
Comparative RNA-Seq Analysis Reveals the Organ-Specific Transcriptomic Response to Zinc Stress in Mulberry


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determining the Contents of Physiological Indicators Related to Zinc Toxicity
2.3. RNA-Seq and Data Processing
2.4. Workflow for Comprehensive Transcriptome Analysis
2.5. RT-qPCR Analysis of Key Genes Involved in the Response to Zinc Toxicity
3. Results
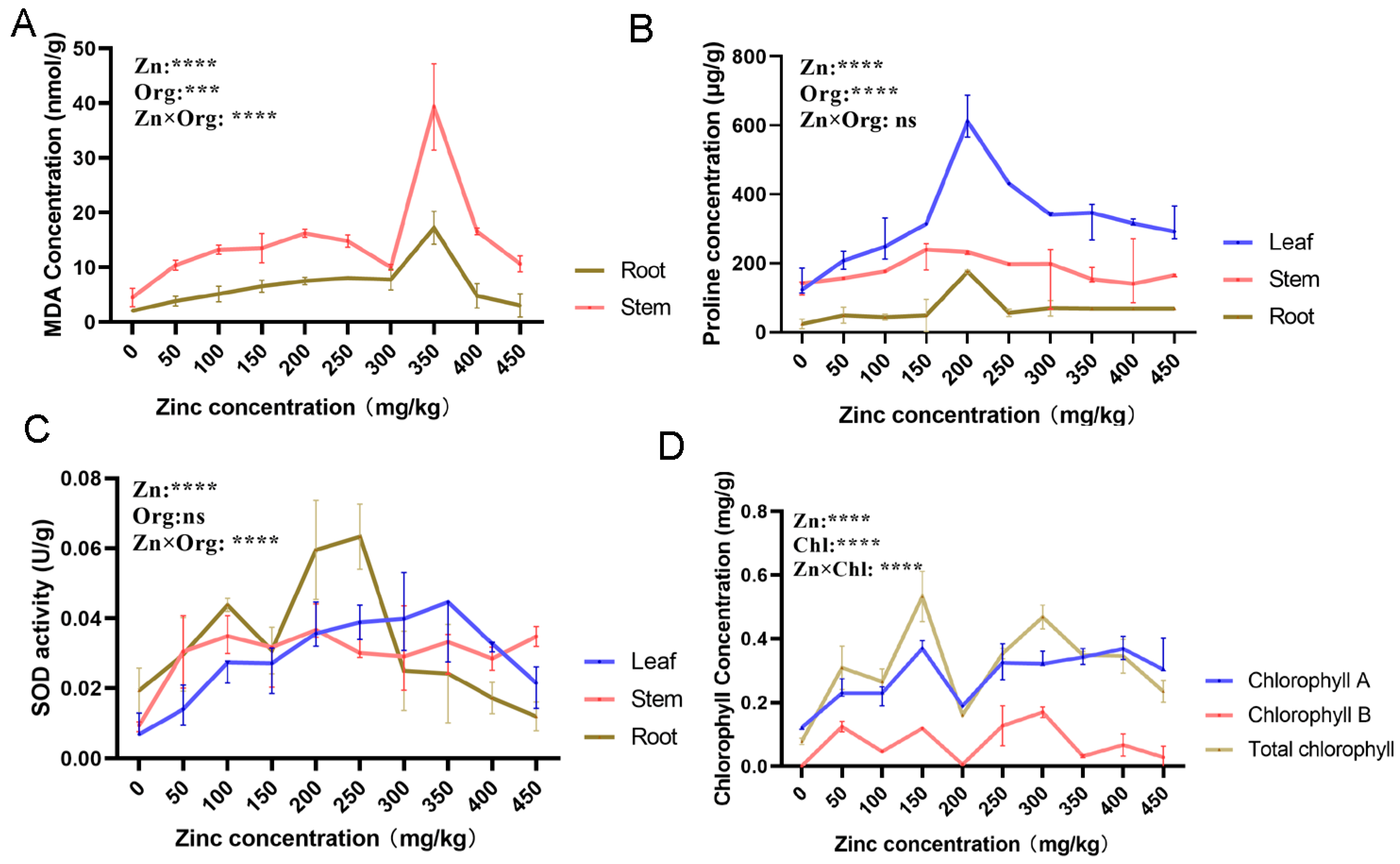
3.1. Mulberry Organ-Specific Physiological Responses to Zinc Treatment
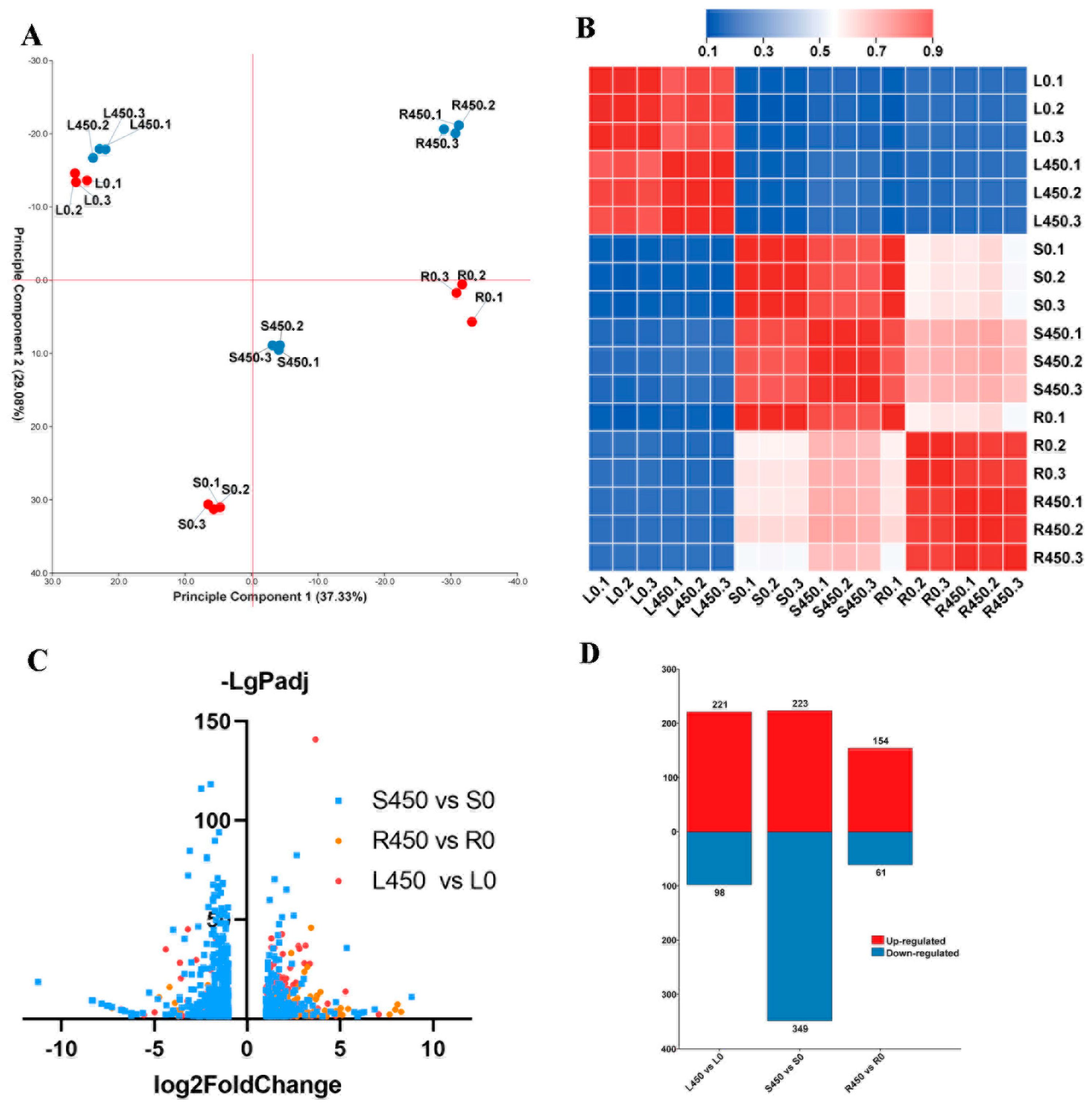
3.2. Transcriptomic Analysis Showed Organ-Specific Differences in Response to Excess Zinc Treatments
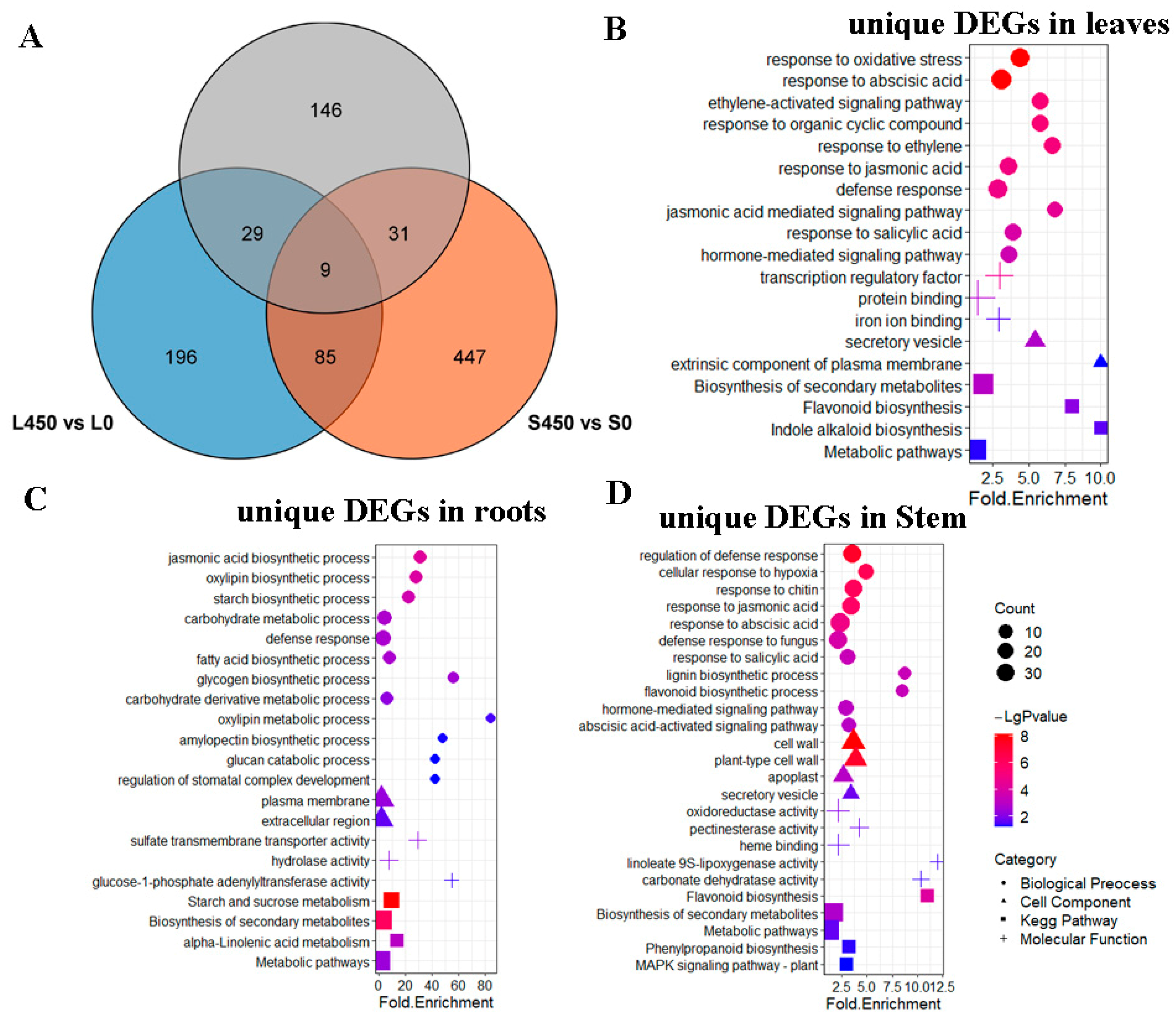
3.3. Organ-Specific DEGs in Mulberry under Zinc Stress
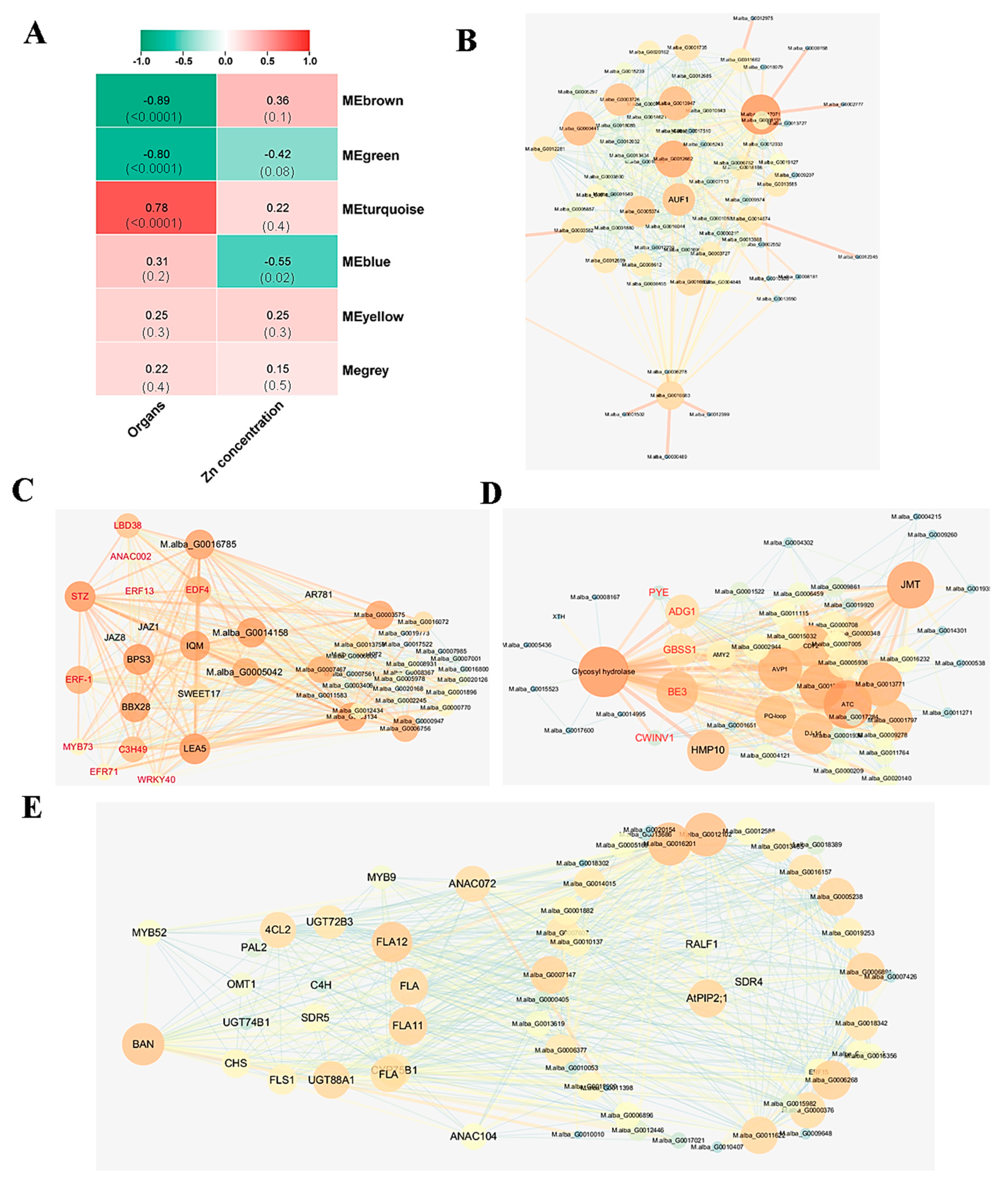
3.4. Network of Genes Involved in the Response to Zinc Toxicity in Different Organs
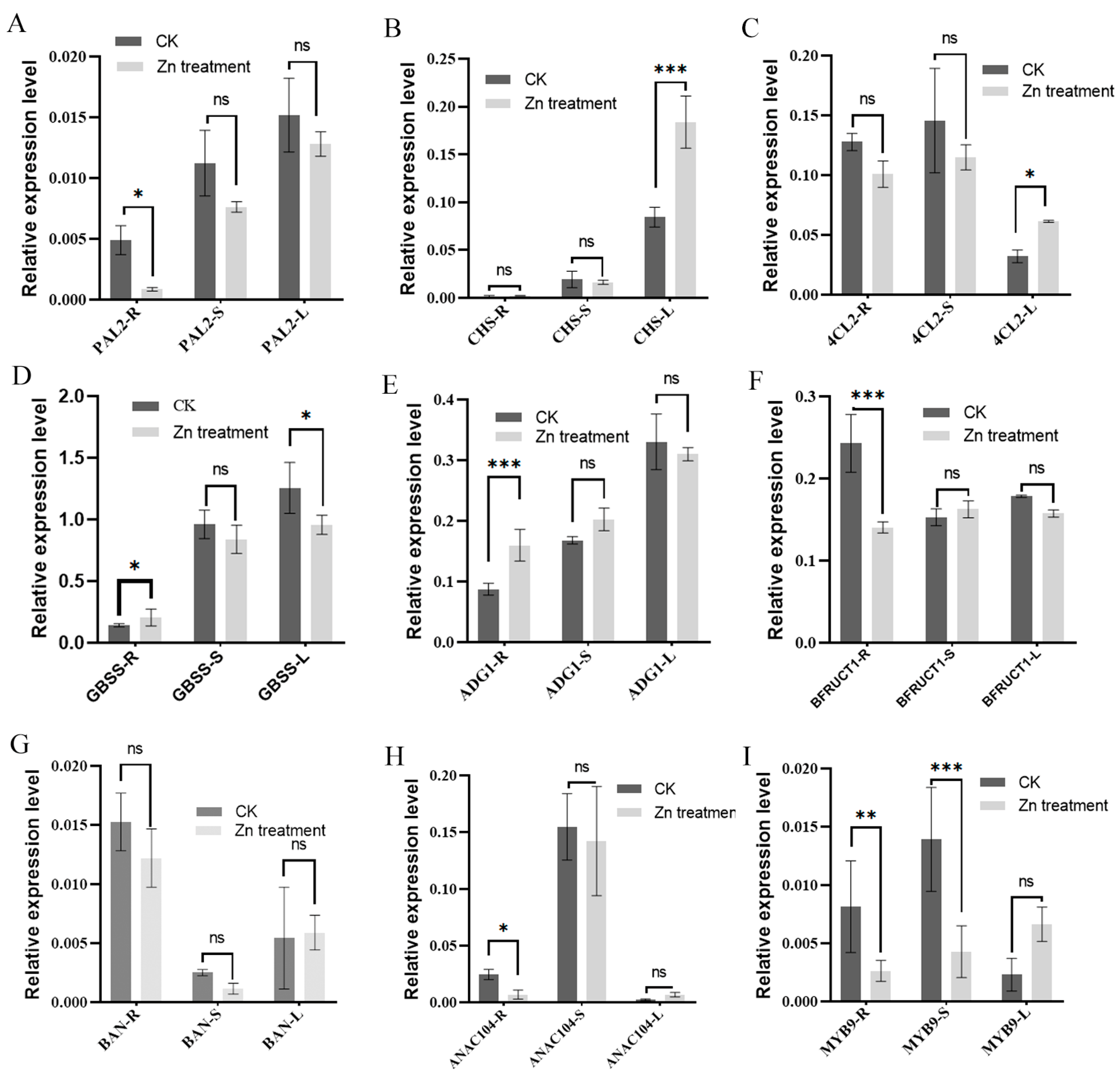
3.5. Validation of Transcription Levels of Key Genes Responsive to Zinc Toxicity
4. Discussion
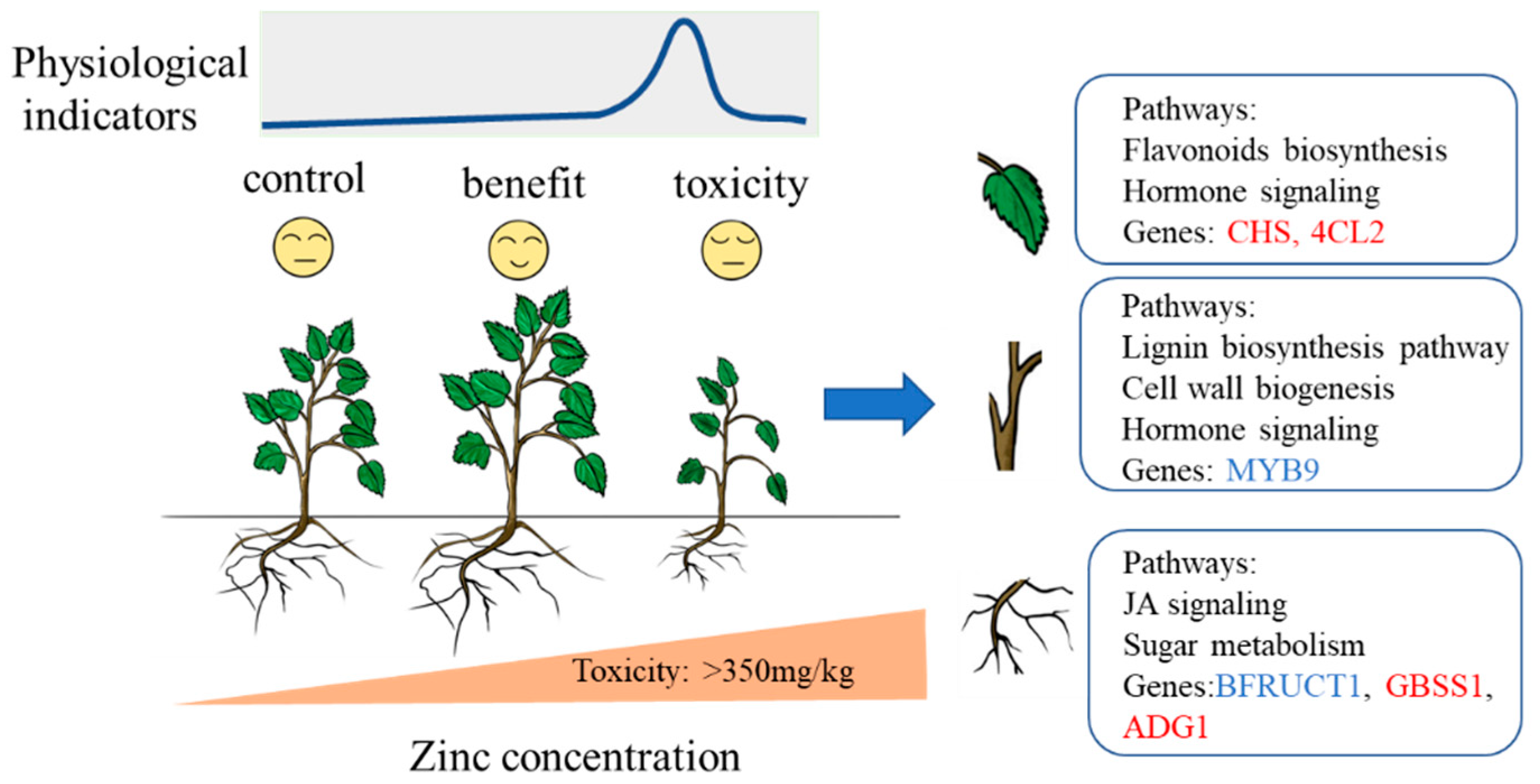
4.1. Limitation of Excess Zinc Supply in Soil for Mulberry
4.2. Organ-Specific Responses to Zinc Toxicity
4.3. Molecular Regulation Network of Genes in Response to Zinc Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hambidge, M. Human Zinc Deficiency. J. Nutr. 2000, 130 (Suppl. S5), 1344S. [Google Scholar] [CrossRef]
- Cherasse, Y.; Urade, Y. Dietary Zinc Acts as a Sleep Modulator. Int. J. Mol. Sci. 2017, 18, 2334. [Google Scholar] [CrossRef]
- Hotz, C.; Brown, K.H. Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr. Bull. 2004, 25, S130–S162. [Google Scholar]
- Gibson, R.S.; King, J.C.; Lowe, N. A Review of Dietary Zinc Recommendations. Food Nutr. Bull. 2016, 37, 443–460. [Google Scholar] [CrossRef]
- Alloway, B.J. Zinc in Soils and Crop Nutrition; International Zinc International Association: Brussels, Belgium; Fertilizer Industry Association: Paris, France, 2008. [Google Scholar]
- Tsonev, T.; Cebola Lidon, F.J. Zinc in plants-an overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Qin, F.; Liu, G.; Huang, G.; Dong, T.; Liao, Y.; Xu, X. Zinc application alleviates the adverse effects of lead stress more in female Morus alba than in males. Environ. Exp. Bot. 2018, 146, 68–76. [Google Scholar] [CrossRef]
- Zlobin, I.E. Current understanding of plant zinc homeostasis regulation mechanisms. Plant Physiol. Biochem. 2021, 162, 327–335. [Google Scholar] [CrossRef]
- Fuentes, D.; Disante, K.B.; Valdecantos, A.; Cortina, J.; Vallejo, V.R. Sensitivity of Mediterranean woody seedlings to copper, nickel and zinc. Chemosphere 2007, 66, 412–420. [Google Scholar] [CrossRef]
- Bose, S.; Bhattacharyya, A.K. Heavy metal accumulation in wheat plant grown in soil amended with industrial sludge. Chemosphere 2008, 70, 1264–1272. [Google Scholar] [CrossRef]
- McGrath, S.; Chang, A.; Page, A.; Witter, E. Land application of sewage sludge: Scientific perspectives of heavy metal loading limits in Europe and the United States. Environ. Rev. 1994, 2, 108–118. [Google Scholar] [CrossRef]
- Disante, K.B.; Fuentes, D.; Cortina, J. Response to drought of Zinc-stressed Quercus suber L. seedlings. Environ. Exp. Bot. 2011, 70, 96–103. [Google Scholar] [CrossRef]
- Pandey, N.; Pathak, G.C.; Singh, A.K.; Sharma, C.P. Enzymic changes in response to zinc nutrition. J. Plant Physiol. 2002, 159, 1151–1153. [Google Scholar] [CrossRef]
- Chakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Choudhary, M.; Bailey, L.; Grant, C.; Leisle, D. Effect of Zinc on the concentration of Cd and Zinc in plant tissue of two durum wheat lines. Can. J. Plant Sci. 1995, 75, 445–448. [Google Scholar] [CrossRef]
- Lin, Y.F.; Aarts, M.G. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- He, P.P.; Lv, X.Z.; Wang, G.Y. Effects of Se and Zinc supplementation on the antagonism against Pb and Cd in vegetables. Environ. Int. 2004, 30, 167–172. [Google Scholar] [CrossRef]
- Li, C.; Xu, G.; Zang, R.; Korpelainen, H.; Berninger, F. Sex-related differences in leaf morphological and physiological responses in Hippophae rhamnoides along an altitudinal gradient. Tree Physiol. 2007, 27, 399–406. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Ding, Z.; Wang, H.; Song, L.; Jia, S.; Ma, D. Zinc stress affects ionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 111, 318–328. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, R.; Yan, X.; Jia, C.; Long, T. Mulberry for environmental protection. Pak. J. Bot. 2017, 49, 781–788. [Google Scholar]
- Srivastava, S.; Kapoor, R.; Thathola, A.; Srivastava, R.P. Nutritional quality of leaves of some genotypes of mulberry (Morus alba). Int. J. Food Sci. Nutr. 2006, 57, 305–313. [Google Scholar] [CrossRef]
- Cao, X.; Du, W.; Shang, C.; Shen, Q.; Liu, L.; Cheng, J. Comparative transcriptome reveals circadian and hormonal control of adventitious rooting in mulberry hardwood cuttings. Acta Physiol. Plant. 2018, 40, 197. [Google Scholar] [CrossRef]
- Shang, C.; Yang, H.; Ma, S.; Shen, Q.; Liu, L.; Hou, C.; Cao, X.; Cheng, J. Physiological and Transcriptomic Changes during the Early Phases of Adventitious Root Formation in Mulberry Stem Hardwood Cuttings. Int. J. Mol. Sci. 2019, 20, 3707. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.; Yu, T.; Hou, C.; Liu, L.; Zhang, L. Genome-wide analysis of the lignin toolbox for morus and the roles of lignin related genes in response to zinc stress. PeerJ 2021, 9, e11964. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.; Wang, R.F.; Hou, C.; Yu, T.; Miao, K.; Cao, F.Y.; Fang, R.J.; Liu, L. Functional characterization of two chalcone isomerase (CHI) revealing their responsibility for anthocyanins accumulation in mulberry. Plant Physiol. Biochem. 2021, 161, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Susheelamma, B.N.; Shekar, K.R.; Sarkar, A.; Rao, M.R.; Datta, R.K. Genotype and hormonal effects on callus formation and regeneration in mulberry. Euphytica 1996, 90, 25–29. [Google Scholar] [CrossRef]
- Subba, P.; Mukhopadhyay, M.; Mahato, S.K.; Bhutia, K.D.; Mondal, T.K.; Ghosh, S.K. Zinc stress induces physiological, ultra-structural and biochemical changes in mandarin orange (Citrus reticulata Blanco) seedlings. Physiol. Mol. Biol. Plants 2014, 20, 461–473. [Google Scholar] [CrossRef]
- Arora, A.; Sairam, R.; Srivastava, G. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Silva, P.; Matos, M. Assessment of the impact of Aluminum on germination, early growth and free proline content in Lactuca sativa L. Ecotoxicol. Environ. Saf. 2016, 131, 151–156. [Google Scholar] [CrossRef]
- Langdon, W.B. Performance of genetic programming optimised Bowtie2 on genome comparison and analytic testing (GCAT) benchmarks. BioData Min. 2015, 8, 1. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Nat. Preced. 2010, 2010, 5. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35 (Suppl. S2), W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Reddy, R.A.; Ponnuvel, K.M.; Rohela, G.K.; Shabnam, A.A.; Ghosh, M.K.; Mishra, R.K. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Mulberry (Morus alba L.) under different abiotic stresses. Mol. Biol. Rep. 2019, 46, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Kumar Tewari, R.; Kumar, P.; Nand Sharma, P. Morphology and physiology of zinc-stressed mulberry plants. J. Plant Nutr. Soil Sci. 2008, 171, 286–294. [Google Scholar] [CrossRef]
- Liu, L.; Chao, N.; Yidilisi, K.; Kang, X.; Cao, X. Comprehensive analysis of the MYB transcription factor gene family in Morus alba. BMC Plant Biol. 2022, 22, 281. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Monnet, F.; Vaillant, N.; Vernay, P.; Coudret, A.; Sallanon, H.; Hitmi, A. Relationship between PSII activity, CO2 fixation, and Zinc, Mn and Mg contents of Lolium perenne under zinc stress. J. Plant Physiol. 2001, 158, 1137–1144. [Google Scholar] [CrossRef]
- Roitto, M.; Rautio, P.; Julkunen-Tiitto, R.; Kukkola, E.; Huttunen, S. Changes in the concentrations of phenolics and photosynthates in Scots pine (Pinus sylvestris L.) seedlings exposed to nickel and copper. Environ. Pollut. 2005, 137, 603–609. [Google Scholar] [CrossRef]
- Burzyński, M.; Żurek, A. Effects of copper and cadmium on photosynthesis in cucumber cotyledons. Photosynthetica 2007, 45, 239–244. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Xin, H.; Xie, Y.; Guo, T. Physiological Responses and Yield of Wheat Plants in Zinc-Mediated Alleviation of Drought Stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Guo, Q.; Liu, C.; Liu, X.; Zhang, M.; Long, D.; Xiang, Z.; Zhao, A. Two mulberry phytochelatin synthase genes confer zinc/cadmium tolerance and accumulation in transgenic Arabidopsis and tobacco. Gene 2018, 645, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Shi, D.; Liu, X.; Qin, J.; Ge, Q.; Xu, L.; Pan, X.; Li, W.; Zhu, Y.; et al. Spatial-temporal analysis of zinc homeostasis reveals the response mechanisms to acute zinc deficiency in Sorghum bicolor. New Phytol. 2013, 200, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217 Pt 1, 67–75. [Google Scholar] [CrossRef]
- Azevedo, H.; Azinheiro, S.G.; Munoz-Merida, A.; Castro, P.H.; Huettel, B.; Aarts, M.G.; Assuncao, A.G. Transcriptomic profiling of Arabidopsis gene expression in response to varying micronutrient zinc supply. Genome Data 2016, 7, 256–258. [Google Scholar] [CrossRef]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of sugars under abiotic stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; Gonzalez, J.A.; Hilal, M.; Prado, F.E. Soluble sugars--metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Illgen, S.; Zintl, S.; Zuther, E.; Hincha, D.K.; Schmulling, T. Characterisation of the ERF102 to ERF105 genes of Arabidopsis thaliana and their role in the response to cold stress. Plant Mol. Biol. 2020, 103, 303–320. [Google Scholar] [CrossRef]
- Je, J.; Chen, H.; Song, C.; Lim, C.O. Arabidopsis DREB2C modulates ABA biosynthesis during germination. Biochem. Biophys. Res. Commun. 2014, 452, 91–98. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, S.J.; Kim, S.Y. AtERF15 is a positive regulator of ABA response. Plant Cell Rep. 2015, 34, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.R.; Xie, X.L.; Xia, X.J.; Yu, J.Q.; Ferguson, I.B.; Giovannoni, J.J.; Chen, K.S. Involvement of an ethylene response factor in chlorophyll degradation during citrus fruit degreening. Plant J. 2016, 86, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, L.; Dai, Y.; Liu, S.; Hong, Y.; Tian, L.; Huang, L.; Cao, Z.; Li, D.; Song, F. Arabidopsis AtERF15 positively regulates immunity against Pseudomonas syringae pv. tomato DC3000 and Botrytis cinerea. Front. Plant Sci. 2015, 6, 686. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Organ | Ortholog | Gene Name | TF Type | Annotation |
|---|---|---|---|---|---|
| M.alba_G0008931 | L | AT3G07340 | CIB3 | bHLH | Photoperiodic flowering |
| M.alba_G0005811 | L | AT2G22850 | bZIP6 | bZIP | Vascular development |
| M.alba_G0006171 | L | AT1G75390 | bZIP44 | bZIP | Stress response and development |
| M.alba_G0017047 | L | AT1G27730 | STZ | C2H2 | Stress response |
| M.alba_G0013540 | L | AT4G29190 | AtC3H49 | C3H | Cold response |
| M.alba_G0018475 | L | AT3G47500 | CDF3 | Dof | Nitrogen responses |
| M.alba_G0006380 | L | AT4G39780 | ERF60 | ERF | Defense response and light stimulus |
| M.alba_G0020260 | L | AT2G44840 | ERF13 | ERF | Ethylene-activated signaling pathway |
| M.alba_G0020348 | L | AT4G17500 | ERF-1 | ERF | Ethylene-activated signaling pathway |
| M.alba_G0003406 | L | AT5G51190 | ERF105 | ERF | Response to cold stress |
| M.alba_G0016087 | L | AT2G47520 | ERF71 | ERF | Response to hypoxia stress |
| M.alba_G0016369 | L | AT1G50640 | ERF3 | ERF | Ethylene-activated signaling pathway |
| M.alba_G0001958 | L | AT3G02550 | LBD41 | LBD | Stress response |
| M.alba_G0017051 | L | AT3G49940 | LBD38 | LBD | Cellular metal ion homeostasis |
| M.alba_G0012646 | L | AT4G37260 | MYB73 | MYB | Stress responses and leaf senescence |
| M.alba_G0006600 | L | AT1G01720 | ANAC002 | NAC | Response to wounding and abscisic acid |
| M.alba_G0007251 | L | AT1G69490 | ANAC029 | NAC | Leaf senescence |
| M.alba_G0013712 | L | AT1G01720 | ANAC002 | NAC | Response to wounding and abscisic acid |
| M.alba_G0019459 | L | AT4G27410 | ANAC072 | NAC | ABA-mediated dehydration response |
| M.alba_G0007498 | L | AT1G13260 | EDF4 | RAV | Response to low temperature |
| M.alba_G0011100 | L | AT1G80840 | WRKY40 | WRKY | Fungus defense |
| M.alba_G0009278 | R | AT4G01500 | NGA4 | B3 | Leaf, stigma development |
| M.alba_G0005063 | R | AT4G17880 | MYC4 | bHLH | Activate JA-responses |
| M.alba_G0010656 | R | AT4G20970 | NA | bHLH | Dehydration stress memory |
| M.alba_G0018432 | R | AT3G47640 | PYE | bHLH | Regulating response to iron deficiency |
| M.alba_G0015523 | R | AT5G28770 | bZIP63 | Bzip | Circadian phase in response to sugars |
| M.alba_G0001737 | R | AT5G39660 | CDF2 | Dof | Photoperiodic flowering response |
| M.alba_G0003407 | R | AT4G17500 | ERF-1 | ERF | Ethylene-activated signaling pathway |
| M.alba_G0004215 | R | AT3G13040 | γ-YB2 | G2-like | Phosphate starvation |
| M.alba_G0016414 | R | AT2G31180 | MYB14 | MYB | Cold or wound stress |
| M.alba_G0006667 | R | AT5G64530 | ANAC104 | NAC | Xylem development |
| M.alba_G0013320 | R | AT1G80840 | WRKY40 | WRKY | Photosynthesis and Iron Homeostasis |
| M.alba_G0013615 | R | AT5G26170 | WRKY50 | WRKY | Defense response to fungus, JA response |
| M.alba_G0018966 | RLS | AT3G18960 | REM7 | B3 | Tissue development |
| M.alba_G0018963 | RLS | AT5G23090 | NF-YB13 | NF-YB | NA |
| M.alba_G0002019 | RS | AT5G16770 | MYB9 | MYB | Suberin biosynthesis and transport |
| M.alba_G0006489 | RS | AT5G64810 | WRKY51 | WRKY | Jasmonic acid-inducible defense responses |
| M.alba_G0013321 | RS | AT1G80840 | WRKY40 | WRKY | Jasmonic acid-inducible defense responses |
| M.alba_G0001053 | S | AT4G29930 | NA | bHLH | NA |
| M.alba_G0004848 | S | AT1G32640 | MYC2 | bHLH | Activate JA-responses |
| M.alba_G0012659 | S | AT1G72210 | BHLH96 | bHLH | Regulation of RNA polymerase II |
| M.alba_G0018293 | S | AT4G20970 | NA | bHLH | Defense response to fungus |
| M.alba_G0019112 | S | AT4G25440 | ZFWD1 | C3H | Development |
| M.alba_G0003582 | S | AT2G40140 | SZF2 | C3H | Response to biotic and abiotic stresses |
| M.alba_G0005725 | S | AT4G38960 | BBX19 | DBB | Photomorphogenesis and flowering |
| M.alba_G0005394 | S | AT5G51990 | CBF4 | ERF | Drought stress and abscisic acid treatment |
| M.alba_G0012399 | S | AT1G21910 | DREB26 | ERF | Response to JA and SA, abiotic stress |
| M.alba_G0016407 | S | AT2G31230 | ERF15 | ERF | Stress response |
| M.alba_G0005395 | S | AT5G51990 | CBF4 | ERF | Drought stress and abscisic acid treatment |
| M.alba_G0014394 | S | AT1G50420 | SCL-3 | GRAS | Response to gibberellin |
| M.alba_G0013518 | S | AT2G22840 | GRF1 | GRF | Leaf development. |
| M.alba_G0019709 | S | AT3G61890 | HB-12 | HD-ZIP | Leaf and stem development |
| M.alba_G0013465 | S | AT4G37540 | LBD39 | LBD | Cell wall biogenesis |
| M.alba_G0011537 | S | AT5G35550 | MYB123 | MYB | Anthocyanin biosynthesis |
| M.alba_G0012042 | S | AT1G17950 | MYB52 | MYB | Lignin, xylan and cellulose biosynthesis |
| M.alba_G0018463 | S | AT5G61420 | MYB28 | MYB | Seed development and aliphatic glucosinolate biosynthesis |
| M.alba_G0018447 | S | AT3G47600 | ATMYB94 | MYB | Cuticular wax biosynthesis |
| M.alba_G0011536 | S | AT2G16720 | MYB7 | MYB | General phenylpropanoid and lignin R2R3-MYB repressors |
| M.alba_G0018280 | S | AT5G14750 | MYB66 | MYB | Anthocyanin production and differentiation of trichome cells |
| M.alba_G0013188 | S | AT1G75250 | ATRL6 | MYB | Signal transduction |
| M.alba_G0019458 | S | AT3G15510 | ANAC056 | NAC | System development |
| M.alba_G0009218 | S | AT3G04070 | ANAC047 | NAC | Response to flooding |
| M.alba_G0009713 | S | AT5G63790 | ANAC102 | NAC | Mediating response to low oxygen stress |
| M.alba_G0011705 | S | AT4G14540 | NF-YB3 | NF-YB | Response to heat, response to water deprivation |
| M.alba_G0006285 | S | AT4G24660 | ATHB22 | ZF-HD | Embryo development ending in seed dormancy |
| M.alba_G0007224 | S | AT1G69600 | ATHB29 | ZF-HD | Early responsive to dehydration stress. |
| M.alba_G0013466 | SL | AT1G27730 | STZ | C2H2 | Stress response |
| M.alba_G0015192 | SL | AT3G46080 | NA | C2H2 | Transient stress |
| M.alba_G0003253 | SL | AT5G52020 | DREB | ERF | Glucosinolate metabolic process |
| M.alba_G0003254 | SL | AT5G51990 | CBF4 | ERF | Drought stress and abscisic acid treatment |
| M.alba_G0003536 | SL | AT2G40340 | AtERF48 | ERF | Response to abscisic and acid stress |
| M.alba_G0005396 | SL | AT5G51990 | CBF4 | ERF | Response to drought stress and abscisic acid treatment |
| M.alba_G0017242 | SL | AT4G34410 | ERF109 | ERF | Retarding programmed cell death under salt stress |
| M.alba_G0019814 | SL | AT1G19210 | ERF17 | ERF | JA, defense to biotic stresses |
| M.alba_G0000389 | SL | AT5G48150 | PAT1 | GRAS | Callus formation, photomorphogenesis, red, far-red light phototransduction |
| M.alba_G0011734 | SL | AT4G17230 | SCL13 | GRAS | Cellular response to hypoxia, heat |
| M.alba_G0004071 | SL | AT5G04760 | DIV2 | MYB | Negative roles in salt stress and is required for ABA signaling in Arabidopsis |
| M.alba_G0014170 | SL | AT3G44350 | ANAC061 | NAC | Response to salt stress |
| M.alba_G0005182 | SL | AT4G11070 | WRKY41 | WRKY | ABA defense response |
| M.alba_G0014899 | SL | AT2G38470 | WRKY33 | WRKY | Stress response |
| M.alba_G0019631 | SL | AT2G46400 | WRKY46 | WRKY | ABA signaling and auxin homeostasis in response to abiotic stress |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Kang, X.; Yu, T.; Yidilisi, K.; Zhang, L.; Cao, X.; Chao, N.; Liu, L. Comparative RNA-Seq Analysis Reveals the Organ-Specific Transcriptomic Response to Zinc Stress in Mulberry. Forests 2023, 14, 842. https://doi.org/10.3390/f14040842
Huang S, Kang X, Yu T, Yidilisi K, Zhang L, Cao X, Chao N, Liu L. Comparative RNA-Seq Analysis Reveals the Organ-Specific Transcriptomic Response to Zinc Stress in Mulberry. Forests. 2023; 14(4):842. https://doi.org/10.3390/f14040842
Chicago/Turabian StyleHuang, Shuai, Xiaoru Kang, Ting Yu, Keermula Yidilisi, Lin Zhang, Xu Cao, Nan Chao, and Li Liu. 2023. "Comparative RNA-Seq Analysis Reveals the Organ-Specific Transcriptomic Response to Zinc Stress in Mulberry" Forests 14, no. 4: 842. https://doi.org/10.3390/f14040842