
Influence of Different Planting Combinations on the Amino Acid Concentration in Pericarp of Zanthoxylum planispinum ‘Dintanensis’ and Soil

Abstract
:1. Introduction
2. Materials and Methods
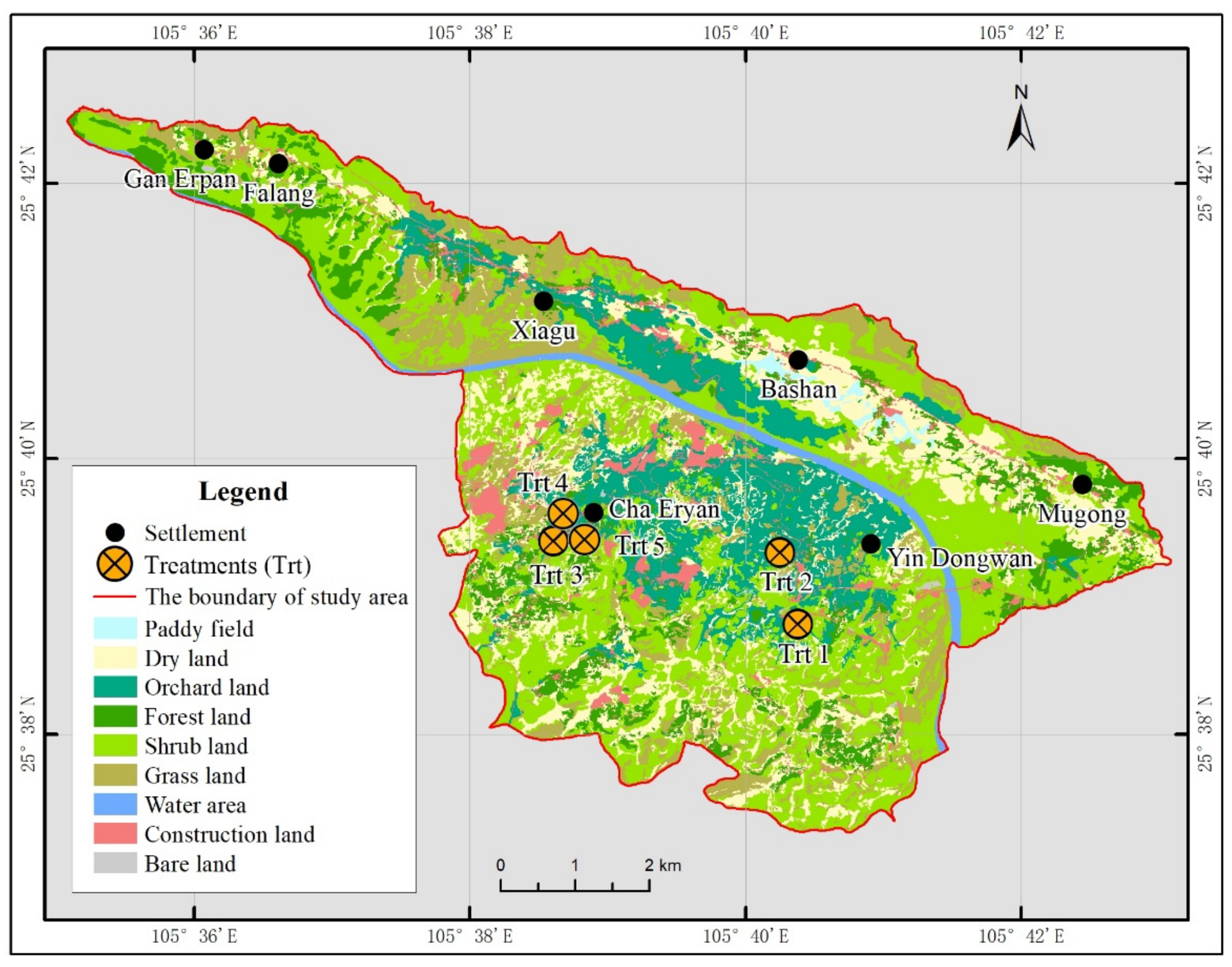
2.1. Overview of the Research Site
2.2. Treatment Setting
2.3. Soil Sample Collection and Soil Parameters
2.4. Fruit Sample Collection
2.5. Free Amino Acid Analysis
2.6. Data Analysis
3. Results
3.1. Amino Acid Concentrations in the Pericarp of Z. planispinum in Different Planting Combinations
3.2. Accumulation of Flavoring Amino Acids in Pericarp of Z. planispinum in Different Planting Combinations
3.3. Principal Component Analysis of Free Amino Acids in Pericarp of Z. planispinum
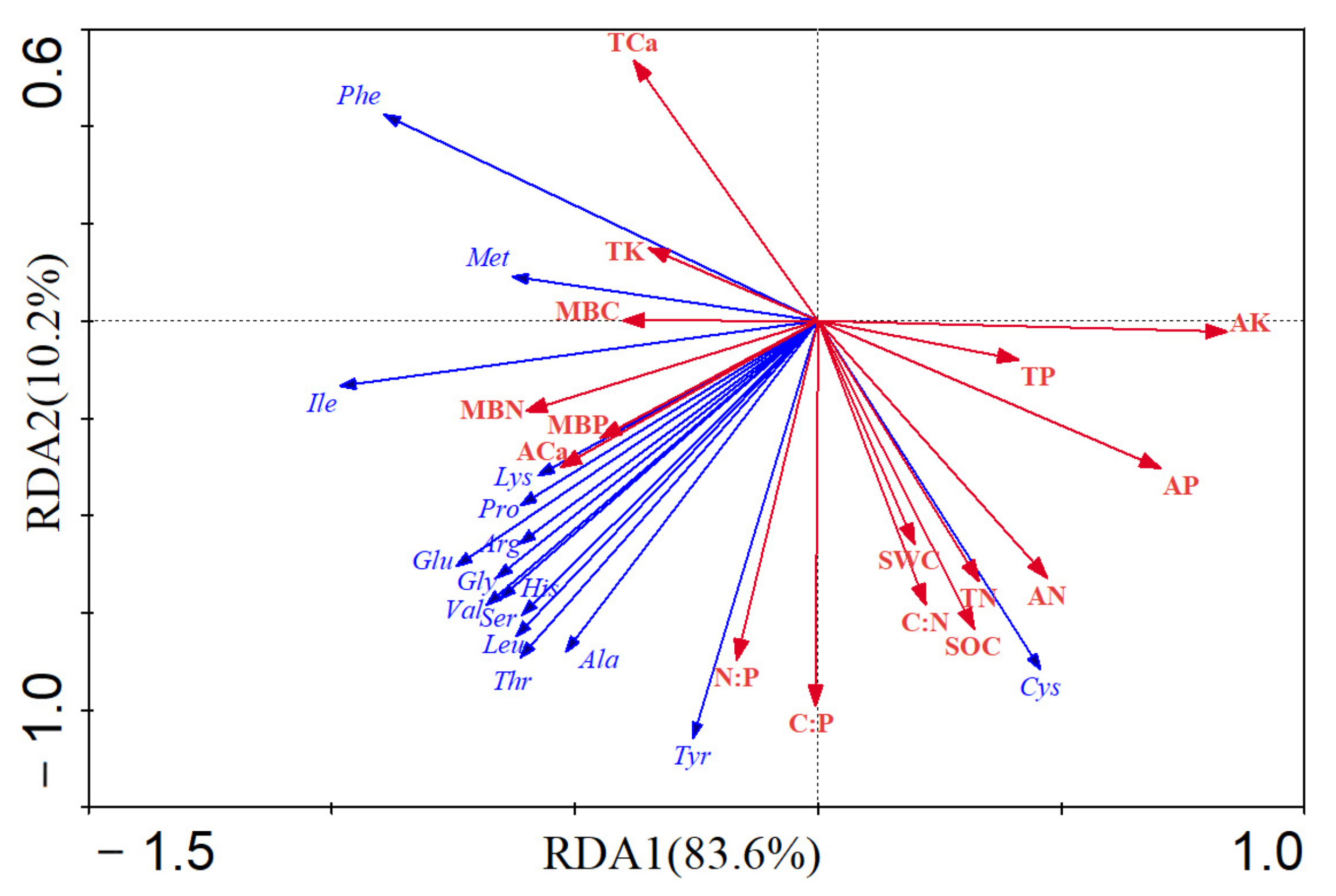
3.4. Effects of Soil Properties on Amino Acid Concentration of Pericarp
4. Discussion
4.1. Effects of Planting Combinations on Amino Acids in the Pericarp of Z. planispinum
4.2. Relationship between Soil Nutrients and Pericarp Quality
5. Conclusions
- (1)
- Planting with S. tonkinensis or L. japonica can significantly increase the concentration of FAAs in the pericarp of Z. planispinum, which is conducive to the formation of pericarp quality. Based on the results of soil quality analysis in the early research and the amino acids in the pericarp, the optimal planting combination was found to be Z. planispinum + L. japonica;
- (2)
- As a characteristic element of karst, available Ca had a high contribution rate to the accumulation of FAAs in the pericarp, which had a positive impact;
- (3)
- The effect of available nutrients on FAAs in the pericarp was greater than that of total nutrients. Soil management in plantations should involve paying more attention to the concentration and proportion of available nutrients.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin. Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Y. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Mandrioli, R.; Mercolini, L.; Raggi, M.A. Recent trends in the analysis of amino acids in fruits and derived foodstuffs. Anal. Bioanal. Chem. 2013, 405, 7941–7956. [Google Scholar] [CrossRef] [PubMed]
- Ardo, Y. Flavour formation by amino acid catabolism. Biotechnol. Adv. 2006, 24, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Hong, T.N.; Shen, G.H.; Gu, Y.T.; Guo, Y.Z.; Han, J. Amino Acid Profiles and Nutritional Evaluation of Fresh Sweet–Waxy Corn from Three Different Regions of China. Nutrients 2022, 14, 3887. [Google Scholar] [CrossRef] [PubMed]
- Jafarikouhini, N.; Kazemeini, S.A.; Sinclair, T.R. Sweet corn nitrogen accumulation, leaf photosynthesis rate, and radiation use efficiency under variable nitrogen fertility and irrigation. Field Crops Res. 2020, 257, 107913. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood. Glob. Ecol. Biogeogr. 2013, 22, 494–507. [Google Scholar] [CrossRef]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting Nitrogen Metabolism and Transport Processes to Improve Plant Nitrogen Use Efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef]
- Aires, A.; Rosa, E.; Carvalho, R. Effect of nitrogen and sulfur fertilization on glucosinolates in the leaves and roots of broccoli sprouts (Brassica oleracea var. italica). J. Sci. Food Agric. 2006, 86, 1512–1516. [Google Scholar] [CrossRef]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Becquer, A.; Trap, J.; Irshad, U.; Ali, M.A.; Claude, P. From soil to plant, the journey of P through trophic relationships and ectomycorrhizal association. Front. Plant Sci 2014, 5, 548. [Google Scholar] [CrossRef] [PubMed]
- Thor, K. Calcium-Nutrient and Messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Ciesla, A.; Mitula, F.; Misztal, L.; Fedorowicz-Stronska, O.; Janicka, S.; Tajdel-Zielinska, M.; Marczak, M.; Janicki, M.; Ludwikow, A.; Sadowski, J. A Role for Barley Calcium-Dependent Protein Kinase CPK2a in the Response to Drought. Front. Plant Sci. 2016, 7, 1550. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Xu, X.L. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.R.; Boersma, M.; Raubenheimer, D. Stoichiometry: Linking elements to biochemicals. Ecology 2004, 85, 1193–1202. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.; Estiarte, M.; Penuelas, J. Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef]
- Tu, Y.L. The study of nutritive components and trace elements of flash of Zhnthoxylum planispinum var. dingtanensis. J. Guizhou Norm. Univ. (Nat. Sci.) 2000, 18, 31–36. (In Chinese) [Google Scholar]
- Duchene, O.; Vian, J.F.; Celette, F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Dong, Y.; Tang, L.; Zheng, Y.; Wei, L.F. Effects of N application on rhizosphere microflora and fusarium wilt occurrence of intercropped faba bean. Acta Ecol. Sin. 2010, 30, 1797–1805. (In Chinese) [Google Scholar]
- Yang, Z.X.; Tang, L.; Zheng, Y.; Dong, K.; Dong, Y. Effects of different wheat cultivars intercropped with faba bean on faba bean Fusarium wilt, root exudates and rhizosphere microbial community functional diversity. J. Plant Nutr. Fertil. 2014, 20, 570–579. (In Chinese) [Google Scholar] [CrossRef]
- Li, Y.T.; Yu, Y.H.; Song, Y.P. Stoichiometry of Soil, Microorganisms, and Extracellular Enzymes of Zanthoxylum planispinum var. dintanensis Plantations for Different Allocations. Agronomy 2022, 12, 1709. [Google Scholar] [CrossRef]
- Li, Y.T.; Yu, Y.H.; Song, Y.P. Soil properties of different planting combinations of Zanthoxylum planispinum var. dintanensis plantations and their driving force on stoichiometry. Agronomy 2022, 12, 2562. [Google Scholar] [CrossRef]
- do Nascimento, T.M.T.; Mansano, C.F.M.; Peres, H.; Rodrigues, F.H.F.; Khan, K.U.; Romaneli, R.S.; Sakomura, N.K.; Fernandes, J.B.K. Determination of the optimum dietary essential amino acid profile for growing phase of Nile tilapia by deletion method. Aquaculture 2020, 523, 735204. [Google Scholar] [CrossRef]
- Lu, P.Y.; Wang, J.; Wu, S.G.; Gao, J.; Dong, Y.; Zhang, H.J.; Qi, G.H. Standardized ileal digestible amino acid and metabolizable energy content of wheat from different origins and the effect of exogenous xylanase on their determination in broilers. Poult. Sci. 2020, 99, 992–1000. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, M. Non-volatile taste active compounds in the meat of Chinese mitten crab (Eriocheir sinensis). Food Chem. 2007, 104, 1200–1205. [Google Scholar] [CrossRef]
- Liu, Y.; Qiu, C.P. Calculated Taste Activity Values and Umami Equivalences Explain Why Dried Sha-chong (Sipunculus nudus) Is a Valuable Condiment. J. Aquat. Food Prod. Technol. 2016, 25, 177–184. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P. Carotenoids, chlorophylls, vitamin E and amino acid profile in fruits of nineteen Chaenomeles cultivars. J. Food Compos. Anal. 2020, 93, 103608. [Google Scholar] [CrossRef]
- Guevara-Terán, M.; Padilla-Arias, K.; Beltrán-Novoa, A.; Gonzalez-Paramas, A.M.; Giampieri, F.; Battino, M.; Vasquez-Castillo, W.; Fernandez-Soto, P.; Tejera, E.; Alvarez-Suarez, J.M. Influence of Altitudes and Development Stages on the Chemical Composition, Antioxidant, and Antimicrobial Capacity of the Wild Andean Blueberry (Vaccinium floribundum Kunth). Molecules 2022, 27, 7525. [Google Scholar] [CrossRef]
- Ozturk, M.; Unal, B.T.; Garcia-Caparros, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Scagel, C.F.; Lee, J. N, P, and K supply to Pinot noir grapevines: Impact on berry phenolics and free amino acids. Am. J. Enol. Vitic. 2014, 65, 43–49. [Google Scholar] [CrossRef]
- Wang, D.C.; Fu, Y.R.; Yu, Y.H.; Chen, M.; Wei, C.S. Amino acid accumulation characteristics in pericarp of germinating branches of Zanthoxylum planispinum var. dintanensis in different seasons. J. South. Agric. 2022, 53, 1963–1972. (In Chinese) [Google Scholar] [CrossRef]
- Gargallo–Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Penuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhu, S.S.; Wang, Z.H.; Chen, D.M.; Dai, G.H.; Feng, B.W.; Su, X.Y.; Hu, H.F.; Li, K.H.; Han, W.X.; et al. Divergent accumulation of microbial necromass and plant lignin components in grassland soils. Nat. Commun. 2018, 9, 3480. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Chen, P.; Liu, S.S.; Luo, K.; Zheng, B.C.; Yang, H.; He, S.; Yang, W.Y.; Yong, T.W. Effect of field microclimate on the difference of soybean flower morphology under maize-soybean relay strip intercropping system. Sci. Agric. Sin. 2021, 54, 2746–2758. (In Chinese) [Google Scholar] [CrossRef]
- Greger, H. Alkamides: A critical reconsideration of a multifunctional class of unsaturated fatty acid amides. Phytochem. Rev. 2016, 15, 729–770. [Google Scholar] [CrossRef]
- Lin, Z.N.; Lu, Z.; Lin, Y.; Li, Y.C.; Liu, M.X. Growth and Quality of Ganoderma lucidum at Tea Plantations under Relay Cropping. Fujian J. Agric. Sci. 2020, 35, 532–537. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, D.Q.; Wang, H.Y.; Tang, C.Z.; Jiang, J.Y.; Du, Y.X.; Zhang, Y.; Wang, S.; Guo, L.P. Influence and mechanism of stress combination on medicinal plants secondary metabolism. China J. Chin. Mater. Med. 2020, 45, 2009–2016. (In Chinese) [Google Scholar]
- Reggiani, R.; Nebuloni, M.; Mattana, M.; Brambilla, I. Anaerobic accumulation of amino acids in rice roots: Role of the glutamine synthetase/glutamate synthase cycle. Amino Acids 2000, 18, 207–217. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savoure, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef]
- Zhong, C.; Jian, S.F.; Huang, J.; Jin, Q.Y.; Cao, X.C. Trade-off of within-leaf nitrogen allocation between photosynthetic nitrogen-use efficiency and water deficit stress acclimation in rice (Oryza sativa L.). Plant Physiol. Biochem. 2019, 135, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.K.; Tang, S.; Zhou, J.J.; Liu, M.J.; Xu, M.; Kuzyakov, Y.; Ma, Q.X.; Wu, L.H. Plant-microbial competition for amino acids depends on soil acidity and the microbial community. Plant Soil 2022, 475, 457–471. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Nam, T.G.; Kim, D.O.; Eom, S.H. Effects of light sources on major flavonoids and antioxidant activity in common buckwheat sprouts. Food Sci. Biotechnol. 2018, 27, 169–176. [Google Scholar] [CrossRef]
- Wang, Z.B.; Zhang, Y.Z.; Bo, G.D.; Zhang, Y.P.; Chen, Y.; Shen, M.C.; Zhang, P.; Li, G.T.; Zhou, J.; Li, Z.F.; et al. Ralstonia solanacearum infection disturbed the microbiome structure throughout the whole Tobacco Crop Niche as well as the nitrogen metabolism in soil. Front. Bioeng. Biotechnol. 2022, 10, 903555. [Google Scholar] [CrossRef]
- Hu, W.; Lv, X.B.; Yang, J.S.; Chen, B.L.; Zhao, W.Q.; Meng, Y.L.; Wang, Y.H.; Zhou, Z.G.; Oosterhuis, D.M. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crops Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Raddatz, N.; de los Ríos, L.M.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef]
- Li, P.F.; Liu, M.; Li, G.L.; Liu, K.; Liu, T.S.; Wu, M.; Saleem, M.; Li, Z.P. Phosphorus availability increases pathobiome abundance and invasion of rhizosphere microbial networks by Ralstonia. Environ. Microbiol. 2021, 23, 5992–6003. [Google Scholar] [CrossRef]
- Song, X.H.; Xie, K.; Zhao, H.B.; Li, Y.L.; Dong, C.X.; Xu, Y.C.; Shen, Q.R. Effects of different organic fertilizers on tree growth, yield, fruit quality, and soil microorganisms in a pear orchard. Eur. J. Hortic. Sci. 2012, 77, 204–210. [Google Scholar]
- Li, H.; Liu, S.Q.; Chen, X.; Wang, Y.; Feng, L.; Liu, J.K. Effects of calcium on growth, photosynthetic characteristics and quality of aquicultural garlic seedlings. J. Plant Nutr. Fertil. 2013, 19, 1118–1128. (In Chinese). Available online: https://x.cnki.net/kcms/detail/detail.aspx?dbcode=CJFD&dbname=CJFD2013&filename=ZWYF201305012&v=x0QVnnll88REIYoqsflQFp%mmd2Fokke2LEPXJdqx1nK4ZPctjFPpE3eziAAq5Di%mmd2F7FXw (accessed on 23 February 2023).
- Li, H.; Liu, S.Q.; Liu, Z.L.; Feng, L.; Liu, J.K.; Chen, X.W.; Wang, Y. Effects of Calcium on Physiological Characteristics and Main Mineral Elements Absorption of Garlic. Sci. Agric. Sin. 2013, 46, 3626–3634. (In Chinese) [Google Scholar] [CrossRef]
- Poovaiah, H.W.; Redd, A.S.N. Calcium and signal transduction in plants. Crit. Rev. Plant Sci. 1993, 12, 185–211. [Google Scholar] [CrossRef] [PubMed]
- Hang, Y.; Wen, L.; Pang, Y.Z.; Hang, B.Y.; Wang, J.; Lü, X.L. Effect of spraying calcium on sugar and acid accumulation in ‘Summer Black’ grape. Soil Fertil. Sci. China 2020, 2, 166–172. (In Chinese) [Google Scholar] [CrossRef]
- Yue, Y.K.; Jin, Z.Y.; Zhang, M.; Li, Z.Y. Effects of different nitrogen and calcium levels on fruit quality of protected peach. China Fruits 2021, 4, 55–58. (In Chinese) [Google Scholar] [CrossRef]
- Sneha, G.R.; Swarnalakshmi, K.; Sharma, M.; Reddy, K.; Bhoumik, A.; Suman, A.; Kannepalli, A. Soil Type Influence Nutrient Availability, Microbial Metabolic Diversity, Eubacterial and Diazotroph Abundance in Chickpea Rhizosphere. World J. Microbiol. Biotechnol. 2021, 37, 167. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plantation Types | Species Combinations | Longitude | Latitude | Growing Area (ha) | Altitude (m asl) | Density (m) | Height (m) | Crown Width (m) | Coverage (%) |
|---|---|---|---|---|---|---|---|---|---|
| Trt 1 | Z. planispinum + P. salicina | 105°40′28.33″ E | 25°37′57.41″ N | 1.34 | 764 | 3 × 3 | 3.5 | 2 × 2.3 | 70 |
| Trt 2 | Z. planispinum + S. tonkinensis | 105°40′19.79″ E | 25°39′25.75″ N | 0.67 | 728 | 2 × 2 | 2.0 | 1.2 × 1.8 | 60 |
| Trt 3 | Z. planispinum + A. hypogaea | 105°38′36.32″ E | 25°39′23.64″ N | 0.67 | 791 | 2 × 2 | 2.5 | 2.5 × 2.8 | 85 |
| Trt 4 | Z. planispinum + L. japonica | 105°38′36.35″ E | 25°39′22.29″ N | 6.67 | 814 | 3.5 × 3 | 2.5 | 1.5 × 2.5 | 70 |
| Trt 5 | Z. planispinum | 105°38′35.64″ E | 25°39′23.35″ N | 33.35 | 788 | 3 × 4 | 2.2 | 2.5 × 2.3 | 65 |
| Soil Parameters | Trt 1 | Trt 2 | Trt 3 | Trt 4 | Trt 5 |
|---|---|---|---|---|---|
| Soil water content | 31.60 ± 6.29 ab | 36.73 ± 2.65 a | 25.05 ± 1.38 bc | 28.69 ± 0.30 bc | 21.15 ± 0.14 c |
| Soil organic carbon concentration | 37.73 ± 7.32 ab | 29.40 ± 0.57 ab | 28.68 ± 12.62 ab | 50.83 ± 13.33 a | 26.50 ± 2.19 b |
| Total nitrogen concentration | 3.53 ± 0.46 ab | 2.64 ± 0.07 b | 2.78 ± 0.74 b | 4.60 ± 0.44 a | 2.76 ± 0.23 b |
| Total phosphorus concentration | 1.37 ± 0.02 a | 0.82 ± 0.03 b | 1.10 ± 0.43 ab | 1.52 ± 0.17 a | 1.26 ± 0.04 ab |
| Total potassium concentration | 6.95 ± 0.34 b | 6.11 ± 1.51 b | 12.33 ± 0.25 a | 11.88 ± 0.53 a | 10.88 ± 0.03 a |
| Total calcium concentration | 0.95 ± 0.28 b | 1.48 ± 0.39 b | 1.85 ± 0.71 b | 1.88 ± 0.18 b | 6.05 ± 0.21 a |
| Available nitrogen concentration | 275.00 ± 74.25 ab | 160.00 ± 5.66 b | 161.75 ± 61.87 b | 350.00 ± 55.15 a | 153.75 ± 15.91 b |
| Available phosphorus concentration | 45.80 ± 13.29 a | 23.38 ± 11.63 a | 26.55 ± 10.54 a | 36.68 ± 10.01 a | 20.08 ± 2.44 a |
| Available potassium concentration | 393.00 ± 107.48 a | 195.85 ± 32.03 b | 172.75 ± 57.63 b | 223.75 ± 98.64 ab | 141.25 ± 2.47 b |
| Available calcium concentration | 317.50 ± 14.85 b | 334.75 ± 0.35 b | 347.75 ± 24.40 ab | 371.00 ± 8.49 a | 350.50 ± 7.07 ab |
| Soil C:N ratio | 10.65 ± 0.70 a | 11.13 ± 0.53 a | 10.07 ± 1.85 a | 10.97 ± 1.85 a | 9.59 ± 0.00 a |
| Soil C:P ratio | 27.68 ± 5.79 abc | 35.83 ± 0.79 a | 25.82 ± 1.33 bc | 33.10 ± 4.99 ab | 21.03 ± 2.38 c |
| Soil N:P ratio | 2.59 ± 0.37 ab | 3.22 ± 0.22 a | 2.60 ± 0.34 ab | 3.02 ± 0.05 a | 2.19 ± 0.25 b |
| Microbial biomass carbon | 243.00 ± 4.95 a | 254.75 ± 2.47 a | 252.00 ± 2.83 a | 262.75 ± 21.57 a | 262.25 ± 26.52 a |
| Microbial biomass nitrogen | 12.40 ± 1.70 a | 13.58 ± 1.31 a | 14.38 ± 0.60 a | 13.90 ± 1.06 a | 14.08 ± 0.18 a |
| Microbial biomass phosphorus | 128.00 ± 23.33 a | 144.50 ± 4.95 a | 148.00 ± 8.49 a | 154.50 ± 13.44 a | 139.00 ± 3.54 a |
| Amino Acid | Trt 1 | Trt 2 | Trt 3 | Trt 4 | Trt 5 | |
|---|---|---|---|---|---|---|
| Essential amino acids (mg/kg) | Valine | 376.51 ± 10.86 b | 500.15 ± 87.33 a | 420.97 ± 10.85 ab | 438.20 ± 8.85 ab | 409.29 ± 22.74 ab |
| Threonine | 282.72 ± 7.50 b | 368.39 ± 58.89 a | 314.29 ± 6.22 ab | 337.44 ± 16.76 ab | 304.01 ± 22.22 ab | |
| Phenylalanine | 8.46 ± 1.40 c | 33.19 ± 2.62 b | 23.96 ± 8.93 bc | 20.92 ± 8.22 bc | 55.35 ± 11.82 a | |
| Methionine | 33.71 ± 0.04 b | 46.81 ± 8.85 a | 41.84 ± 3.45 ab | 34.09 ± 4.92 ab | 41.17 ± 3.22 ab | |
| Leucine | 471.19 ± 10.18 b | 608.99 ± 98.29 a | 514.84 ± 1.06 ab | 555.49 ± 24.27 ab | 507.19 ± 25.65 ab | |
| Lysine | 347.38 ± 0.42 b | 470.78 ± 90.79 a | 361.81 ± 47.02 ab | 384.33 ± 16.50 ab | 386.29 ± 2.37 ab | |
| Isoleucine | 43.18 ± 6.89 b | 458.46 ± 90.71 a | 375.71 ± 12.23 a | 370.20 ± 25.94 a | 380.95 ± 12.11 a | |
| Total | 1563.14 ± 36.47 b | 2486.75 ± 437.47 a | 2053.41 ± 68.29 ab | 2140.67 ± 27.30 a | 2084.22 ± 70.05 a | |
| Nonessential amino acids (mg/kg) | Histidine | 175.01 ± 1.83 b | 224.90 ± 36.95 a | 189.15 ± 11.31 ab | 205.69 ± 7.85 ab | 187.80 ± 10.38 ab |
| Arginine | 367.86 ± 21.83 a | 496.41 ± 125.63 a | 439.46 ± 43.59 a | 420.37 ± 6.47 a | 396.40 ± 12.56 a | |
| Serine | 325.79 ± 12.94 a | 447.59 ± 98.81 a | 380.91 ± 22.85 a | 406.63 ± 16.93 a | 366.69 ± 25.24 a | |
| Proline | 427.79 ± 36.36 a | 608.59 ± 128.90 a | 458.82 ± 88.71 a | 601.17 ± 21.00 a | 533.05 ± 87.49 a | |
| Glycine | 381.66 ± 17.76 b | 491.17 ± 83.21 a | 422.02 ± 14.86 ab | 437.38 ± 9.76 ab | 416.49 ± 0.92 ab | |
| Glutamate | 738.39 ± 30.78 b | 1012.02 ± 203.02 a | 863.35 ± 9.32 ab | 899.38 ± 18.59 ab | 846.80 ± 22.80 ab | |
| Aspartate | 477.48 ± 48.72 b | 744.83 ± 181.96 a | 646.66 ± 126.96 ab | 597.16 ± 8.89 ab | 578.21 ± 1.11 ab | |
| Alanine | 363.17 ± 9.57 a | 454.11 ± 69.92 a | 386.75 ± 0.01 a | 406.52 ± 20.65 a | 377.00 ± 29.29 a | |
| Tyrosine | 232.20 ± 8.64 a | 270.88 ± 19.56 a | 245.27 ± 11.75 a | 273.96 ± 10.92 a | 221.14 ± 37.96 a | |
| Cystine | 52.02 ± 4.85 a | 48.37 ± 2.22 a | 39.48 ± 8.34 a | 54.93 ± 12.49 a | 37.41 ± 4.61 a | |
| Total | 2998.50 ± 87.20 b | 4077.54 ± 783.16 a | 3443.26 ± 236.39 ab | 3677.10 ± 59.48 ab | 3376.77 ± 209.43 ab | |
| Total free amino acids (mg/kg) | 5104.49 ± 147.33 b | 7285.59 ± 1383.21 a | 6125.28 ± 359.58 ab | 6443.82 ± 88.16 ab | 6045.19 ± 302.42 ab | |
| EAAs/TFAAs (%) | 30.62 | 34.13 | 33.52 | 33.22 | 34.48 | |
| EAAs/NEAAs (%) | 44.14 | 51.82 | 50.43 | 49.75 | 52.62 | |
| Amino Acid | Trt 1 | Trt 2 | Trt 3 | Trt 4 | Trt 5 |
|---|---|---|---|---|---|
| Bitter amino acid (mg/kg) | 1292.44 ± 49.82 b | 2110.81 ± 410.80 a | 1792.82 ± 62.15 a | 1818.35 ± 4.21 a | 1734.99 ± 69.83 ab |
| Sweet amino acid (mg/kg) | 1956.13 ± 13.25 b | 2594.73 ± 476.69 a | 2151.94 ± 101.79 ab | 2394.82 ± 50.96 ab | 2185.03 ± 175.55 ab |
| Delicious amino acid (mg/kg) | 1563.25 ± 79.08 b | 2227.62 ± 475.77 a | 1871.82 ± 183.31 ab | 1880.86 ± 26.21 ab | 1811.29 ± 26.28 ab |
| Aromatic amino acid (mg/kg) | 292.68 ± 5.19 b | 352.43 ± 19.97 a | 308.71 ± 12.34 ab | 349.79 ± 15.20 a | 313.89 ± 30.76 ab |
| Amino Acid | Taste Threshold [27] (mg/g) | Taste Activity Value | |||||
|---|---|---|---|---|---|---|---|
| Trt 1 | Trt 2 | Trt 3 | Trt 4 | Trt 5 | |||
| Bitter amino acid | Valine | 1.50 | 0.25 | 0.33 | 0.28 | 0.29 | 0.27 |
| Leucine | 3.80 | 0.12 | 0.16 | 0.14 | 0.15 | 0.13 | |
| Isoleucine | 0.90 | 0.05 | 0.51 | 0.42 | 0.41 | 0.42 | |
| Methionine | 0.30 | 0.11 | 0.16 | 0.14 | 0.11 | 0.14 | |
| Arginine | 0.10 | 3.68 | 4.96 | 4.39 | 4.20 | 3.96 | |
| Sweet amino acid | Glycine | 1.10 | 0.35 | 0.45 | 0.38 | 0.40 | 0.38 |
| Alanine | 0.60 | 0.61 | 0.76 | 0.64 | 0.68 | 0.63 | |
| Serine | 1.50 | 0.22 | 0.30 | 0.25 | 0.27 | 0.24 | |
| Threonine | 2.60 | 0.11 | 0.14 | 0.12 | 0.13 | 0.12 | |
| Proline | 3.00 | 0.14 | 0.20 | 0.15 | 0.20 | 0.18 | |
| Histidine | 0.20 | 0.88 | 1.12 | 0.95 | 1.03 | 0.94 | |
| Delicate amino acid | Lysine | 0.50 | 0.69 | 0.94 | 0.72 | 0.77 | 0.77 |
| Glutamate | 0.05 | 14.77 | 20.24 | 17.27 | 17.99 | 16.94 | |
| Aspartate | 0.03 | 15.92 | 24.83 | 21.56 | 19.91 | 19.27 | |
| Aromatic amino acid | Phenylalanine | 1.50 | 0.01 | 0.02 | 0.02 | 0.01 | 0.04 |
| Tyrosine | 2.60 | 0.09 | 0.10 | 0.09 | 0.11 | 0.09 | |
| Cystine | 0.02 | 2.60 | 2.42 | 1.97 | 2.75 | 1.87 | |
| Factors | Principal Component Load Matrix | |
|---|---|---|
| PC1 | PC2 | |
| Aspartate | 0.866 | 0.378 |
| Glutamate | 0.971 | 0.212 |
| Serine | 0.988 | 0.102 |
| Histidine | 0.993 | 0.031 |
| Glycine | 0.951 | 0.174 |
| Threonine | 0.989 | −0.021 |
| Arginine | 0.916 | 0.249 |
| Alanine | 0.986 | −0.034 |
| Tyrosine | 0.811 | −0.458 |
| Cystine | 0.157 | −0.915 |
| Valine | 0.989 | 0.123 |
| Methionine | 0.563 | 0.673 |
| Phenylalanine | 0.111 | 0.881 |
| Isoleucine | 0.702 | 0.513 |
| Leucine | 0.993 | 0.019 |
| Lysine | 0.901 | 0.231 |
| Proline | 0.838 | 0.055 |
| Eigenvalue | 12.639 | 2.599 |
| Variance contribution rate/% | 72.557 | 17.076 |
| Cumulative variance contribution rate/% | 72.557 | 89.633 |
| Plantation Types | Factor Score | Comprehensive Score | Ranking | |
|---|---|---|---|---|
| PC1 | PC2 | |||
| Trt 1 | −3.78 | −2.71 | −3.20 | 5 |
| Trt 2 | 4.46 | 1.69 | 3.52 | 1 |
| Trt 3 | −0.52 | 0.56 | −0.28 | 3 |
| Trt 4 | 0.94 | −1.23 | 0.47 | 2 |
| Trt 5 | −1.10 | 1.70 | −0.51 | 4 |
| Soil Factors | Contribution/% | Pseudo-F | p |
|---|---|---|---|
| Available potassium | 0.646 | 23.763 | 0.002 |
| Available phosphorus | 0.472 | 11.626 | 0.002 |
| Microbial biomass nitrogen | 0.365 | 7.458 | 0.012 |
| Available calcium | 0.286 | 5.217 | 0.018 |
| Microbial biomass phosphorus | 0.235 | 3.989 | 0.038 |
| Available nitrogen | 0.218 | 3.633 | 0.052 |
| Microbial biomass carbon | 0.191 | 3.062 | 0.076 |
| Total calcium | 0.171 | 2.691 | 0.066 |
| Total phosphorus | 0.156 | 2.402 | 0.15 |
| Total potassium | 0.134 | 2.02 | 0.174 |
| Soil organic carbon | 0.131 | 1.955 | 0.146 |
| Total nitrogen | 0.125 | 1.854 | 0.15 |
| Soil N:P ratio | 0.087 | 1.233 | 0.276 |
| Soil C:N ratio | 0.078 | 1.106 | 0.354 |
| Soil water content | 0.072 | 1.012 | 0.354 |
| Soil C:P ratio | 0.071 | 0.995 | 0.364 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yu, Y.; Song, Y.; Wei, C. Influence of Different Planting Combinations on the Amino Acid Concentration in Pericarp of Zanthoxylum planispinum ‘Dintanensis’ and Soil. Forests 2023, 14, 843. https://doi.org/10.3390/f14040843
Li Y, Yu Y, Song Y, Wei C. Influence of Different Planting Combinations on the Amino Acid Concentration in Pericarp of Zanthoxylum planispinum ‘Dintanensis’ and Soil. Forests. 2023; 14(4):843. https://doi.org/10.3390/f14040843
Chicago/Turabian StyleLi, Yitong, Yanghua Yu, Yanping Song, and Changsheng Wei. 2023. "Influence of Different Planting Combinations on the Amino Acid Concentration in Pericarp of Zanthoxylum planispinum ‘Dintanensis’ and Soil" Forests 14, no. 4: 843. https://doi.org/10.3390/f14040843