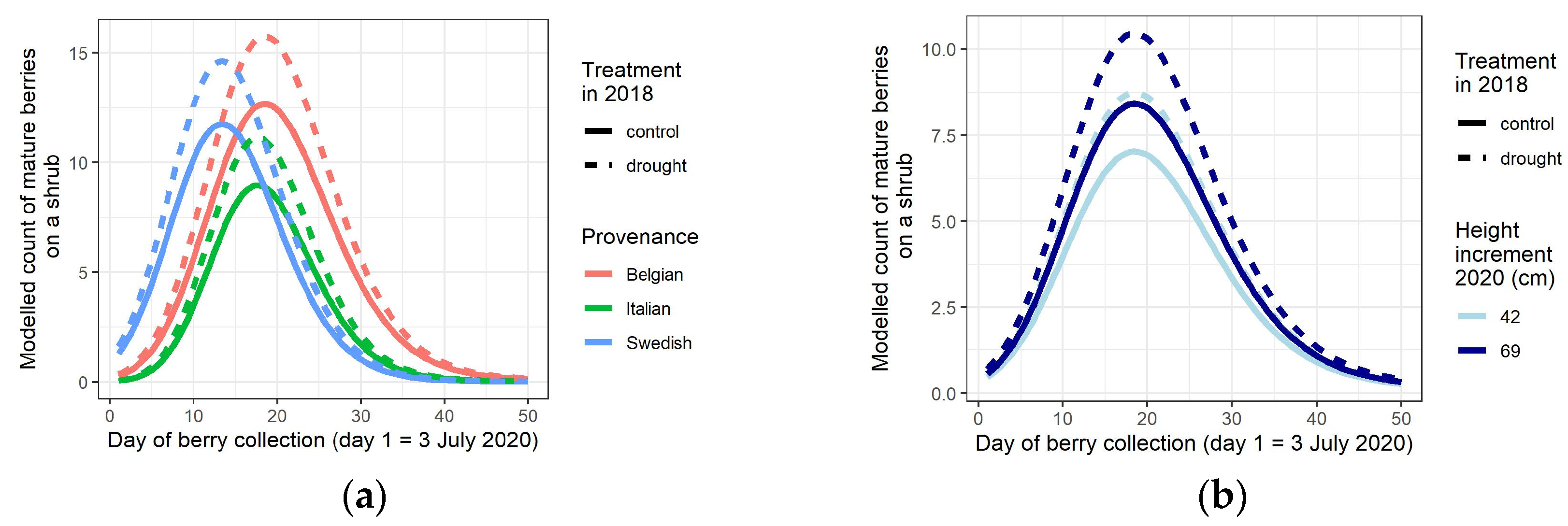
3.2. Mature Berry Counts, One and Two Years after the Drought Treatment
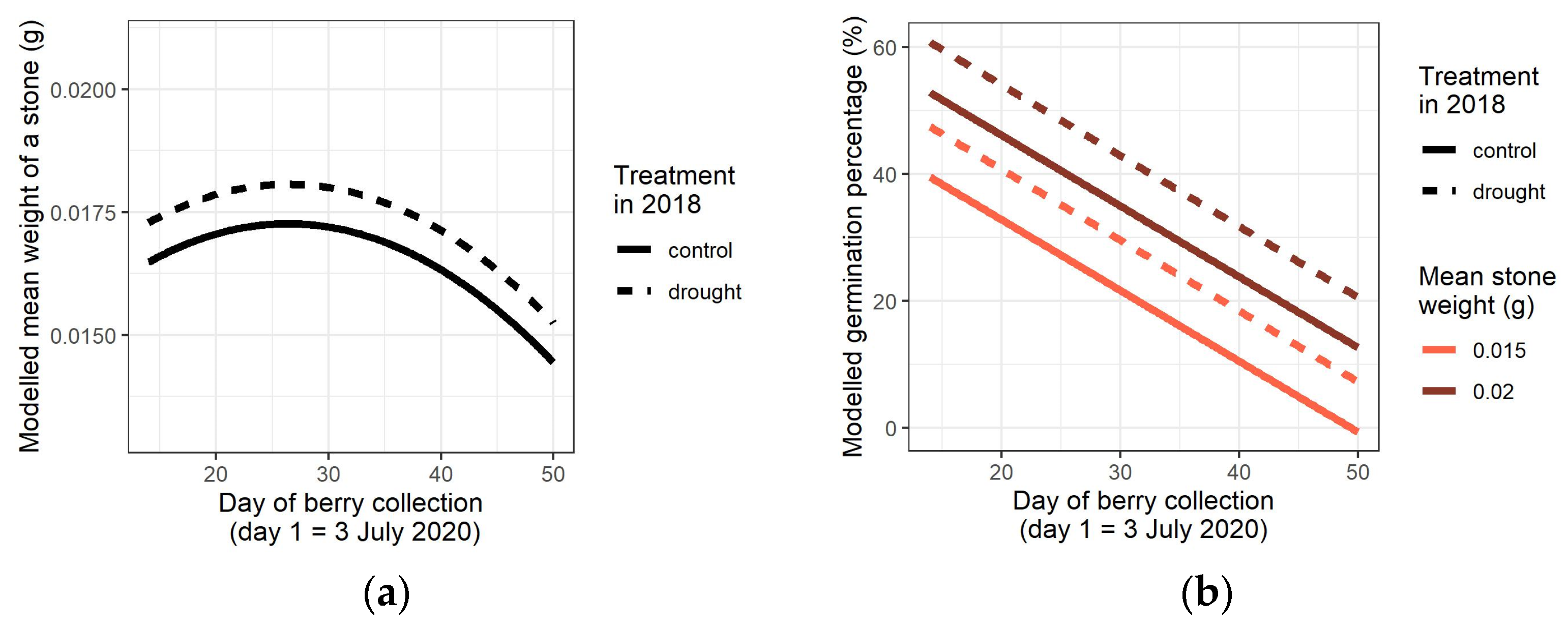
3.3. Mean Stone Weight
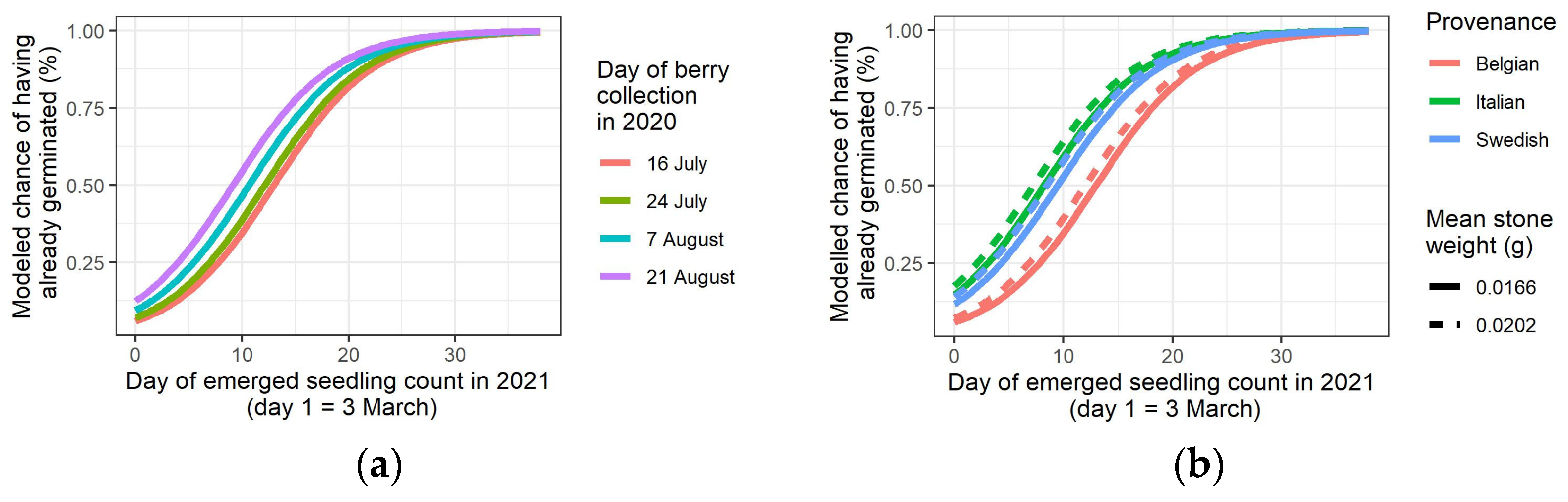
3.4. Germination of the Stones in the Spring of 2021
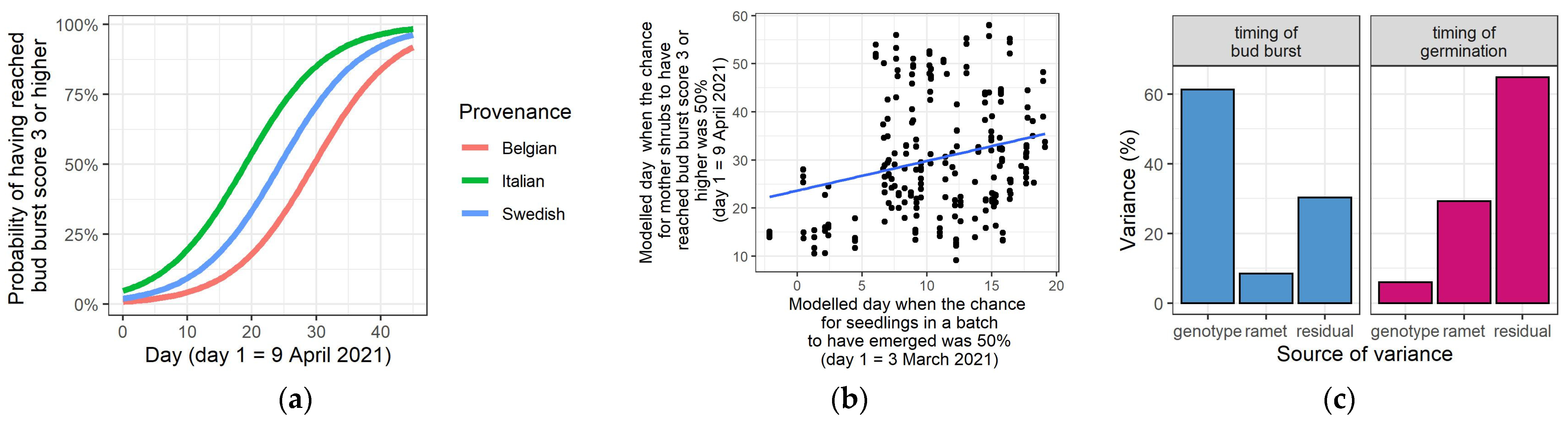
3.5. Timing of Bud Burst and Seedling Emergence
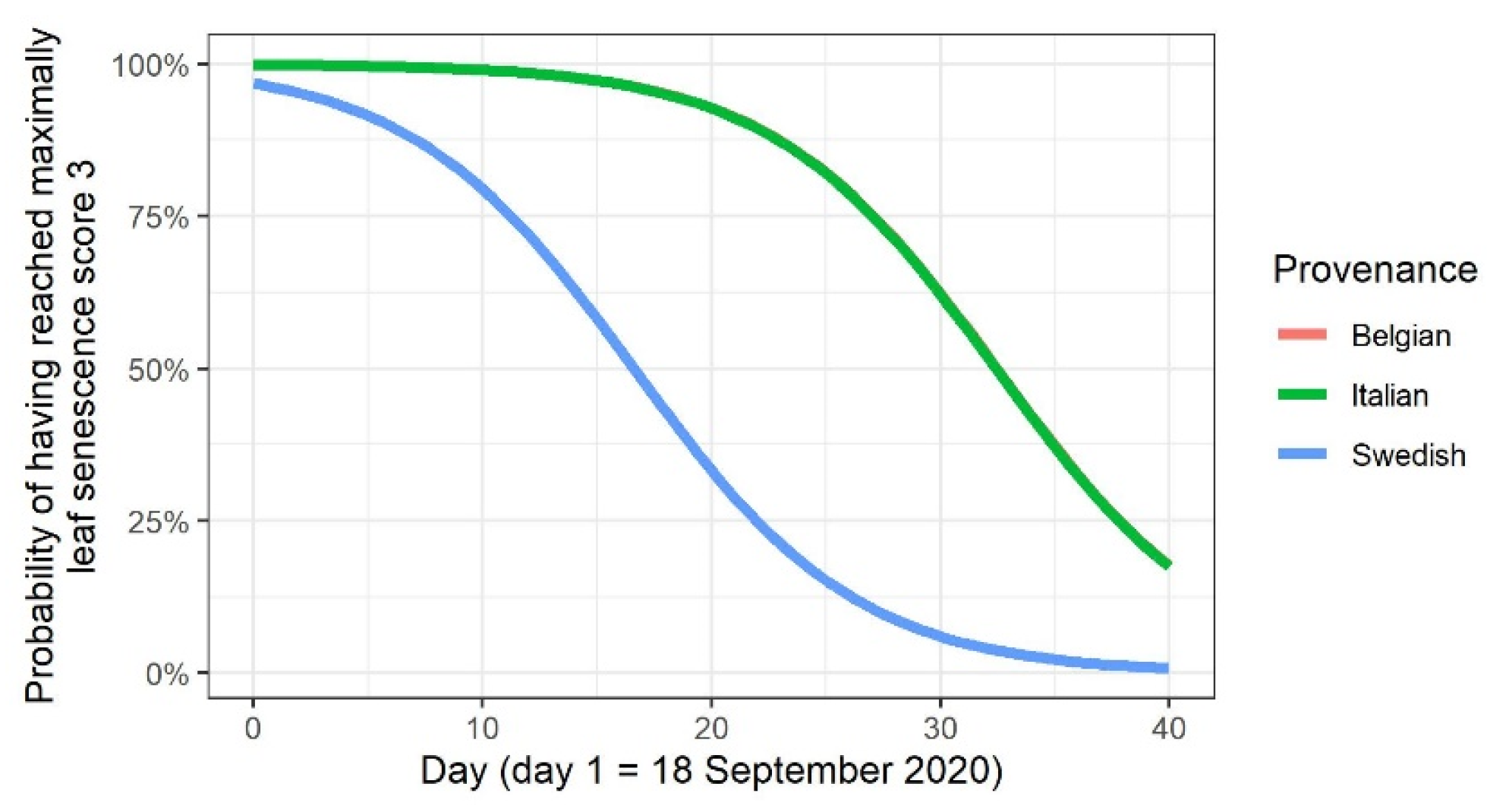
3.6. Leaf Senescence
4. Discussion
4.1. Influence of Former Water Limitation on Growth and Reproduction
4.2. Population Differentiation in the Common Garden
4.3. Phenology of Berry Maturation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant increase in natural disturbance impacts on European forests since 1950. Glob. Chang. Biol. 2022, 29, 1359–1376. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Kelly, D.; Thomas, P.A.; Lageard, J.G.; Hacket-Pain, A. Climate warming disrupts mast seeding and its fitness benefits in European beech. Nat. Plants 2020, 6, 88–94. [Google Scholar] [CrossRef]
- Monks, A.; Monks, J.M.; Tanentzap, A.J. Resource limitation underlying multiple masting models makes mast seeding sensitive to future climate change. New Phytol. 2016, 210, 419–430. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Ascoli, D.; Hacket-Pain, A.; Koenig, W.D.; Pearse, I.; Pesendorfer, M.; Satake, A.; Thomas, P.; Vacchiano, G.; Wohlgemuth, T.; et al. From theory to experiments for testing the proximate mechanisms of mast seeding: An agenda for an experimental ecology. Ecol. Lett. 2020, 23, 210–220. [Google Scholar] [CrossRef]
- Richardson, S.J.; Allen, R.B.; Whitehead, D.; Carswell, F.E.; Ruscoe, W.A.; Platt, K.H. Climate and Net Carbon Availability Determine Temporal Patterns of Seed Production by Nothofagus. Ecology 2005, 86, 972–981. [Google Scholar] [CrossRef]
- Redmond, M.D.; Forcella, F.; Barger, N.N. Declines in pinyon pine cone production associated with regional warming. Ecosphere 2012, 3, 1–14. [Google Scholar] [CrossRef]
- Allen, R.B.; Hurst, J.M.; Portier, J.; Richardson, S.J. Elevation-dependent responses of tree mast seeding to climate change over 45 years. Ecol. Evol. 2014, 4, 3525–3537. [Google Scholar] [CrossRef]
- Pesendorfer, M.B.; Bogdziewicz, M.; Szymkowiak, J.; Borowski, Z.; Kantorowicz, W.; Espelta, J.M.; Fernández-Martínez, M. Investigating the relationship between climate, stand age, and temporal trends in masting behavior of European forest trees. Glob. Chang. Biol. 2020, 26, 1654–1667. [Google Scholar] [CrossRef]
- Leuzinger, S.; Luo, Y.; Beier, C.; Dieleman, W.; Vicca, S.; Körner, C. Do global change experiments overestimate impacts on terrestrial ecosystems? Trends Ecol. Evol. 2011, 26, 236–241. [Google Scholar] [CrossRef]
- Herman, J.; Sultan, S. Adaptive Transgenerational Plasticity in Plants: Case Studies, Mechanisms, and Implications for Natural Populations. Front. Plant Sci. 2011, 2, 102. [Google Scholar] [CrossRef]
- Galloway, L.F.; Etterson, J.R. Transgenerational Plasticity Is Adaptive in the Wild. Science 2007, 318, 1134–1136. [Google Scholar] [CrossRef]
- Zecchin, B.; Caudullo, G.; de Rigo, D. Frangula alnus in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Vander Mijnsbrugge, K.; De Clerck, L.; Van der Schueren, N.; Moreels, S.; Lauwers, A.; Steppe, K.; De Ligne, L.; Campioli, M.; Van den Bulcke, J. Counter-Intuitive Response to Water Limitation in a Southern European Provenance of Frangula alnus Mill. in a Common Garden Experiment. Forests 2020, 11, 1186. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Schouppe, M.; Moreels, S.; Aguas Guerreiro, Y.; Decorte, L.; Stessens, M. Influence of Water Limitation and Provenance on Reproductive Traits in a Common Garden of Frangula alnus Mill. Forests 2022, 13, 1744. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Schouppe, M.; Moreels, S.; De Leenheer, S. Transgenerational Effects of Water Limitation on Reproductive Mother Plants in a Common Garden of the Shrub Frangula alnus. Forests 2023, 14, 348. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 1 February 2020).
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Christensen, R.H.B. Ordinal: Regression Models for Ordinal Data, R Package Version 2015.6-28; R Foundation for Statistical Computing: Vienna, Austria, 2015. Available online: http://www.cran.r-project.org/package=ordinal/(accessed on 1 February 2020).
- Hoch, G.; Siegwolf, R.T.W.; Keel, S.G.; Körner, C.; Han, Q. Fruit production in three masting tree species does not rely on stored carbon reserves. Oecologia 2013, 171, 653–662. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Fernández-Martínez, M.; Espelta, J.M.; Ogaya, R.; Penuelas, J. Is forest fecundity resistant to drought? Results from an 18-yr rainfall-reduction experiment. New Phytol. 2020, 227, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Vergotti, M.J.; Fernández-Martínez, M.; Kefauver, S.C.; Janssens, I.A.; Peñuelas, J. Weather and trade-offs between growth and reproduction regulate fruit production in European forests. Agric. For. Meteorol. 2019, 279, 107711. [Google Scholar] [CrossRef]
- Caignard, T.; Kremer, A.; Firmat, C.; Nicolas, M.; Venner, S.; Delzon, S. Increasing spring temperatures favor oak seed production in temperate areas. Sci. Rep. 2017, 7, 8555. [Google Scholar] [CrossRef] [PubMed]
- Hacket-Pain, A.J.; Lageard, J.G.A.; Thomas, P.A. Drought and reproductive effort interact to control growth of a temperate broadleaved tree species (Fagus sylvatica). Tree Physiol. 2017, 37, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Lauder, J.D.; Moran, E.V.; Hart, S.C. Fight or flight? Potential tradeoffs between drought defense and reproduction in conifers. Tree Physiol. 2019, 39, 1071–1085. [Google Scholar] [CrossRef]
- Takeno, K. Stress-induced flowering: The third category of flowering response. J. Exp. Bot. 2016, 67, 4925–4934. [Google Scholar] [CrossRef]
- Zwolak, R.; Celebias, P.; Bogdziewicz, M. Global patterns in the predator satiation effect of masting: A meta-analysis. Proc. Natl. Acad. Sci. USA 2022, 119, e2105655119. [Google Scholar] [CrossRef]
- Bogdziewicz, M. How will global change affect plant reproduction? A framework for mast seeding trends. New Phytol. 2022, 234, 14–20. [Google Scholar] [CrossRef]
- Piovesan, G.; Adams, J.M. Masting behaviour in beech: Linking reproduction and climatic variation. Can. J. Bot. 2001, 79, 1039–1047. [Google Scholar] [CrossRef]
- Davi, H.; Cailleret, M.; Restoux, G.; Amm, A.; Pichot, C.; Fady, B. Disentangling the factors driving tree reproduction. Ecosphere 2016, 7, e01389. [Google Scholar] [CrossRef]
- Mojzes, A.; Kalapos, T.; Kröel-Dulay, G. Drought in maternal environment boosts offspring performance in a subordinate annual grass. Environ. Exp. Bot. 2021, 187, 104472. [Google Scholar] [CrossRef]
- Germain, R.M.; Caruso, C.M.; Maherali, H. Mechanisms and Consequences of Water Stress–Induced Parental Effects in an Invasive Annual Grass. Int. J. Plant Sci. 2013, 174, 886–895. [Google Scholar] [CrossRef]
- Tielbörger, K.; Valleriani, A. Can seeds predict their future? Germination strategies of density-regulated desert annuals. Oikos 2005, 111, 235–244. [Google Scholar] [CrossRef]
- Stahl, U.; Kattge, J.; Reu, B.; Voigt, W.; Ogle, K.; Dickie, J.; Wirth, C. Whole-plant trait spectra of North American woody plant species reflect fundamental ecological strategies. Ecosphere 2013, 4, art128. [Google Scholar] [CrossRef]
- Leishman, M.R.; Westoby, M. The Role of Seed Size in Seedling Establishment in Dry Soil Conditions—Experimental Evidence from Semi-Arid Species. J. Ecol. 1994, 82, 249–258. [Google Scholar] [CrossRef]
- Rice, K.J.; Knapp, E.E. Effects of Competition and Life History Stage on the Expression of Local Adaptation in Two Native Bunchgrasses. Restor. Ecol. 2008, 16, 12–23. [Google Scholar] [CrossRef]
- Hamann, E.; Kesselring, H.; Armbruster, G.F.J.; Scheepens, J.F.; Stöcklin, J. Evidence of local adaptation to fine- and coarse-grained environmental variability in Poa alpina in the Swiss Alps. J. Ecol. 2016, 104, 1627–1637. [Google Scholar] [CrossRef]
- Schippers, J.H.M.; Schmidt, R.; Wagstaff, C.; Jing, H.-C. Living to Die and Dying to Live: The Survival Strategy behind Leaf Senescence. Plant Physiol. 2015, 169, 914–930. [Google Scholar] [CrossRef]
- Rosecrance, R.C.; Weinbaum, S.A.; Brown, P.H. Alternate Bearing Affects Nitrogen, Phosphorus, Potassium and Starch Storage Pools in Mature Pistachio Trees. Ann. Bot. 1998, 82, 463–470. [Google Scholar] [CrossRef]
- Rohde, A.; Bhalerao, R. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Mehlenbacher, S.A.; Voordeckers, A.M. Relationship of flowering time, rate of seed germination, and time of leaf budbreak and usefulness in selecting for late-flowering apples. J. Am. Soc. Hortic. Sci. 1991, 116, 565–568. [Google Scholar] [CrossRef]
- Vahdati, K.; Aslani Aslamarz, A.; Rahemi, M.; Hassani, D.; Leslie, C. Mechanism of seed dormancy and its relationship to bud dormancy in Persian walnut. Environ. Exp. Bot. 2012, 75, 74–82. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Moreels, S. Varying Levels of Genetic Control and Phenotypic Plasticity in Timing of Bud Burst, Flower Opening, Leaf Senescence and Leaf Fall in Two Common Gardens of Prunus padus L. Forests 2020, 11, 1070. [Google Scholar] [CrossRef]
- Soularue, J.-P.; Kremer, A. Assortative mating and gene flow generate clinal phenological variation in trees. BMC Evol. Biol. 2012, 12, 79. [Google Scholar] [CrossRef]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene Flow and Local Adaptation in Trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Muffler, L.; Schmeddes, J.; Weigel, R.; Barbeta, A.; Beil, I.; Bolte, A.; Buhk, C.; Holm, S.; Klein, G.; Klisz, M.; et al. High plasticity in germination and establishment success in the dominant forest tree Fagus sylvatica across Europe. Glob. Ecol. Biogeogr. 2021, 30, 1583–1596. [Google Scholar] [CrossRef]
- Arana, M.V.; Gonzalez-Polo, M.; Martinez-Meier, A.; Gallo, L.A.; Benech-Arnold, R.L.; Sánchez, R.A.; Batlla, D. Seed dormancy responses to temperature relate to Nothofagus species distribution and determine temporal patterns of germination across altitudes in Patagonia. New Phytol. 2016, 209, 507–520. [Google Scholar] [CrossRef]
- Rousi, M.; Heinonen, J.; Neuvonen, S. Intrapopulation variation in flowering phenology and fecundity of silver birch, implications for adaptability to changing climate. For. Ecol. Manag. 2011, 262, 2378–2385. [Google Scholar] [CrossRef]
- Pauwels, E.; Eyssen, R.; Keulemans, J. Seed dormancy and bud break of apple seedlings. Acta Hortic. 1998, 484, 119–122. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Berry Collection Day | Belgian | Italian | Swedish | Total | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Drought | Control | Drought | Control | Drought | Control | Drought | |||||||
| nb° | ns° | nb° | ns° | nb° | ns° | nb° | ns° | nb° | ns° | nb° | ns° | ns° | ns° | |
| 16 July | 17 | 1214 | 16 | 954 | 7 | 442 | 5 | 118 | 14 | 648 | 13 | 564 | 2304 | 1636 |
| 24 July | 17 | 724 | 14 | 729 | 8 | 234 | 4 | 47 | 13 | 445 | 8 | 119 | 1403 | 895 |
| 7 August | 17 | 462 | 16 | 332 | 6 | 101 | 3 | 30 | 7 | 115 | 3 | 12 | 678 | 374 |
| 21 August | 9 | 52 | 12 | 42 | 2 | 4 | 1 | 2 | 1 | 3 | 5 | 13 | 59 | 57 |
| Variable | Estimate | Std. Error | df | t-Value | p-Value |
|---|---|---|---|---|---|
| (Intercept) | 64.948 | 2.404 | 196 | 27.021 | <0.001 *** |
| It | 4.528 | 4.625 | 33 | 0.979 | 0.335 |
| Sw | −6.117 | 3.480 | 33 | −1.758 | 0.088 |
| T | −9.095 | 1.830 | 196 | −10.434 | <0.001 *** |
| Year | Variable | Estimate | Std. Error | z-Value | p-Value |
|---|---|---|---|---|---|
| 2019 | (Intercept) | 0.840 | 0.377 | 2.23 | 0.026 * |
| It | −0.153 | 0.391 | −0.392 | 0.695 | |
| Sw | −0.592 | 0.370 | −1.600 | 0.110 | |
| T | 0.754 | 0.123 | 6.119 | <0.001 *** | |
| H19 | 0.002 | 0.004 | 0.540 | 0.589 | |
| 2020 | (Intercept) | 0.45 | 0.26 | 1.76 | 0.079 |
| It | −0.97 | 0.31 | −3.13 | 0.002 ** | |
| Sw | −0.83 | 0.22 | −3.74 | <0.001 *** | |
| T | 0.22 | 0.11 | 2.05 | 0.040 * | |
| H20 | 0.01 | 0.00 | 2.03 | 0.042 * | |
| C | −18.67 | 1.45 | −12.89 | <0.001 *** | |
| C2 | −28.35 | 1.09 | −26.08 | <0.001 *** | |
| C3 | 9.08 | 0.95 | 9.58 | <0.001 *** | |
| C:It | −7.62 | 4.95 | −1.54 | 0.123 | |
| C2:It | −6.55 | 3.47 | −1.89 | 0.059 | |
| C3:It | 5.77 | 2.94 | 1.97 | 0.049 * | |
| C:Sw | −29.60 | 3.98 | −7.44 | <0.001 *** | |
| C2:Sw | 5.50 | 2.78 | 1.98 | 0.048 * | |
| C3:Sw | 1.56 | 1.95 | 0.80 | 0.423 |
| Variable | Estimate | Std. Error | df | t-Value | p-Value |
|---|---|---|---|---|---|
| (Intercept) | 0.0166 | 0.0006 | 131 | 28.15 | <0.001 *** |
| It | 0.0018 | 0.0012 | 31 | 1.56 | 0.128 |
| Sw | 0.0008 | 0.0009 | 31 | 0.96 | 0.343 |
| T | 0.0008 | 0.0004 | 34 | 2.25 | 0.031 * |
| C | −0.0205 | 0.0022 | 131 | −9.27 | <0.001 *** |
| C2 | −0.0172 | 0.0022 | 131 | −7.84 | <0.001 *** |
| Germination Percentage | Timing of Seedling Emergence | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Estimate | Std. Error | df | t-Value | p-Value | Estimate | Std. Error | z-Value | p-Value |
| (Intercept) | 15.22 | 9.39 | 128 | 1.62 | 0.107 | −4.04 | 0.46 | −8.80 | <0.001 *** |
| It | −8.02 | 5.20 | 31 | −1.54 | 0.133 | 1.01 | 0.28 | 3.60 | <0.001 *** |
| Sw | −1.11 | 3.85 | 31 | −0.29 | 0.775 | 0.75 | 0.20 | 3.73 | <0.001 *** |
| T | 7.93 | 2.44 | 33 | 3.25 | 0.003 ** | −0.24 | 0.17 | −1.44 | 0.151 |
| S | 2661.75 | 434.73 | 128 | 6.12 | <0.001 *** | 57.89 | 21.57 | 2.68 | 0.007 ** |
| C | −1.12 | 0.11 | 128 | −9.86 | <0.001 *** | 0.02 | 0.01 | 2.24 | 0.025 * |
| D | 0.21 | 0.00 | 90.95 | <0.001 *** | |||||
| Timing of Bud Burst on Mother Shrubs | Timing of Seedling Emergence | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Estimate | Std. Error | z-Value | p-Value | Estimate | Std. Error | z-Value | p-Value |
| It | −1.68 | 0.55 | −3.03 | 0.002 ** | 0.87 | 0.36 | 2.42 | 0.016 * |
| Sw | −0.83 | 0.46 | −1.81 | 0.070 | 0.71 | 0.28 | 2.58 | 0.010 * |
| H20/S | 0.04 | 0.01 | 6.41 | <0.001 *** | 100.33 | 60.47 | 1.66 | 0.097 |
| D | −0.16 | 0.01 | −29.36 | <0.001 *** | 0.19 | 0.00 | 70.59 | <0.001 *** |
| Variable | Estimate | Std. Error | z-Value | p-Value |
|---|---|---|---|---|
| It | 0.009 | 0.455 | 0.019 | 0.984 |
| Sw | 3.244 | 0.401 | 8.088 | <0.001 *** |
| T | −0.049 | 0.210 | −0.236 | 0.813 |
| H20 | 0.023 | 0.006 | 3.747 | <0.001 *** |
| D | 0.205 | 0.010 | 20.287 | <0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vander Mijnsbrugge, K.; Schouppe, M.; Moreels, S.; Aguas Guerreiro, Y.; Decorte, L.; Stessens, M. Severe Drought Still Affects Reproductive Traits Two Years Later in a Common Garden Experiment of Frangula alnus. Forests 2023, 14, 857. https://doi.org/10.3390/f14040857
Vander Mijnsbrugge K, Schouppe M, Moreels S, Aguas Guerreiro Y, Decorte L, Stessens M. Severe Drought Still Affects Reproductive Traits Two Years Later in a Common Garden Experiment of Frangula alnus. Forests. 2023; 14(4):857. https://doi.org/10.3390/f14040857
Chicago/Turabian StyleVander Mijnsbrugge, Kristine, Marc Schouppe, Stefaan Moreels, Yorrick Aguas Guerreiro, Laura Decorte, and Marie Stessens. 2023. "Severe Drought Still Affects Reproductive Traits Two Years Later in a Common Garden Experiment of Frangula alnus" Forests 14, no. 4: 857. https://doi.org/10.3390/f14040857